Гем - Heme

Гем (Американский английский) или гем (Британский английский[1]) является веществом, предшественником гемоглобин, что необходимо для привязки кислород в кровоток. Гем биосинтезированный в обоих Костный мозг и печень.[2]
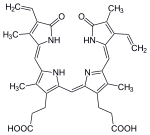
С микробиологической точки зрения гем - это координационный комплекс "состоящий из иона железа, координированного с порфирин действуя как тетрадентатный лиганд, и к одному или двум аксиальным лигандам ".[3] Определение расплывчатое, и на многих изображениях аксиальные лиганды отсутствуют.[4] Среди металлопорфиринов, используемых металлопротеины в качестве протезные группы, гем - один из наиболее широко используемых[5] и определяет семейство белков, известных как гемопротеины. Гемы чаще всего считаются компонентами гемоглобин, красный пигмент в кровь, но также встречаются в ряде других биологически важные гемопротеины, такие как миоглобин, цитохромы, каталазы, гем пероксидаза, и эндотелиальная синтаза оксида азота.[6][7]
Слово гем происходит от Греческий αἷμα Хайма что означает «кровь».
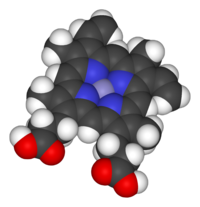
Функция

Гемопротеины имеют разнообразные биологические функции, включая транспортировку двухатомный газы, химические катализ, обнаружение двухатомного газа и перенос электронов. Гемовое железо служит источником или стоком электронов во время переноса электронов или редокс химия. В пероксидаза реакции, порфирин молекула также служит источником электронов, способным делокализовать радикальные электроны в сопряженном кольце. При транспортировке или обнаружении двухатомных газов газ связывается с гемовым железом. При обнаружении двухатомных газов связывание газа лиганд к гемовому железу конформационные изменения в окружающем белке.[8] В общем, двухатомные газы связываются только с восстановленным гемом, как двухвалентное Fe (II), в то время как большинство пероксидаз совершают цикл между Fe (III) и Fe (IV), а гемепротеины участвуют в окислительно-восстановительном, окислительно-восстановительном цикле между Fe (II) и Fe (III).
Было высказано предположение, что первоначальная эволюционная функция гемопротеины был перенос электрона в примитивном сера -основан фотосинтез пути в наследственных цианобактерии -подобно организмы до появления молекулярных кислород.[9]
Гемопротеины достигают своего замечательного функционального разнообразия за счет изменения среды макроцикла гема в белковой матрице.[10] Например, способность гемоглобин эффективно доставлять кислород к ткани связано с конкретными аминокислота остатки, расположенные рядом с молекулой гема.[11] Гемоглобин обратимо связывается с кислородом в легких, когда pH высокий, а углекислый газ концентрация низкая. Когда ситуация меняется на противоположную (низкий pH и высокая концентрация углекислого газа), гемоглобин выделяет кислород в ткани. Это явление, которое утверждает, что кислород гемоглобина сродство связывания является обратно пропорциональный как для кислотность и концентрация углекислого газа, известная как Эффект Бора.[12] Молекулярный механизм за этим эффектом стоит стерический организация глобин цепь; а гистидин остаток, расположенный рядом с гемовой группой, становится положительно заряженным в кислых условиях (которые вызваны растворенный CO2 в работающих мышцах и др.), выделяя кислород из гемовой группы.[13]
Типы
Основные рубцы
Существует несколько биологически важных видов гема:
| Гем А | Heme B | Heme C | Heme O | ||
|---|---|---|---|---|---|
| Номер PubChem | 7888115 | 444098 | 444125 | 6323367 | |
| Химическая формула | C49ЧАС56О6N4Fe | C34ЧАС32О4N4Fe | C34ЧАС36О4N4S2Fe | C49ЧАС58О5N4Fe | |
| Функциональная группа в C3 |  | –CH (OH) CH2Далеко | –CH = CH2 | –CH (цистеинS-yl ) CH3 | –CH (OH) CH2Далеко |
| Функциональная группа в C8 | –CH = CH2 | –CH = CH2 | –CH (цистеинS-yl ) CH3 | –CH = CH2 | |
| Функциональная группа в C18 | –CH = O | –CH3 | –CH3 | –CH3 | |


Самый распространенный тип - гем B; другие важные типы включают гем А и гем C. Изолированные гемы обычно обозначаются заглавными буквами, а гемы, связанные с белками, обозначаются строчными буквами. Цитохром а относится к гему А в специфической комбинации с мембранным белком, образующим часть цитохром с оксидаза.[16]
Другие рубцы
- Следующая система углеродной нумерации порфиринов - это более старая нумерация, используемая биохимиками, а не система нумерации 1-24, рекомендованная ИЮПАК который показан в таблице выше.
- Гем л является производным гема B, который ковалентно присоединен к белку лактопероксидаза, эозинофильная пероксидаза, и пероксидаза щитовидной железы. Добавление перекись с глутамил -375 и аспартил -225 лактопероксидазы образует сложноэфирные связи между этими аминокислотными остатками и 1- и 5-метильными группами гема соответственно.[17] Считается, что аналогичные сложноэфирные связи с этими двумя метильными группами образуются в эозинофильных и тироидных пероксидазах. Гем л является одной из важных характеристик пероксидаз животных; пероксидазы растений включают гем B. Лактопероксидаза и пероксидаза эозинофилов являются защитными ферментами, ответственными за разрушение вторгающихся бактерий и вирусов. Пероксидаза щитовидной железы - это фермент, катализирующий биосинтез важных гормонов щитовидной железы. Поскольку лактопероксидаза уничтожает вторгшиеся в легкие и экскременты организмы, считается, что она является важным защитным ферментом.[18]
- Гем м производное гема B, ковалентно связанное с активным центром перекись. Гем м содержит два сложноэфирные связи у гема 1- и 5-метил, как в геме л содержится в пероксидах других млекопитающих. Кроме того, уникальный сульфонамид ионная связь между серой остатка метиониламинокислоты и гем-2-виниловой группой, придает этому ферменту уникальную способность легко окисляться хлористый и бромид ионы. Миелопероксидаза присутствует у млекопитающих нейтрофилы и отвечает за уничтожение вторгающихся бактерий и вирусов. Он также синтезирует гипобромит по ошибке". Гипохлорит и гипобромит вызывают образование галогенированных нуклеозидов, которые являются мутагенными соединениями.[19][20]
- Heme D является другим производным гема B, но в котором пропионовая кислота боковая цепь при атоме углерода в положении 6, который также гидроксилирован, образует γ-спиролактон. Кольцо III также гидроксилировано в положении 5 в конформации транс к новой группе лактонов.[21] Гем D является местом восстановления кислорода в воде многих видов бактерий при низком давлении кислорода.[22]
- Heme S связан с гемом B наличием формальный группа в положении 2 вместо 2-винильной группы. Гем S содержится в гемоглобине морских червей. Правильные структуры гема B и гема S были впервые выяснены немецким химиком. Ганс Фишер.[23]
Имена цитохромы обычно (но не всегда) отражают типы гемов, которые они содержат: цитохром a содержит гем A, цитохром c содержит гем C и т. д. Это соглашение, возможно, было впервые введено с публикацией структуры гем А.
Использование заглавных букв для обозначения типа гема
Практика обозначения гема заглавными буквами была формализована в сноске в статье Пуустинена и Викстрома.[24] который объясняет, при каких условиях следует использовать заглавную букву: «мы предпочитаем использовать заглавные буквы для описания структуры гема как изолированной. Строчные буквы могут затем свободно использоваться для цитохромов и ферментов, а также для описания отдельного гема, связанного с белками. группы (например, комплексы цитохрома bc и aa3, цитохром b5, гем c1 до н.э.1 комплекс, гем А3 из аа3 комплекс и т. д.) ». Другими словами, химическое соединение будет обозначаться заглавной буквой, но конкретные примеры в структурах - строчными буквами. Таким образом, цитохромоксидаза, которая имеет два гема А (гем а и гем а3) в своей структуре содержит два моля гема А на моль белка. Цитохром bc1, с рубцами bЧАС, бL, и c1, содержит гем B и гем C в соотношении 2: 1. Эта практика, по-видимому, зародилась в статье Коуги и Йорка, в которой продукт новой процедуры выделения гема цитохрома аа3 был обозначен как гем А, чтобы отличить его от предыдущих препаратов: «Наш продукт не во всех отношениях идентичен гему цитохрома аа3. гем А, полученный в растворе другими исследователями путем восстановления гемина А, выделенного ранее (2). По этой причине мы будем обозначать наш продукт гем А, пока очевидные различия не будут объяснены ».[25] В более поздней статье[26] Группа Коши использует заглавные буквы для обозначения изолированного гема B и C, а также для A.
Синтез

Ферментативный процесс, производящий гем, правильно называется порфирин синтез, поскольку все промежуточные соединения тетрапирролы которые химически классифицируются как порфирины. Этот процесс очень консервативен в биологии. У людей этот путь служит почти исключительно для образования гема. В бактерии, он также производит более сложные вещества, такие как кофактор F430 и кобаламин (витамин B12 ).[27]
Путь инициируется синтезом δ-аминолевулиновая кислота (dALA или δALA) от аминокислота глицин и сукцинил-КоА от цикл лимонной кислоты (Цикл Кребса). Ограничивающий скорость фермент, ответственный за эту реакцию, ALA-синтаза, отрицательно регулируется концентрацией глюкозы и гема. Механизм ингибирования ALA гемом или гемином заключается в снижении стабильности синтеза мРНК и уменьшении поступления мРНК в митохондрии. Этот механизм имеет терапевтическое значение: вливание гем аргинат или же гематин и глюкоза может купировать приступы острая перемежающаяся порфирия у пациентов с врожденная ошибка обмена веществ этого процесса, уменьшая транскрипцию синтазы ALA.[28]
Органы, в основном участвующие в синтезе гема, - это печень (в котором скорость синтеза сильно варьируется в зависимости от системного пула гема) и Костный мозг (в котором скорость синтеза гема относительно постоянна и зависит от производства цепи глобина), хотя каждой клетке необходим гем для правильного функционирования. Однако из-за своих токсичных свойств белки, такие как Гемопексин (Hx) необходимы для поддержания физиологических запасов железа, чтобы его можно было использовать в синтезе.[29] Гем рассматривается как промежуточная молекула в катаболизме гемоглобина в процессе метаболизм билирубина. Дефекты в различных ферментах синтеза гема могут привести к группе расстройств, называемых порфириями, к ним относятся: острая перемежающаяся порфирия, врожденная эритропоэтическая порфирия, поздняя кожная порфирия, наследственная копропорфирия, пестрая порфирия, эритропоэтическая протопорфирия.[30][нужна цитата ]
Синтез для еды
Невозможная еда, производители растительной заменитель мяса, используйте ускоренный процесс синтеза гема с участием корня сои леггемоглобин и дрожжи, добавляя полученный гем к таким предметам, как постные (веган ) Невозможные котлеты для бургеров. ДНК для леггемоглобин продукцию экстрагировали из клубеньков корня сои и экспрессировали в дрожжевых клетках, чтобы получить избыточное производство гема для использования в бургерах без мяса.[31] Этот процесс утверждает, что в получаемых продуктах создается мясной вкус.[32][33]
Деградация
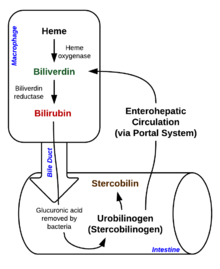
Деградация начинается внутри макрофагов селезенка, которые удаляют старые и поврежденные эритроциты из кровотока. На первом этапе гем преобразуется в биливердин ферментом гемоксигеназа (HO).[34] НАДФН используется как восстановитель, в реакцию вступает молекулярный кислород, монооксид углерода (CO) образуется, и железо высвобождается из молекулы в виде железо ион (Fe2+).[35] CO действует как клеточный посредник и участвует в расширении сосудов.[36]
Кроме того, деградация гема, по-видимому, является эволюционно законсервированной реакцией на окислительный стресс. Вкратце, когда клетки подвергаются воздействию свободные радикалы, происходит быстрая индукция экспрессии стресс-чувствительных гемоксигеназа-1 (HMOX1) изофермент, который катаболизирует гем (см. Ниже).[37] Причина, по которой клетки должны экспоненциально увеличивать свою способность разлагать гем в ответ на окислительный стресс, остается неясной, но, по-видимому, это часть цитопротекторного ответа, который позволяет избежать вредного воздействия свободного гема. Когда накапливается большое количество свободного гема, системы детоксикации / деградации гема перегружаются, что позволяет гему оказывать свое разрушающее действие.[29]
| гем | гемоксигеназа-1 | биливердин + Fe2+ | |
 |  | ||
| ЧАС+ + НАДФН + O2 | НАДФ+ + CO | ||
Во второй реакции биливердин конвертируется в билирубин к биливердин редуктаза (BVR):[38]
| биливердин | биливердин редуктаза | билирубин | |
|  | ||
| ЧАС+ + НАДФН | НАДФ+ | ||
Билирубин транспортируется в печень путем облегченной диффузии, связываясь с белком (сывороточный альбумин ), где он сопряжен с глюкуроновая кислота стать более растворимыми в воде. Реакция катализируется ферментом UDP-глюкуронозилтрансфераза.[39]
| билирубин | UDP-глюкуронозилтрансфераза | билирубин диглюкуронид | |
|  | ||
| 2 UDP-глюкуронид | 2 UMP + 2 пя | ||
Эта форма билирубина выводится из печени в желчь. Экскреция билирубина из печени в желчные каналы - это активный, энергозависимый и ограничивающий скорость процесс. В кишечные бактерии деконъюгировать билирубин диглюкуронид и преобразовать билирубин в уробилиногены. Некоторое количество уробилиногена всасывается клетками кишечника и транспортируется в почки и выводится с моча (уробилин, который является продуктом окисления уробилиногена и отвечает за желтый цвет мочи). Остаток проходит по пищеварительному тракту и превращается в стеркобилиноген. Это окисляется до стеркобилин, который выводится из организма и отвечает за коричневый цвет кал.[40]
В здоровье и болезни
Под гомеостаз реактивность гема контролируется его внедрением в «гемовые карманы» гемопротеинов.[нужна цитата ] Однако при окислительном стрессе некоторые гемопротеины, например гемоглобин, может высвобождать их гем простетические группы.[41][42] Не связанный с белками (свободный) гем, полученный таким образом, становится очень цитотоксичным, скорее всего, из-за атома железа, содержащегося в его кольце протопорфирина IX, который может действовать как Реактив Фентона неограниченно катализировать образование свободных радикалов.[43] Он катализирует окисление и агрегацию белка, образование цитотоксической перекиси липидов посредством перекисного окисления липидов и повреждает ДНК из-за окислительного стресса. Благодаря своим липофильным свойствам он нарушает липидные бислои в органеллах, таких как митохондрии и ядра.[44] Эти свойства свободного гема могут повышать чувствительность различных типов клеток. запрограммированная гибель клеток в ответ на провоспалительные агонисты, вредный эффект, который играет важную роль в патогенезе некоторых воспалительных заболеваний, таких как малярия[45] и сепсис.[46] Существует связь между высоким потреблением гемового железа, полученного из мяса, и повышенным риском рак толстой кишки.[47] Содержание гема в красном мясе в 10 раз выше, чем в белом мясе, таком как курица.[48]
Гены
Следующие гены являются частью химического пути образования гема:
- ALAD: аминолевулиновая кислота, δ-, дегидратаза (дефицит вызывает порфирию с дефицитом ала-дегидратазы)[49]
- ALAS1: аминолевулинат, δ-, синтаза 1
- ALAS2: аминолевулинат, δ-, синтаза 2 (дефицит вызывает сидеробластную / гипохромную анемию)
- CPOX: копропорфириноген оксидаза (дефицит вызывает наследственную копропорфирию)[50]
- FECH: феррохелатаза (протопорфирия)
- HMBS: гидроксиметилбилан синтаза (дефицит вызывает острую перемежающуюся порфирию)[51]
- PPOX: протопорфириноген оксидаза (дефицит вызывает пеструю порфирию)[52]
- UROD: уропорфириноген декарбоксилаза (дефицит вызывает позднюю кожную порфирию)[53]
- UROS: уропорфириноген III синтаза (дефицит вызывает врожденную эритропоэтическую порфирию)
Примечания и ссылки
- ^ "Определение Гема". www.merriam-webster.com. Получено 2020-01-26.
- ^ Блумер, Джозеф Р. (1998). «Печеночный метаболизм порфиринов и гема». Журнал гастроэнтерологии и гепатологии. 13 (3): 324–329. Дои:10.1111 / j.1440-1746.1998.01548.x. PMID 9570250.
- ^ Химия, Международный союз теоретических и прикладных наук (2009). «Гемы (производные гема)». Сборник химической терминологии ИЮПАК. iupac.org. ИЮПАК. Дои:10.1351 / goldbook.H02773. ISBN 978-0-9678550-9-7. В архиве из оригинала 22 августа 2017 г.. Получено 28 апреля 2018.
- ^ Стандартный текст по биохимии определяет гем как «железопорфириновую простетическую группу гемовых белков» (Nelson, D. L .; Cox, M. M. «Lehninger, Principles of Biochemistry», 3-е изд. Стоит опубликовать: New York, 2000. ISBN 1-57259-153-6.)
- ^ Поулос, Томас Л. (9 апреля 2014 г.). «Структура и функция гемового фермента». Химические обзоры. 114 (7): 3919–3962. Дои:10.1021 / cr400415k. ISSN 0009-2665. ЧВК 3981943. PMID 24400737.
- ^ Паоли, М. (2002). «Структурно-функциональные отношения в гем-белках». ДНК клетки биол. 21 (4): 271–280. Дои:10.1089/104454902753759690. PMID 12042067.
- ^ Олдертон, В. (2001). «Синтазы оксида азота: структура, функции и ингибирование». Biochem. J. 357 (3): 593–615. Дои:10.1042 / bj3570593. ЧВК 1221991. PMID 11463332.
- ^ Милани, М. (2005). «Структурные основы связывания гема и распознавания двухатомных лигандов в усеченных гемоглобинах». J. Inorg. Биохим. 99 (1): 97–109. Дои:10.1016 / j.jinorgbio.2004.10.035. PMID 15598494.
- ^ Хардисон, Р. (1999). «Эволюция гемоглобина: исследования очень древнего белка показывают, что изменения в регуляции генов являются важной частью эволюционной истории». Американский ученый. 87 (2): 126. Дои:10.1511/1999.20.809.
- ^ Поулос, Т. (2014). «Структура и функция гемового фермента». Chem. Rev. 114 (7): 3919–3962. Дои:10.1021 / cr400415k. ЧВК 3981943. PMID 24400737.
- ^ Том, С. С. (2013). «Варианты гемоглобина: биохимические свойства и клинические корреляты». Холодный источник Харб. Перспектива. Med. 3 (3): a011858. Дои:10.1101 / cshperspect.a011858. ЧВК 3579210. PMID 23388674.
- ^ Бор; Hasselbalch, Krogh. «О биологически важной взаимосвязи - влиянии содержания углекислого газа в крови на связывание с кислородом». В архиве из оригинала от 18.04.2017. Цитировать журнал требует
| журнал =(помощь) - ^ Ackers, G.K .; Холт, Дж. М. (2006). «Асимметричная кооперативность в симметричном тетрамере: гемоглобин человека». J. Biol. Chem. 281 (17): 11441–3. Дои:10.1074 / jbc.r500019200. PMID 16423822.
- ^ Caughey, W. S .; Смайт, Г. Э .; O'Keeffe, D.H .; Маскаски, Дж. Э .; Смит, М. Л. (1975). «Гем A цитохром с оксидазы: структура и свойства: сравнение с гемами B, C и S и производными». J. Biol. Chem. 250 (19): 7602–7622. PMID 170266.
- ^ Хегг, Эрик Л. (2004). «Гем А-синтаза не включает молекулярный кислород в формильную группу гема А». Биохимия. 43 (27): 8616–8624. Дои:10.1021 / bi049056m. PMID 15236569.
- ^ Йошикава, С. (2012). «Структурные исследования цитохром с оксидазы сердца крупного рогатого скота». Биохим. Биофиз. Acta. 1817 (4): 579–589. Дои:10.1016 / j.bbabio.2011.12.012. PMID 22236806.
- ^ Rae, T .; Гофф, Х. (1998). «Гем простетическая группа лактопероксидазы. Структурные характеристики гема I и гем-I-пептидов». Журнал биологической химии. 273 (43): 27968–27977. Дои:10.1074 / jbc.273.43.27968. PMID 9774411.
- ^ Парди, М.А. (1983). «Влияние фазы роста и структуры клеточной оболочки на восприимчивость триумфальных сальмонелл к системе лактопероксидаза-тиоцианат-пероксид водорода». Заразить. Иммунная. 39 (3): 1187–95. ЧВК 348082. PMID 6341231.
- ^ Охима, Х. (2003). «Химические основы канцерогенеза, вызванного воспалением». Arch. Biochem. Биофизы. 417 (1): 3–11. Дои:10.1016 / с0003-9861 (03) 00283-2. PMID 12921773.
- ^ Хендерсон, Дж. П. (2003). «Фагоциты продуцируют 5-хлороурацил и 5-бромурацил, два мутагенных продукта миелопероксидазы, в воспалительной ткани человека». J. Biol. Chem. 278 (26): 23522–8. Дои:10.1074 / jbc.m303928200. PMID 12707270.
- ^ Муршудов, Г .; Гребенко, А .; Барынин, В .; Dauter, Z .; Wilson, K .; Вайнштейн, Б .; Мелик-Адамян, З .; Браво, Дж .; Ferrán, J .; Ferrer, J.C .; Switala, J .; Loewen, P.C .; Фита, И. (1996). «Структура гема d из Penicillium vitale и кишечная палочка каталазы " (pdf). Журнал биологической химии. 271 (15): 8863–8868. Дои:10.1074 / jbc.271.15.8863. PMID 8621527.
- ^ Белевич, И. (2005). «Оксигенированный комплекс цитохрома bd из Escherichia coli: стабильность и фотолабильность». Письма FEBS. 579 (21): 4567–70. Дои:10.1016 / j.febslet.2005.07.011. PMID 16087180.
- ^ Fischer, H .; Орт, Х. (1934). Die Chemie des Pyrrols. Лиепциг: Ischemia Verlagsgesellschaft.
- ^ Пуустинен А., Викстрём М. (1991). «Гемовые группы цитохрома o из Escherichia coli». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 88 (14): 6122–6. Bibcode:1991ПНАС ... 88.6122П. Дои:10.1073 / pnas.88.14.6122. ЧВК 52034. PMID 2068092.
- ^ Caughey WS, York JL (1962). «Выделение и некоторые свойства зеленого гема цитохромоксидазы из сердечной мышцы говядины». J. Biol. Chem. 237: 2414–6. PMID 13877421.
- ^ Caughey WS, Smythe GA, O'Keeffe DH, Maskasky JE, Smith ML (1975). «Гем А цитохром с оксидазы. Структура и свойства: сравнение с гемами В, С, S и производными». J. Biol. Chem. 250 (19): 7602–22. PMID 170266.
- ^ Баттерсби, Алан Р. (2000). «Тетрапирролы: пигменты жизни». Отчеты о натуральных продуктах. 17 (6): 507–526. Дои:10.1039 / B002635M. PMID 11152419.
- ^ Шридеви, Коллури (28 апреля 2018 г.). Повышающая регуляция фермента пути гема АЛК-синтаза-1 глутетимидом и 4,6-диоксогептановой кислотой и понижающая регуляция глюкозой и гемом: диссертация. Стипендия ES @ UMMS (Тезис). Медицинская школа Массачусетского университета. Дои:10.13028 / yyrz-qa79. В архиве из оригинала от 8 августа 2016 г.. Получено 28 апреля 2018.
- ^ а б Кумар, Санджай; Bandyopadhyay, Удай (июль 2005 г.). «Токсичность свободного гема и системы его детоксикации у человека». Письма токсикологии. 157 (3): 175–188. Дои:10.1016 / j.toxlet.2005.03.004. PMID 15917143.
- ^ Пюи, Эрве; Гуя, Лоран; Дейбах, Жан-Шарль (март 2010 г.). "Порфирии". Ланцет. 375 (9718): 924–937. Дои:10.1016 / S0140-6736 (09) 61925-5.
- ^ Фрейзер, Рэйчел З .; Шиту, Митхила; Агравал, пуджа; Мендес, Одете; Клапхольц, Сью (2018-04-11). «Оценка безопасности препарата соевого леггемоглобина, полученного из Pichia pastoris, предназначенного для использования в качестве вкусового катализатора в мясе на растительной основе». Международный журнал токсикологии. 37 (3): 241–262. Дои:10.1177/1091581818766318. ISSN 1091-5818. ЧВК 5956568. PMID 29642729.
- ^ "Внутри странной науки о фальшивом мясе, которое кровоточит'". Проводной. 2017-09-20. В архиве из оригинала 24 марта 2018 г.. Получено 28 апреля 2018.
- ^ «Кремниевая долина пробуждает вкус к еде». Экономист. 2015-03-05. ISSN 0013-0613. Получено 2019-04-08.
- ^ Мейнс, Махин Д. (июль 1988 г.). «Гемоксигеназа: функция, множественность, регуляторные механизмы и клиническое применение». Журнал FASEB. 2 (10): 2557–2568. Дои:10.1096 / fasebj.2.10.3290025. ISSN 0892-6638.
- ^ Принципы биохимии Ленингера (5-е изд.). Нью-Йорк: В. Х. Фриман и компания. 2008. С.876. ISBN 978-0-7167-7108-1.
- ^ Ханафи, К.А. (2013). «Угарный газ и мозг: время переосмыслить догму». Curr. Pharm. Des. 19 (15): 2771–5. Дои:10.2174/1381612811319150013. ЧВК 3672861. PMID 23092321.
- ^ Abraham, N.G .; Каппас, А. (2008). «Фармакологические и клинические аспекты гемоксигеназы». Pharmacol. Rev. 60 (1): 79–127. Дои:10.1124 / пр.107.07104. PMID 18323402.
- ^ Florczyk, U.M .; Jozkowicz, A .; Дулак, Дж. (Январь – февраль 2008 г.). «Биливердинредуктаза: новые свойства старого фермента и его потенциальное терапевтическое значение». Фармакологические отчеты. 60 (1): 38–48. ЧВК 5536200. PMID 18276984.
- ^ King, C .; Rios, G .; Грин, М .; Тефли Т. (2000). «УДФ-глюкуронозилтрансферазы». Текущий метаболизм лекарств. 1 (2): 143–161. Дои:10.2174/1389200003339171. PMID 11465080.
- ^ Хельменстин, Энн Мари. «Химические вещества, отвечающие за цвет мочи и кала». ThoughtCo. Получено 2020-01-24.
- ^ Bunn, H.F .; Джандл, Дж. Х. (сентябрь 1966 г.). «Обмен гема между молекулами гемоглобина». Proc. Natl. Акад. Sci. Соединенные Штаты Америки. 56 (3): 974–978. Bibcode:1966ПНАС ... 56..974Б. Дои:10.1073 / пнас.56.3.974. ЧВК 219955. PMID 5230192.
- ^ Smith, M. L .; Paul, J .; Ohlsson, P. I .; Hjortsberg, K .; Пол, К. Г. (февраль 1991 г.). «Деление гем-белка в неденатурирующих условиях». Proc. Natl. Акад. Sci. Соединенные Штаты Америки. 88 (3): 882–886. Bibcode:1991ПНАС ... 88..882С. Дои:10.1073 / pnas.88.3.882. ЧВК 50918. PMID 1846966.
- ^ Everse, J .; Ся, Н. (1197). «Токсичность нативных и модифицированных гемоглобинов». Свободная радикальная биология и медицина. 22 (6): 1075–1099. Дои:10.1016 / S0891-5849 (96) 00499-6. PMID 9034247.
- ^ Кумар, Санджай; Bandyopadhyay, Удай (июль 2005 г.). «Токсичность свободного гема и его системы детоксикации у человека». Письма токсикологии. 157 (3): 175–188. Дои:10.1016 / j.toxlet.2005.03.004. PMID 15917143.
- ^ Памплона, А .; Ferreira, A .; Balla, J .; Джени, В .; Balla, G .; Epiphanio, S .; Chora, A .; Rodrigues, C.D .; Gregoire, I.P .; Cunha-Rodrigues, M .; Португалия, S .; Soares, M. P .; Мота, М. М. (июнь 2007 г.). «Гемоксигеназа-1 и окись углерода подавляют патогенез экспериментальной церебральной малярии». Природа Медицина. 13 (6): 703–710. Дои:10,1038 / нм1586. PMID 17496899.
- ^ Larsen, R .; Gozzelino, R .; Джени, В .; Tokaji, L .; Bozza, F.A .; Japiassú, A.M .; Бонапарт, Д .; Кавальканте, М. М .; Chora, A .; Ferreira, A .; Маргути, I .; Cardoso, S .; Sepúlveda, N .; Smith, A .; Соарес, М. П. (2010). «Центральная роль свободного гема в патогенезе тяжелого сепсиса». Научная трансляционная медицина. 2 (51): 51ra71. Дои:10.1126 / scitranslmed.3001118. PMID 20881280.
- ^ Bastide, N.M .; Pierre, F. H .; Корпет, Д. Э. (2011). «Гемовое железо из мяса и риск колоректального рака: метаанализ и обзор задействованных механизмов» (PDF). Рак Пред. Res. 4 (2): 177–184. Дои:10.1158 / 1940-6207.CAPR-10-0113. PMID 21209396. В архиве (PDF) из оригинала от 25.09.2015.
- ^ Bastide, Nadia M .; Pierre, Fabrice H.F .; Корпет, Денис Э. (1 февраля 2011 г.). «Гемовое железо из мяса и риск колоректального рака: метаанализ и обзор задействованных механизмов». Исследования по профилактике рака. 4 (2): 177–184. Дои:10.1158 / 1940-6207.CAPR-10-0113. PMID 21209396. В архиве из оригинала 11 октября 2017 г.. Получено 28 апреля 2018 - через Cancepreventionresearch.aacrjournals.org.
- ^ Плевинская, Магдалена; Тунелл, Стиг; Холмберг, Ларс; Ветмур, Джеймс; Десник, Роберт (1991). «Порфирия с дефицитом дельта-аминолевулинатдегидратазы: идентификация молекулярных повреждений в сильно пораженной гомозиготе». Американский журнал генетики человека. 49 (1): 167–174. ЧВК 1683193. PMID 2063868.
- ^ Aurizi, C .; Lupia Palmieri, G .; Barbieri, L .; Macri, A .; Sorge, F .; Usai, G .; Биолкати, Г. (февраль 2009 г.). «Четыре новые мутации гена оксидазы копропорфириноген III». Клеточная и молекулярная биология. 55 (1): 8–15.
- ^ Bustad, H.J .; Vorland, M .; Ronneseth, E .; Sandberg, S .; Martinez, A .; Тоска, К. (8 августа 2013 г.). «Анализ конформационной стабильности и активности двух мутантов гидроксиметилбилансинтазы, K132N и V215E, с различной фенотипической ассоциацией с острой перемежающейся порфирией». Отчеты по бионауке. 33 (4): 617–626. Дои:10.1042 / BSR20130045. ЧВК 3738108. PMID 23815679.
- ^ Мартинес ди Монтемурос, Ф .; Di Pierro, E .; Patti, E .; Tavazzi, D .; Danielli, M. G .; Biolcati, G .; Rocchi, E .; Капплини, М. Д. (декабрь 2002 г.). «Молекулярная характеристика порфирий в Италии: диагностическая блок-схема». 48 (8): 867–876. Цитировать журнал требует
| журнал =(помощь) - ^ Баденас, С .; Фигерасу, Дж .; Phillips, J.D .; Warby, C.A .; Muñoz, C .; Эрреро, К. (апрель 2009 г.). «Идентификация и характеристика новых мутаций гена уропорфириногендекарбоксилазы в большой серии пациентов и родственников поздней кожной порфирии». Клиническая генетика. 75 (4): 346–353. Дои:10.1111 / j.1399-0004.2009.01153.x. ЧВК 3804340. PMID 19419417.