Газообмен - Gas exchange
Газообмен это физический процесс пассивного движения газов распространение по поверхности. Например, эта поверхность может быть границей раздела воздух / вода в водоеме, поверхностью газового пузыря в жидкости, газопроницаемой мембрана, или биологическая мембрана который образует границу между организмом и его внеклеточной средой.
Газы постоянно потребляются и производятся клеточные и метаболические реакции в большинстве живых существ, поэтому требуется эффективная система газообмена, в конечном счете, между внутренним пространством клетки (-ей) и внешней средой. Небольшие, особенно одноклеточные организмы, такие как бактерии и простейшие, иметь высокий отношение площади поверхности к объему. У этих существ газообменная мембрана обычно представляет собой клеточная мембрана. Некоторые мелкие многоклеточные организмы, такие как плоские черви, также способны производить достаточный газообмен через кожа или кутикула что окружает их тела. Однако у большинства более крупных организмов, которые имеют небольшое отношение площади поверхности к объему, существуют специализированные структуры с извилистыми поверхностями, такие как жабры, легочные альвеолы и губчатый мезофилл обеспечить большую площадь, необходимую для эффективного газообмена. Эти извитые поверхности могут иногда проникать внутрь организма. Так обстоит дело с альвеолами, которые образуют внутреннюю поверхность млекопитающее легкое, губчатый мезофилл, который находится внутри листьев некоторые виды растений, или жабры тех моллюски что они есть, которые находятся в мантия полость.
В аэробные организмы, газообмен особенно важен для дыхание, что предполагает усвоение кислород (О
2) и выпуск углекислый газ (CO
2). И наоборот, в кислородные фотосинтезирующие организмы такие как большинство наземные растения поглощение углекислого газа и выделение как кислорода, так и водяного пара - основные процессы газообмена, происходящие в течение дня. Другие процессы газообмена важны для менее известных организмов: например углекислый газ, метан и водород обмениваются через клеточную мембрану метаногенные археи. В азотфиксация от диазотрофный бактерии и денитрификация от гетеротрофный бактерии (такие как Paracoccus denitrificans и различные псевдомонады ),[1] газообразный азот обменивается с окружающей средой, поглощается первым и выпускается в него вторым, в то время как гигантские трубчатые черви полагаться на бактерии для окисления сероводород извлечены из их глубоководной среды,[2] используя растворенный в воде кислород в качестве акцептора электронов.
Физические принципы газообмена
Распространение и площадь поверхности
Обмен газов происходит в результате распространение вниз по градиенту концентрации. Молекулы газа перемещаются из области, в которой они находятся в высокой концентрации, в область, в которой они находятся в низкой концентрации. Распространение - это пассивный процесс, что означает, что для питания транспорта не требуется энергии, и Закон Фика:[нужна цитата ]

В отношении типичной биологической системы, где два отсека («внутри» и «снаружи») разделены мембранным барьером и где газу позволяют спонтанно диффундировать вниз по градиенту его концентрации:[нужна цитата ]
- J это поток, количество газа диффузия на единицу площади мембраны в единицу времени. Обратите внимание, что это уже масштабировано для площади мембраны.
- D это коэффициент диффузии, которые будут различаться от газа к газу и от мембраны к мембране, в зависимости от размера молекулы рассматриваемого газа и природы самой мембраны (особенно ее вязкость, температура и гидрофобность ).
- φ это концентрация газа.
- Икс позиция по толщине мембраны.
- dφ/ дИкс следовательно, градиент концентрации через мембрану. Если два отсека по отдельности хорошо перемешаны, то это упрощается до разницы в концентрации газа между внутренним и внешним отсеками, деленной на толщину мембраны.
- Отрицательный знак означает, что диффузия всегда происходит в том направлении, которое со временем разрушит градиент концентрации, т.е. газ переходит от высокой концентрации к низкой до тех пор, пока в конечном итоге внутренние и внешние отсеки не достигнут равновесие.

Рисунок 1. Закон Фика для газообменной поверхности
Газы должны сначала раствориться в жидкости, чтобы диффундировать через мембрана, поэтому для всех биологических газообменных систем требуется влажная среда.[3] Как правило, чем выше градиент концентрации на газообменной поверхности, тем выше скорость диффузии через нее. И наоборот, чем тоньше газообменная поверхность (при той же разнице концентраций), тем быстрее газы будут диффундировать через нее.[4]
В приведенном выше уравнении J это поток выражается на единицу площади, поэтому увеличение площади не повлияет на ее значение. Однако увеличение доступной площади поверхности увеличит количество газа, который может диффундировать за определенное время.[4] Это связано с тем, что количество газа, диффундирующего в единицу времени (dq/ дт) является продуктом J и площадь газообменной поверхности, А:

Одноклеточные организмы такие как бактерии и амебы не имеют специализированных газообменных поверхностей, потому что они могут использовать большую площадь поверхности по сравнению с их объемом. Количество газа, которое организм производит (или требует) в данный момент времени, будет примерно пропорционально объему его газа. цитоплазма. Объем одноклеточного организма очень мал, поэтому он производит (и требует) относительно небольшое количество газа в заданное время. По сравнению с этим небольшим объемом, площадь его поверхности клеточная мембрана очень большой и без дополнительных модификаций подходит для нужд газообмена. Однако по мере увеличения размера организма его площадь и объем изменяются по-разному. Представьте себе воображаемый организм, представляющий собой куб с длиной стороны, L. Его объем увеличивается с кубом (L3) длины, но площадь внешней поверхности увеличивается только с квадратом (L2) его длины. Это означает, что внешняя поверхность быстро становится непригодной для быстро увеличивающихся потребностей в газообмене большего объема цитоплазмы. Кроме того, толщина поверхности, которую газы должны пересекать (dИкс по закону Фика) также может быть больше у более крупных организмов: в случае одноклеточного организма типичная клеточная мембрана имеет толщину всего 10 нм;[5] но у более крупных организмов, таких как круглые черви (Nematoda) эквивалентная поверхность обмена - кутикула - значительно толще на 0,5 мкм.[6]
Взаимодействие с кровеносными системами

В многоклеточный Таким образом, у организмов часто используются специализированные органы дыхания, такие как жабры или легкие, чтобы обеспечить дополнительную площадь поверхности для требуемой скорости газообмена с внешней средой. Однако расстояния между газообменником и более глубокими тканями часто слишком велики для диффузии, чтобы удовлетворить потребности этих тканей в газе. Поэтому газообменники часто присоединяются к газораспределительным устройствам. системы кровообращения, которые транспортируют газы равномерно ко всем тканям тела независимо от их расстояния от газообменника.[7]
Некоторые многоклеточные организмы, такие как плоские черви (Platyhelminthes) относительно большие, но очень тонкие, что позволяет их внешней поверхности тела действовать как поверхность газообмена без необходимости в специализированном газообменном органе. Поэтому у плоских червей отсутствуют жабры или легкие, а также отсутствует система кровообращения. Другие многоклеточные организмы, такие как губки (Porifera) по своей природе имеют большую площадь поверхности, потому что они очень пористые и / или разветвленные. Губкам не нужна система кровообращения или специальные органы газообмена, потому что их стратегия питания включает одностороннюю прокачку воды через их пористые тела с использованием бичеванный воротниковые клетки. Таким образом, каждая клетка тела губки подвергается постоянному воздействию свежей насыщенной кислородом воды. Следовательно, они могут полагаться на диффузию через клеточные мембраны для осуществления газообмена, необходимого для дыхания.[8]
У организмов, кровеносные системы которых связаны с их специализированными газообменными поверхностями, для взаимодействия между ними используется большое количество различных систем.
В противоток системы воздух (или, чаще, вода, содержащая растворенный воздух) всасывается в напротив направление потока крови в газообменнике. Противоточная система, такая как эта, поддерживает крутой градиент концентрации по длине газообменной поверхности (см. Нижнюю диаграмму на рис. 2). Такова ситуация в жабры рыбы и многие другие водные существа.[9] Вода из окружающей среды, содержащая газ, втягивается в одном направлении через поверхность газообмена, а кровоток в нижних жаберных капиллярах течет в противоположном направлении.[9][10][11] Хотя это теоретически позволяет почти полностью переносить респираторный газ от одной стороны теплообменника к другой, у рыб обычно менее 80% кислорода в воде, протекающей по жабрам, передается в кровь.[9]
Альтернативные варианты системы перекрестного тока встречается у птиц.[12][13] и тупиковые системы мешков, заполненных воздухом, найденные в легкие млекопитающих.[14][15] В прямоточный поток В системе кровь и газ (или жидкость, содержащая газ) движутся в одном направлении через газообменник. Это означает, что величина градиента меняется по длине поверхности газообмена, и обмен в конечном итоге прекратится, когда будет достигнуто равновесие (см. Верхнюю диаграмму на рис. 2).[9]Системы газообмена с прямоточным потоком, как известно, не используются в природе.
Млекопитающие

Газообменник у млекопитающих интернализуется, образуя легкие, как и у большинства крупных наземных животных.[нужна цитата ] Газообмен происходит в микроскопических тупиковых мешочках с воздухом, называемых альвеолы, где очень тонкая мембрана (называемая гемато-воздушный барьер ) отделяет кровь в альвеолярных капиллярах (в стенках альвеол) от альвеолярного воздуха в мешочках.
Обменная мембрана

Мембрана, через которую происходит газообмен в альвеолах (т. Е. Гемато-воздушный барьер), чрезвычайно тонкая (у людей в среднем толщина 2,2 мкм).[14] Он состоит из альвеолярные эпителиальные клетки, их подвальные мембраны и эндотелиальные клетки легочных капилляров (рис. 4).[14][16] Большая площадь поверхности мембраны обусловлена складыванием мембраны примерно в 300 миллионов альвеол с диаметром приблизительно 75-300 мкм каждая. Это обеспечивает чрезвычайно большую площадь поверхности (примерно 145 м 2).2), через которые может происходить газообмен.[14]
Альвеолярный воздух
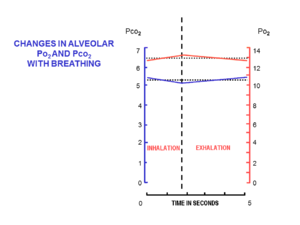

Воздуха доставляется в альвеолы в малых дозах (так называемый дыхательный объем ), от дыхание в (вдыхание ) и из (выдох ) сквозь дыхательные пути, набор относительно узких и умеренно длинных трубок, которые начинаются у носа или рта и заканчиваются альвеолами легких в груди. Воздух входит и выходит через один и тот же набор трубок, в которых поток идет в одном направлении во время вдоха и в противоположном направлении во время выдоха.
Во время каждого вдоха в состоянии покоя через нос поступает примерно 500 мл свежего воздуха. Его согревают и увлажняют, когда он течет через нос и глотка. К тому времени, когда воздух достигает трахеи, температура вдыхаемого воздуха составляет 37 ° C, и он насыщается водяным паром. По прибытии в альвеолы он разбавляется и тщательно смешивается с примерно 2,5–3,0 литрами воздуха, оставшимся в альвеолах после последнего выдоха. Этот относительно большой объем воздуха, который полупостоянно присутствует в альвеолах на протяжении дыхательного цикла, известен как функциональная остаточная емкость (FRC).[15]
В начале вдоха дыхательные пути заполняются неизменным альвеолярным воздухом, оставшимся после последнего выдоха. Это мертвый космос объем, который обычно составляет около 150 мл.[17] Это первый воздух, который повторно попадает в альвеолы при вдохе. Только после того, как воздух мертвого пространства вернулся в альвеолы, остаток дыхательного объема (500 мл - 150 мл = 350 мл) попадает в альвеолы.[15] Поступление такого небольшого объема свежего воздуха при каждом вдохе гарантирует, что состав FRC практически не изменится во время цикла дыхания (рис. 5).[15] Альвеолярный парциальное давление кислорода остается очень близким к 13–14кПа (100 мм рт. Ст.), А парциальное давление диоксида углерода минимально колеблется около 5,3 кПа (40 мм рт. ст.) на протяжении дыхательного цикла (вдоха и выдоха).[15] Соответствующие парциальные давления кислорода и углекислого газа в окружающем (сухом) воздухе на уровне моря составляют 21 кПа (160 мм рт. Ст.) И 0,04 кПа (0,3 мм рт. Ст.) Соответственно.[15]

Этот альвеолярный воздух, составляющий FRC, полностью окружает кровь в альвеолярных капиллярах (рис. 6). У млекопитающих газообмен происходит между альвеолярным воздухом (который значительно отличается от свежего воздуха) и кровью в альвеолярных капиллярах. Газы по обе стороны от газообменной мембраны уравновешиваются простой диффузией. Это гарантирует, что парциальные давления кислорода и углекислого газа в крови, покидающей альвеолярные капилляры и, в конечном счете, циркулирующей по всему телу, такие же, как и в FRC.[15]
Заметная разница между составом альвеолярного воздуха и окружающего воздуха может сохраняться, поскольку функциональная остаточная емкость содержится в тупиковых мешочках, связанных с наружным воздухом длинными узкими трубками (дыхательные пути: нос, глотка, гортань, трахея, бронхи и их ответвлений и подотраслей до бронхиолы ). Эта анатомия и тот факт, что легкие не опорожняются и не наполняются заново с каждым вдохом, обеспечивает млекопитающих «переносимой атмосферой», состав которой значительно отличается от современный атмосферный воздух.[18]
Состав воздуха в FRC тщательно контролируется путем измерения парциального давления кислорода и углекислого газа в артериальной крови. Если давление газа отклоняется от нормы, возникают рефлексы, изменяющие частоту и глубину дыхания таким образом, что нормальное состояние восстанавливается в течение секунд или минут.[15]
Легочное кровообращение
Вся кровь возвращается из тканей тела в правую часть сердце течет через альвеолярные капилляры прежде, чем снова окунуть его по телу. Проходя через легкие, кровь вступает в тесный контакт с альвеолярным воздухом, отделенным от него очень тонкой диффузионной мембраной, которая в среднем составляет всего около 2 мкм.[14] Таким образом, давление газа в крови быстро уравновесится с давлением в крови. альвеолы, гарантируя, что артериальная кровь, которая циркулирует по всем тканям тела, имеет напряжение кислорода 13-14 кПа (100 мм рт. ст.), а напряжение углекислого газа 5,3 кПа (40 мм рт. ст.). Эти артериальные парциальные давления кислорода и углекислого газа равны гомеостатически управляемый. Подъем артериального , и, в меньшей степени, падение артериального , рефлекторно вызовет более глубокое и быстрое дыхание, пока давление газов в крови не вернется к норме. Обратное происходит, когда давление углекислого газа падает или, опять же, в меньшей степени, повышается давление кислорода: частота и глубина дыхания снижаются до тех пор, пока не восстанавливается нормальность газов крови.


Поскольку кровь, поступающая в альвеолярные капилляры, имеет в среднем 6 кПа (45 мм рт. ст.), в то время как давление в альвеолярном воздухе составляет 13 кПа (100 мм рт. ст.), будет чистая диффузия кислорода в капиллярную кровь, изменяющую состав 3 литров альвеолярного воздуха. немного. Точно так же, поскольку кровь, поступающая в альвеолярные капилляры, имеет также около 6 кПа (45 мм рт. ст.), в то время как альвеолярный воздух составляет 5,3 кПа (40 мм рт. ст.), наблюдается чистое перемещение углекислого газа из капилляров в альвеолы. Изменения, вызываемые этими чистыми потоками отдельных газов в функциональную остаточную емкость и из нее, требуют замены примерно 15% альвеолярного воздуха окружающим воздухом каждые 5 секунд или около того. Это очень жестко контролируется постоянным мониторингом напряжений газов артериальной крови (которые точно отражают парциальные давления дыхательных газов в альвеолярном воздухе). аортальные тела, то каротидные тела, а датчик газов крови и pH на передней поверхности продолговатый мозг в мозгу. В легких также есть датчики кислорода и углекислого газа, но они в первую очередь определяют диаметр бронхиолы и легочные капилляры, и поэтому несут ответственность за направление потока воздуха и крови к разным частям легких.
Только в результате точного поддержания состава 3 литров альвеолярного воздуха с каждым вдохом некоторое количество углекислого газа выбрасывается в атмосферу, а некоторое количество кислорода забирается из внешнего воздуха. Если за короткий период времени было потеряно больше углекислого газа, чем обычно. гипервентиляция, дыхание замедлится или остановится до тех пор, пока альвеолярный вернулся к 5,3 кПа (40 мм рт. ст.). Поэтому, строго говоря, неверно, что основная функция дыхательной системы - избавлять организм от «отходов» углекислого газа. Фактически общая концентрация углекислого газа в артериальной крови составляет около 26 мМ (или 58 мл на 100 мл),[19] по сравнению с концентрацией кислорода в насыщенной артериальной крови около 9 мМ (или 20 мл на 100 мл крови).[15] Эта большая концентрация углекислого газа играет ключевую роль в определение и поддержание pH внеклеточной жидкости. Углекислый газ, выдыхаемый при каждом вдохе, правильнее было бы рассматривать как побочный продукт внеклеточной жидкости организма. углекислый газ и pH-гомеостаты
Если эти гомеостаты скомпрометированы, тогда респираторный ацидоз, или респираторный алкалоз произойдет. В долгосрочной перспективе они могут быть компенсированы почечной корректировкой H+ и HCO3− концентрации в плазме; но поскольку это требует времени, синдром гипервентиляции может, например, произойти, когда возбуждение или тревога заставляют человека дышать быстро и глубоко[20] таким образом выдувая слишком много CO2 из крови в наружный воздух, вызывая набор неприятных симптомов, которые возникают из-за чрезмерно высокого pH внеклеточной жидкости.[21]
Кислород имеет очень низкую растворимость в воде и поэтому переносится кровью в слабом сочетании с гемоглобин. Кислород удерживается в гемоглобине четырьмя двухвалентное железо -содержащий гем групп на молекулу гемоглобина. Когда все группы гема содержат один O2 Говорят, что каждая молекула крови «насыщена» кислородом, и никакое дальнейшее повышение парциального давления кислорода не приведет к значительному увеличению концентрации кислорода в крови. Большая часть углекислого газа в крови переносится в виде HCO.3− ионы в плазме. Однако конверсия растворенного CO2 в HCO3− (за счет добавления воды) слишком медленный для скорости, с которой кровь циркулирует через ткани, с одной стороны, и альвеолярные капилляры, с другой. Следовательно, реакция катализируется карбоангидраза, фермент внутри красные кровяные клетки.[22] Реакция может идти в любом направлении в зависимости от преобладающего парциального давления диоксида углерода. Небольшое количество диоксида углерода переносится белковой частью молекул гемоглобина в виде карбамино группы. Общая концентрация углекислого газа (в виде ионов бикарбоната, растворенного CO2, и карбаминогруппы) в артериальной крови (т.е. после того, как она уравновесится с альвеолярным воздухом) составляет около 26 мМ (или 58 мл / 100 мл),[19] по сравнению с концентрацией кислорода в насыщенной артериальной крови около 9 мМ (или 20 мл / 100 мл крови).[15]
Другие позвоночные
Рыбы

Содержание растворенного кислорода в пресная вода составляет примерно 8–10 миллилитров на литр по сравнению с воздухом, который составляет 210 миллилитров на литр.[23] Вода в 800 раз плотнее воздуха[24] и в 100 раз более вязкой.[23] Следовательно, скорость диффузии кислорода в воздухе в 10 000 раз выше, чем в воде.[23] Следовательно, использование мешковидных легких для удаления кислорода из воды было бы недостаточно эффективным для поддержания жизни.[23] Вместо использования легких, газообмен происходит на поверхности сильно васкуляризованных жабры. Жабры - это специализированные органы, содержащие нити, которые далее делятся на ламели. Ламели содержат капилляры которые обеспечивают большую площадь поверхности и короткие расстояния диффузии, поскольку их стенки чрезвычайно тонкие.[25] Жаберные тычинки находятся в системе обмена, чтобы фильтровать пищу и поддерживать чистоту жабр.
Жабры используют противоток система, которая увеличивает эффективность поглощения кислорода (и потери отработанного газа).[9][10][11] Кислородная вода втягивается через рот и проходит через жабры в одном направлении, а кровь течет через ламели в противоположном направлении. Эта противоток поддерживает резкие градиенты концентрации по всей длине каждого капилляра (см. диаграмму на «Взаимодействие с кровеносными системами» раздел выше). Кислород может непрерывно диффундировать по градиенту в кровь, а углекислый газ - вниз по градиенту в воду.[10] Деоксигенированная вода в конечном итоге будет проходить через крышка (жаберная крышка). Хотя противоточные системы обмена теоретически позволяют почти полностью переносить респираторный газ с одной стороны теплообменника на другую, у рыб обычно менее 80% кислорода в воде, протекающей по жабрам, передается в кровь.[9]
Амфибии
У земноводных есть три основных органа, участвующих в газообмене: легкие, кожа и жабры, которые можно использовать по отдельности или в различных комбинациях. Относительная важность этих структур зависит от возраста, среды обитания и вида амфибии. Кожа амфибий и их личинок сильно васкуляризована, что приводит к относительно эффективному газообмену, когда кожа влажная. Личинки амфибий, например, до метаморфоза головастик этап лягушки, также есть внешние жабры. Жабры всасываются в тело во время метаморфоза, после чего легкие возьмут верх. Легкие обычно проще, чем в легких. другие наземные позвоночные с небольшим количеством внутренних перегородок и более крупными альвеолами; однако жабы, которые проводят больше времени на суше, имеют большую альвеолярную поверхность и более развитые легкие. Чтобы увеличить скорость газообмена за счет диффузии, земноводные поддерживают градиент концентрации на респираторной поверхности, используя процесс, называемый буккальное откачивание.[26] Нижний дно рта перемещается «качающим» образом, что можно наблюдать невооруженным глазом.
Рептилии
Все рептилии дышите легкими. В чешуйчатые (в ящерицы и змеи ) вентиляция управляется осевая мускулатура, но эта мускулатура также используется во время движения, поэтому некоторые чешуйки полагаются на буккальное откачивание для поддержания эффективности газообмена.[27]
За счет жесткости черепаха и черепаха снаряды, значительное расширение и сжатие грудной клетки затруднено. Черепахи и черепахи зависят от мышечных слоев, прикрепленных к их панцирям, которые обвивают их легкие, наполняя и опустошая их.[28] Некоторые водные черепахи также могут качать воду в сильно васкуляризованный рот или клоака добиться газообмена.[29][30]
Крокодилы имеют структуру, похожую на диафрагму млекопитающих - диафрагмальную мышцу, - но эта мышца помогает создавать однонаправленный поток воздуха через легкие, а не приливный поток: это больше похоже на воздушный поток, наблюдаемый в птицы чем у млекопитающих.[31] Во время вдоха диафрагмальная мышца втягивает печень назад, раздувая легкие в создаваемое пространство.[32][33] Воздух поступает в легкие из бронхов во время вдоха, но во время выдоха воздух выходит из легких в бронх другим путем: это одностороннее движение газа достигается за счет аэродинамических клапанов в дыхательных путях.[34][35]
Птицы


У птиц есть легкие, но без диафрагмы. Они полагаются в основном на воздушные мешочки для вентиляция. Эти воздушные мешочки не играют непосредственной роли в газообмене, но помогают перемещать воздух в одном направлении через поверхности газообмена в легких. При вдохе свежий воздух забирается из трахеи в задние воздушные мешочки и парабронхи которые ведут из задних воздушных мешков в легкие. Воздух, поступающий в легкие, присоединяется к воздуху, который уже находится в легких, и направляется вперед через газообменник в передние воздушные мешочки. Во время выдоха задние воздушные мешки вытесняют воздух в ту же область. парабронхи легких, течет в том же направлении, что и при вдохе, обеспечивая непрерывный газообмен независимо от дыхательного цикла. Воздух, выходящий из легких во время выдоха, присоединяется к воздуху, выходящему из передних воздушных мешков (оба они состоят из «отработанного воздуха», прошедшего через газообменник), поступающего в трахею для выдоха (рис. 10).[13] Селективный бронхоспазм в различных точках ветвления бронхов гарантирует, что воздух не будет течь через бронхи во время вдоха и выдоха, как это происходит у млекопитающих, а следует путями, описанными выше.
Однонаправленный поток воздуха через парабронхи обеспечивает обмен дыхательных газов с воздухом. перекрестное течение кровоток (рис. 9).[12][13] Парциальное давление O2 () в парабронхиолах убывает по длине как O2 проникает в кровь. Капилляры, выходящие из теплообменника около входа воздушного потока, забирают больше O2 чем капилляры, уходящие около выходного конца парабронхов. Когда содержимое всех капилляров смешивается, последний смешанной легочной венозной крови выше, чем у выдыхаемого воздуха, но ниже, чем у вдыхаемого воздуха.[12][13]
Растения
В газообмене растений доминируют углекислый газ, кислород и водяной пар. CO
2 единственный источник углерода для автотрофный рост на фотосинтез, и когда растение активно фотосинтезирует на свету, оно будет поглощать углекислый газ, и потеря водяного пара и кислород. Ночью растения дышать, и газообмен частично меняется на противоположный: водяной пар все еще теряется (но в меньшей степени), но теперь поглощается кислород и выделяется углекислый газ.[36]

Газообмен у растений происходит в основном через листья. Газы диффундируют в межклеточные пространства листа и из них через поры, называемые устьица, которые обычно находятся на нижней поверхности листа. Газы попадают в фотосинтетическую ткань листа через растворение на влажной поверхности палисада и губчатой ткани. мезофилл клетки. Губчатые клетки мезофилла неплотно упакованы, что позволяет увеличить площадь поверхности и, как следствие, увеличить скорость газообмена. Поглощение углекислого газа обязательно приводит к потере водяного пара,[37] потому что обе молекулы входят и выходят через одни и те же устьица, поэтому растения сталкиваются с дилеммой газообмена: CO
2 не теряя слишком много воды. Следовательно, потеря воды с других частей листа сводится к минимуму за счет восковой кутикулы на листе. эпидермис. Размер стомы регулируется путем открытия и закрытия двух замыкающие клетки: отечность этих клеток определяет состояние устьичного отверстия, что само регулируется водным стрессом. Растения показаны метаболизм крассулоидной кислоты устойчивы к засухе ксерофиты и осуществляют почти весь газообмен ночью, потому что только ночью эти растения открывают устьица. Открытие устьиц только на ночь сводит к минимуму потери водяного пара, связанные с поглощением углекислого газа. Однако это происходит за счет медленного роста: растение должно накапливать углекислый газ в виде яблочная кислота для использования в течение дня и не может хранить неограниченное количество.[38]

Измерения газообмена являются важными инструментами в науке о растениях: обычно это включает герметизацию растения (или его части) в камере и измерение изменений концентрации диоксида углерода с помощью инфракрасный газоанализатор. Если условия окружающей среды (влажность, CO
2 концентрация, свет и температура ) полностью контролируются, измерения CO
2 поглощение и выпуск воды раскрывают важную информацию о CO
2 ассимиляция и испарение ставки. Межклеточный CO
2 концентрация раскрывает важную информацию о фотосинтетическом состоянии растений.[39][40] В определенных обстоятельствах можно использовать более простые методы: индикатор гидрокарбоната может использоваться для мониторинга потребления CO
2 в растворе, содержащем один лист растения при разной интенсивности света,[41] и производство кислорода водорослями Элодея можно измерить, просто собрав газ в погруженную пробирку, содержащую небольшой кусочек растения.
Беспозвоночные
Механизм газообмена у беспозвоночных зависит от их размера, стратегии питания и среды обитания (водной или наземной).

В губки (Porifera) сидячие существа, то есть они не могут двигаться самостоятельно и обычно остаются привязанными к своему субстрат. Они получают питательные вещества через поток воды через свои клетки и обмениваются газами путем простой диффузии через клеточные мембраны. Поры называются Остия втяните воду в губку, и впоследствии вода будет циркулировать через губку посредством клеток, называемых хоаноциты который имеет волосовидные структуры которые перемещают воду через губку.[42]

В книдарийцы включают кораллы, морские анемоны, медуза и гидры. Эти животные всегда обитают в водной среде, от пресной до соленой. У них нет выделенных органы дыхания; Вместо этого каждая клетка их тела может поглощать кислород из окружающей воды и выделять в нее отработанные газы. Одним из ключевых недостатков этой функции является то, что книдарие могут умереть в среде, где вода застойный, поскольку они истощают воду его кислород поставка.[43] Кораллы часто вступают в симбиоз с другими организмами, особенно с фотосинтезирующими. динофлагелляты. В этом симбиоз, то коралл обеспечивает укрытие, а другой организм обеспечивает коралл питательными веществами, включая кислород.

В круглые черви (Нематода), плоские черви (Platyhelminthes) и многие другие мелкие беспозвоночные животные, живущие в водных или других влажных средах обитания, не имеют специальной поверхности газообмена или кровеносной системы. Вместо этого они полагаются на распространение из CO
2 и О
2 прямо через кутикулу.[44][45] Кутикула - это полупроницаемый внешний слой их тел.
Другие водные беспозвоночные, такие как большинство моллюски (Mollusca) и крупнее ракообразные (Ракообразные), такие как лобстеры, имеют жабры, аналогичные жабрам рыб, которые действуют аналогичным образом.

В отличие от упомянутых выше групп беспозвоночных, насекомые обычно являются земными и обмениваются газами через влажную поверхность в непосредственном контакте с атмосферой, а не в контакте с окружающей водой. Насекомое экзоскелет непроницаем для газов, в том числе для водяного пара, поэтому они имеют более специализированную систему газообмена, требующую, чтобы газы передавались непосредственно к тканям через сложную сеть трубок. Эта дыхательная система отделена от их кровеносной системы. Газы входят в тело и выходят из него через отверстия, называемые дыхальца, расположенный сбоку по грудная клетка и брюшная полость. Подобно растениям, насекомые могут контролировать открытие и закрытие дыхалец, но вместо того, чтобы полагаться на тургорное давление, они полагаются на мышечные сокращения.[46] Эти схватки привести к тому, что брюшко насекомого будет закачиваться и выдыхаться. Дыхальца соединены с трубками, называемыми трахеи, которые многократно разветвляются и разветвляются в теле насекомого. Эти отделения заканчиваются специализированными трахеольные клетки который обеспечивает тонкую влажную поверхность для эффективного газообмена непосредственно с ячейками.[47]
Другая основная группа наземных членистоногие, то паукообразные (пауки, скорпион, клещи, и их родственники) обычно производят газообмен с книга легкое.[48]
Краткое изложение основных систем газообмена
| Площадь поверхности | Расстояние диффузии | Поддержание градиента концентрации | Органы дыхания | |
|---|---|---|---|---|
| Человек | Всего альвеол[49] = 70–100 м2 | Альвеола и капилляр (две клетки) | Постоянный кровоток в капиллярах; дыхание | Легкие |
| Рыбы | Много ламелей и нитей на жабру | Обычно одна ячейка | Противоточный поток | Жабры |
| Насекомые | Специализированная трахеольная клетка | Одна ячейка | Буккальное сцеживание | Дыхальца |
| Губки | Остия поры | Одна ячейка | Движение воды | Никто |
| Плоские черви | Плоская форма тела | Обычно одна ячейка | Противоточный поток | Никто |
| Книдарианцы | Оральные руки | Обычно одна ячейка | Движение воды | Никто |
| Рептилии | Много ламелей и нитей на жабру[требуется разъяснение ] | Альвеола и капилляр (две клетки) | Противоточный поток | Легкие |
| Амфибии | Много ламелей и нитей на жабру | Альвеола и капилляр (две клетки) или одна клетка | Противоточный поток | Легкие, кожа и жабры |
| Растения | Высокая плотность устьиц; воздушные пространства внутри листа | Одна ячейка | Постоянный воздушный поток | Устьица |
Смотрите также
- Дыхательная система - Биологическая система у животных и растений для газообмена
использованная литература
- ^ Carlson, C.A .; Ингрэм, Дж. Л. (1983). "Сравнение денитрификации Pseudomonas stutzeri, Синегнойная палочка, и Paracoccus denitrificans". Appl. Environ. Микробиол. 45 (4): 1247–1253. Дои:10.1128 / AEM.45.4.1247-1253.1983. PMID 6407395.
- ^ К. Майкл Хоган. 2011 г. Сера. Энциклопедия Земли, ред. А. Йоргенсен и К. Дж. Кливленд, Национальный совет по науке и окружающей среде, Вашингтон, округ Колумбия В архиве 28 октября 2012 г. Wayback Machine
- ^ Пийпер Дж., Дежурс П., Хааб П. и Ран Х. (1971). «Понятия и основные величины в физиологии газообмена». Физиология дыхания. 13 (3): 292–304. Дои:10.1016 / 0034-5687 (71) 90034-х. PMID 5158848.CS1 maint: использует параметр авторов (ссылка на сайт)
- ^ а б Кети СС (1951). «Теория и приложения обмена инертного газа в легких и тканях». Фармакологические обзоры. 3 (1): 1–41. PMID 14833874.
- ^ Schneiter, R; Брюггер, В; Sandhoff, R; Зелльниг, G; Лебер, А; Лампл, М; Athenstaedt, K; Храстник, Ц; Эдер, S; Daum, G; Paltauf, F; Виланд, FT; Кольвейн, SD (1999). «Анализ тандемной масс-спектрометрии с ионизацией электрораспылением (ESI-MS / MS) молекулярного липидного состава субклеточных мембран дрожжей выявляет сортировку / ремоделирование на основе ацильных цепей отдельных молекулярных видов на пути к плазматической мембране». Журнал клеточной биологии. 146 (4): 741–54. Дои:10.1083 / jcb.146.4.741. ЧВК 2156145. PMID 10459010.
- ^ Кокс, Г. Н. (1 июля 1981 г.). "Кутикула Caenorhabditis elegans: его изолированность и частичная характеристика ». Журнал клеточной биологии. 90 (1): 7–17. Дои:10.1083 / jcb.90.1.7. ЧВК 2111847. PMID 7251677.
- ^ Флегр, Ярослав. «Замороженная эволюция». Замороженная эволюция. Получено 21 марта 2017.
- ^ «Дыхательная система и прямая диффузия». Безграничный. Получено 19 марта 2017.
- ^ а б c d е ж Кэмпбелл, Нил А. (1990). Биология (Второе изд.). Редвуд-Сити, Калифорния: Бенджамин / Каммингс Паблишинг Компани, Инк., Стр. 836–838. ISBN 978-0-8053-1800-5.
- ^ а б c Хьюз GM (1972). «Морфометрия жабр рыб». Физиология дыхания. 14 (1–2): 1–25. Дои:10.1016 / 0034-5687 (72) 90014-х. PMID 5042155.
- ^ а б Сторер, Трейси I .; Usinger, R.L .; Стеббинс, Роберт С .; Нюбаккен, Джеймс У. (1997). Общая зоология (шестое изд.). Нью-Йорк: Макгроу-Хилл. стр.668–670. ISBN 978-0-07-061780-3.
- ^ а б c d Скотт, Грэм Р. (2011). «Комментарий: Повышенная производительность: уникальная физиология птиц, летающих на больших высотах». Журнал экспериментальной биологии. 214 (15): 2455–2462. Дои:10.1242 / jeb.052548. PMID 21753038.
- ^ а б c d Ритчсон, Г. «BIO 554/754 - Орнитология: дыхание птиц». Департамент биологических наук Университета Восточного Кентукки. Получено 2009-04-23.
- ^ а б c d е Уильямс, Питер Л; Уорик, Роджер; Дайсон, Мэри; Баннистер, Лоуренс Х. (1989). Анатомия Грея (Тридцать седьмое изд.). Эдинбург: Черчилль Ливингстон. С. 1278–1282. ISBN 0443-041776.
- ^ а б c d е ж г час я j Тортора, Джерард Дж .; Анагностакос, Николас П. (1987). Основы анатомии и физиологии (Пятое изд.). Нью-Йорк: Harper & Row, Publishers. стр.570–580. ISBN 978-0-06-350729-6.
- ^ «Газообмен у человека». Получено 19 марта 2013.
- ^ «Объем мертвого пространства - Оксфордский справочник».
- ^ Лавлок, Джеймс (1991). Healing Gaia: практическая медицина для планеты. Нью-Йорк: Книги Гармонии. стр.21 –34, 73–88. ISBN 978-0-517-57848-3.
- ^ а б Diem, K .; Лентнер, К. (1970). «Кровь - неорганические вещества». в: Научные таблицы (Седьмое изд.). Базель, Швейцария: CIBA-GEIGY Ltd. стр. 571.
- ^ Шу, Британская Колумбия; Чанг, ГГ; Ли, ФГ; Tzeng, DS; Lin, HY; Легкое, FW (2007-10-31). «Родительская привязанность, преморбидная личность и психическое здоровье у молодых мужчин с синдромом гипервентиляции». Психиатрические исследования. 153 (2): 163–70. Дои:10.1016 / j.psychres.2006.05.006. PMID 17659783. S2CID 3931401.
- ^ "eMedicine - синдром гипервентиляции: статья Эдварда Ньютона, доктора медицины". Получено 2007-12-20.
- ^ Раймонд Х и Свенсон Э (2000). «Распределение и физиологическое значение карбоангидразы в органах газообмена позвоночных». Физиология дыхания. 121 (1): 1–12. Дои:10.1016 / s0034-5687 (00) 00110-9. PMID 10854618.CS1 maint: использует параметр авторов (ссылка на сайт)
- ^ а б c d М. б. против Робертса; Майкл Рейсс; Грейс Монгер (2000). Продвинутая биология. Лондон, Великобритания: Нельсон. С. 164–165.
- ^ Тайсон, П. Д .; Престон-Уайт, Р.А. (2013). Погода и климат юга Африки (Второе изд.). Кейптаун, Южная Африка: Издательство Оксфордского университета. п. 14. ISBN 9780195718065.
- ^ Ньюстед Джеймс Д. (1967). «Тонкое строение дыхательных пластинок жабр костистых животных». Исследования клеток и тканей. 79 (3): 396–428. Дои:10.1007 / bf00335484. PMID 5598734. S2CID 20771899.
- ^ Брейнерд, Э. Л. (1999). «Новые взгляды на эволюцию механизмов вентиляции легких у беспозвоночных». Экспериментальная биология онлайн. 4 (2): 1–28. Дои:10.1007 / s00898-999-0002-1. S2CID 35368264.
- ^ Тейлор, Э. У .; Кэмпбелл, Х. А .; Лейте, С; Abe, A. S .; Ван, Т. (2007). «Дыхание у рептилий». Сравнительная биохимия и физиология A - Молекулярная и интегративная физиология. 148: S110 – S111. Дои:10.1016 / j.cbpa.2007.06.431.
- ^ Кляйн, Уилфид; Абэ, Аугусто; Андраде, Денис; Перри, Стивен (2003). «Структура постпеченочной перегородки и ее влияние на висцеральную топологию ящерицы тегу, Tupinambis merianae (Teidae: Reptilia) ". Журнал морфологии. 258 (2): 151–157. Дои:10.1002 / jmor.10136. PMID 14518009. S2CID 9901649.
- ^ Оренштейн, Рональд (2001). Черепахи, черепахи и черепахи: выжившие в доспехах. Книги Светлячка. ISBN 978-1-55209-605-5.
- ^ Feder, Martin E .; Бурггрен, Уоррен В. (1985). «Кожный газообмен у позвоночных: дизайн, модели, контроль и последствия» (PDF). Биологические обзоры. 60 (1): 1–45. Дои:10.1111 / j.1469-185X.1985.tb00416.x. PMID 3919777. S2CID 40158158.
- ^ Фермер, CG; Сандерс, К. (2010). «Однонаправленный поток воздуха в легких аллигаторов». Наука. 327 (5963): 338–340. Bibcode:2010Sci ... 327..338F. Дои:10.1126 / наука.1180219. PMID 20075253. S2CID 206522844.
- ^ Farmer, C.G .; Кэрриер Д. Р. (2000). «Тазовое стремление у американского аллигатора (Аллигатор миссисипиенсис)". Журнал экспериментальной биологии. 203 (11): 1679–1687. PMID 10804158.
- ^ Munns, S.L .; Owerkowicz, T .; Andrewartha, S.J .; Фраппелл, П. Б. (2012). «Дополнительная роль диафрагмальной мышцы в вентиляции легких у устьевого крокодила. Crocodylus porosus". Журнал экспериментальной биологии. 215 (5): 845–852. Дои:10.1242 / jeb.061952. PMID 22323207.
- ^ Farmer, C.G .; Сандерс, К. (2010). «Однонаправленный поток воздуха в легких аллигаторов» (PDF). Наука. 327 (5963): 338–340. Bibcode:2010Sci ... 327..338F. Дои:10.1126 / наука.1180219. PMID 20075253. S2CID 206522844. Архивировано из оригинал (PDF) на 2016-06-24. Получено 2017-04-20.
- ^ Schachner, E. R .; Hutchinson, J. R .; Фармер, К. (2013). «Анатомия легких у нильского крокодила и эволюция однонаправленного воздушного потока у архозавров». PeerJ. 1: e60. Дои:10.7717 / peerj.60. ЧВК 3628916. PMID 23638399.
- ^ Уитмарш Дж, Говинджи (1999). «Глава 2: Основной фотосинтетический процесс». В Singhal GS, Renger G, Sopory SK, Irrgang KD, Govindjee (ред.). Концепции фотобиологии: фотосинтез и фотоморфогенез. Бостон: Kluwer Academic Publishers. п. 13. ISBN 978-0-7923-5519-9.
- ^ К. Рашке (1976). «Как Стома решает дилемму противоположных приоритетов». Фил. Пер. R. Soc. Лондон. B. 273 (927): 551–560. Bibcode:1976РСПТБ.273..551Р. Дои:10.1098 / рстб.1976.0031.
- ^ Тинг, ИП (1985). «Кислотный метаболизм у крассулейных». Ежегодный обзор физиологии растений. 36 (1): 595–622. Дои:10.1146 / annurev.pp.36.060185.003115. HDL:10150/552219.
- ^ S Von Caemmerer; Г. Д. Фаркуар (1981). «Некоторые взаимосвязи между биохимией фотосинтеза и газообменом листьев». Planta. 153 (4): 376–387. Дои:10.1007 / bf00384257. PMID 24276943. S2CID 22760377.
- ^ Переносная газообменная флуоресцентная система GFS-3000. Справочник по эксплуатации (PDF), 20 марта 2013 г.
- ^ BBC Bitesize - Биология GCSE - Газообмен в растениях
- ^ Андерсон, Д. (2001) Зоология беспозвоночных Oxford University Press
- ^ «Книдарианская респираторная система». study.com. Получено 20 марта 2017.
- ^ «Дыхательная система нематод». study.com. Получено 20 марта 2017.
- ^ "Дыхательная система Platyhelminthes". http://rspp.weebly.com. Получено 20 марта 2017. Внешняя ссылка в
| сайт =(Помогите) - ^ Lane, N.J .; Харрисон, Дж. Б. (1986). «Соединения и цитоскелет в тканях насекомых». Журнал клеточной биологии. 103 (5): A69.
- ^ Клоуден, М. Дж. 2007. Физиологические системы у насекомых. Elsevier / Academic Press. стр. 440-442
- ^ Гарвуд, Рассел Дж. И Эджкомб, Грегори Д. (сентябрь 2011 г.). «Ранние наземные животные, эволюция и неопределенность». Эволюция: образование и пропаганда. 4 (3): 489–501. Дои:10.1007 / s12052-011-0357-у.
- ^ Бассет Дж., Крон С., Саумон Дж. (1987). «Значение активного ионного транспорта в трансальвеолярном поглощении воды: исследование изолированного легкого крысы». Журнал физиологии. 384: 311–324. Дои:10.1113 / jphysiol.1987.sp016456. ЧВК 1192264. PMID 3656149.CS1 maint: использует параметр авторов (ссылка на сайт)