Внешняя морфология Lepidoptera - External morphology of Lepidoptera






В внешняя морфология Lepidoptera это физиологическая структура тел насекомые принадлежащий к порядку Чешуекрылые, также известный как бабочки и моль. Lepidoptera отличаются от других отрядов наличием напольные весы на внешних частях тела и придатках, особенно на крыльях. Бабочки и мотыльки различаются по размеру от микролепидоптеры всего несколько миллиметров в длину, с размахом крыльев в несколько дюймов, как у Атласский мотылек. Состоящие из более чем 160 000 описанных видов чешуекрылые обладают вариациями основной структуры тела, которые эволюционировали, чтобы получить преимущества в адаптации и распространении.[1]
Чешуекрылые подвергаются полная метаморфоза, проходя четырехступенчатую жизненный цикл: яйцо, личинка или же гусеница, куколка или же куколка, и имаго (множественное число: воображает) / взрослый. Личинки - гусеницы - имеют закаленную (склеротизированный ) головная капсула, жевательная ротовые органы, и мягкое тело, которое может иметь волосовидные или другие выступы, три пары настоящие ноги, и до пяти пар бездельники. Большинство гусениц травоядные животные, но некоторые из них плотоядные животные (некоторые едят муравьев, тлей или других гусениц) или детритофаги.[2] Личинки находятся на стадии питания и роста и периодически подвергаются гормонально-индуцированной шелушение, развиваясь с каждым возраст, пока они не пройдут заключительную личиночно-кукольную линьку. Личинки многих видов чешуекрылых могут сделать шелковую оболочку, называемую кокон и окукливаться внутри него или окукливается в клетке под землей. У многих бабочек куколка подвешена к кремастер и называется куколка.
Тело взрослого человека окаменело. экзоскелет за исключением живота, который менее склеротизирован. Голова имеет форму капсулы с отходящими от нее придатками. Ротовой аппарат взрослых включает заметные хоботок сформированный из верхней челюсти Galeae, приспособлены для сосания нектара. Некоторые виды не питаются во взрослом состоянии и могут иметь уменьшенный ротовой аппарат, в то время как другие модифицировали их для прокалывания и сосания крови или фруктовых соков.[3] Челюсти отсутствуют во всех, кроме Micropterigidae который имеет жевательные аппараты.[4] У взрослых чешуекрылых есть два неподвижных многогранных сложные глаза, и только два простые глаза или глазки, которые могут быть уменьшены.[5] Три сегмента грудная клетка сливаются вместе. Антенны являются заметными и, помимо способности обоняния, также помогают в навигации, ориентации и равновесии во время полета.[6] У бабочек у самцов часто больше перистых антенн, чем у самок, для обнаружения самок. феромоны На расстоянии. Есть две пары перепончатый крылья которые возникают из среднегрудного (среднего) и заднегрудного (третьего) сегментов; они обычно полностью покрываются минутой напольные весы. Два крыла с каждой стороны действуют как одно в силу механизмы блокировки крыльев. В некоторых группах самки нелетают и имеют уменьшенные крылья. В брюшной полости десять сегментов, соединенных подвижными межсегментарными перепонками. Последние сегменты брюшка образуют наружный гениталии. Гениталии сложны и служат основой для идентификации семьи и различения видов.[7]
Крылья, головные части грудной клетки и брюшко чешуекрылых покрыты мелкими чешуйками, от которых отряд чешуекрылых получил свое название, слово лепид в Древнегреческий что означает «масштаб». Большинство чешуек являются пластинчатыми (пластинчатыми) и прикрепляются к ножке, в то время как другие формы могут быть похожи на волосы или специализироваться как вторичные половые признаки. Просвет, или поверхность ламели, имеет сложное строение. Цвет придает либо за счет пигменты содержащиеся в нем или через его трехмерную структуру.[8] Весы предоставляют ряд функций, в том числе: изоляция, терморегуляция, а также помощь в полете, среди прочего, наиболее важным из которых является большое разнообразие ярких или нечетких рисунков, которые они обеспечивают, которые помогают организму защитить себя камуфляж, мимикрия, и искать себе пару.
Внешняя морфология

Как и другие участники суперзаказ Холометабола, Lepidoptera проходят полный метаморфоза, проходящий четырехступенчатый жизненный цикл: яйцо, личинка / гусеница, куколка / куколка и имаго (множественное число: воображает) / взрослый.[9]
Размер чешуекрылых варьируется от нескольких миллиметров в длину, как в случае с микролепидоптерами, до размаха крыльев в несколько дюймов, например, у чешуекрылых. Атласский мотылек и самая большая бабочка в мире Птичье крыло королевы Александры.[10]:246
Общий план тела
Тело взрослой бабочки или моли (имаго) состоит из трех отдельных частей, называемых тагмата, связанные при перетяжках; эти тагматы - голова, грудная клетка и живот. У взрослых чешуекрылых четыре крыла - переднее и заднее на левой и правой стороне грудной клетки - и, как у всех насекомых, три пары ног.[11]
В морфологический Характеристики, которые отличают отряд Lepidoptera от других отрядов насекомых:[10]:246
- Голова: На голове большие сложные глаза, и, если присутствует ротовой аппарат, это почти всегда хоботок, похожий на соломинку для питья.
- Напольные весы: Чешуя покрывает внешнюю поверхность тела и придатков.
- Грудная клетка: The переднегрудь обычно снижается.
- Крылья: Две пары крыльев есть почти у всех таксонов. Крыльев очень мало поперечные вены.
- Брюшная полость: Задние сегменты брюшной полости сильно модифицированы для воспроизводства. Cerci отсутствуют.
- Личинка: Личинки чешуекрылых известны как гусеницы, у них хорошо развиты голова и челюсти. У них может быть от нуля до пяти пар бездельники, обычно четыре.
- Куколка: Куколки у большинства видов адектический (без функциональных нижних челюстей в куколочном состоянии) и обнаруживать (с отростками, сросшимися или приклеенными к телу), а другие - сладкий (с функциональными нижними челюстями, присутствующими в кукольном состоянии) и исключать (без усиков, лап и крыльев).
Отличительные таксономические признаки
Основные характеристики, используемые для классификации видов, родов и семейств чешуекрылых:[12]
- ротовой аппарат
- форма и жилкование крыльев
- являются ли крылья однородными (жилкование передних и задних крыльев одинаково) или неоднородными (передние и задние крылья разные)
- являются ли крылья заостренными (более или менее покрытыми специальными щетинками, называемыми микросетками) или неакулеатными
- тип сочленения крыла (югатное или френатное)
- анатомия репродуктивных органов
- строение личинки и положение первичных щетинок
- независимо от того, является ли куколка удаленной или запакованной
Морфологические характеристики гусениц и куколок, используемые для классификации, полностью отличаются от таковых взрослых;[13]:637[14] различные схемы классификации иногда предоставляются отдельно для классификации взрослых особей, личинок и куколок.[14][15]:28–40 Характеристики незрелых стадий все чаще используются в таксономических целях, поскольку они дают представление о систематике и филогении чешуекрылых, которые не очевидны при осмотре взрослых особей.[15]:28
Голова

Как и все головы животных, голова бабочки или моли содержит органы питания и основные органы чувств. Голова обычно состоит из двух антенн, двух сложных глаз, двух пальпи, и хоботок.[11] Чешуекрылые имеют глазки которые могут быть или не быть видимыми. У них также есть сенсорные структуры, называемые хетосема, функции которых практически неизвестны. Голова в основном заполнена мозгом, всасывающим насосом и связанными с ним мышечными пучками.[16] В отличие от взрослых, личинки имеют одночлениковые мандибулы.[16]
Головная капсула хорошо склеротизирована и имеет ряд склериты или тарелки, разделенные швы. Склериты трудно отличить от борозды (единственное число - борозда ), которые являются вторичными утолщениями. Области головы были разделены на несколько областей, которые служат топографическим ориентиром для описания лепидоптерологами, но не могут быть различимы с точки зрения их развития.[16] Голова покрыта волосковидными или пластинчатыми чешуйками и находится либо в виде пучков на голове, либо лоб или же вершина (именуемый грубый) или прижат к голове (называемый сглаженный).
Органы чувств и структуры на голове очень разнообразны, и форма и форма этих структур, а также их наличие или отсутствие являются важными таксономическими показателями для классификации таксонов по семействам.[13]

Глава семейства бабочек Gracillariidae показывая степень чешуи на голове

Голова бабочки с грубой чешуей Monopis icterogastra (семья Tineidae )

Голова бабочки с гладкой чешуей Glyphipterix simpliciella (семья Glyphipterigidae )

Голова бабочки с гладкой чешуей Stegasta variana (семья Gelechiidae )




Антенны


Усики - это выступающие парные отростки, которые выступают вперед между глазами животного и состоят из нескольких сегментов. У бабочек их длина колеблется от половины длины переднего крыла до трех четвертей длины переднего крыла. Усики бабочек либо тонкие, с выступами на конце, а в случае Hesperiidae, зацеплены за кончик. У некоторых родов бабочек, таких как Либитея и Тарактротера ручка внизу полая.[11] Усики бабочки бывают нитевидный (нить нравится), однотонный (как гребешок), двояковыпуклый (как перо), крючковатая, булавовидная или утолщенная.[13]:636 Бомбикс мандарина пример с двояковыпуклыми усиками.[17] У некоторых бабочек есть бугорчатые усики, похожие на антенны бабочек, в том числе у семейств Castniidae, Neocastniidae, и Euschemonidae.[18]
Усики - главные органы обоняние (запах) у чешуекрылых. Поверхность антенны покрыта большим количеством обонятельных чешуек, волосков или ямок; целых 1,370,000 обнаружены на антеннах монарх. Усики чрезвычайно чувствительны; пернатые усики самцов моли из Saturniidae, Lasiocampidae и многих других семейств настолько чувствительны, что могут обнаруживать феромоны самок бабочек на расстоянии до 2 км (1,2 мили). Усики чешуекрылых можно наклонять во многих положениях. Они помогают насекомому обнаружить запах и могут рассматриваться как своего рода «обонятельный радар».[6] У бабочек у самцов часто есть более перистые усики, чем у самок, для обнаружения женских феромонов на расстоянии.[7] Поскольку самкам не нужно обнаруживать самцов, у них более простые усики.[6] Было также обнаружено, что антенны играют роль в компенсированной по времени солнечный компас ориентация в мигрирующие бабочки-монархи.[19]

Нитевидные усики - Eriocrania cicatricella (Eriocraniidae ) 
Одноядерные усики - Abantiades barcas (Hepialidae ) 
Двояковидные усики - Actias Artemis (Saturniidae ) 
Крючковые усики - Epargyreus clarus (Hesperiidae ) 
Усики булавовидные - Ванесса аталанта (Нимфалиды ) 
Утолщенные усики - Делейфила эльпенор (Сфингиды ) 
Усики моли булавоватые - Athis inca (Castniidae )








Глаза


У чешуекрылых есть два больших неподвижных сложных глаза, состоящих из большого количества грани или линзы, каждая из которых соединена с линзовидным цилиндром, который прикреплен к нерву, ведущему к мозг.[11] Каждый глаз может иметь до 17000 индивидуальных световых рецепторов (омматидия ), которые в совокупности обеспечивают широкий мозаичный вид на окрестности.[6] Одна тропическая азиатская семья, Amphitheridae, имеет сложные глаза, разделенные на два отчетливых сегмента.[13][20] Глаза обычно гладкие, но могут быть покрыты мелкими волосками. Глаза бабочек обычно коричневые, золотисто-коричневые или даже красные, как у некоторых видов бабочек. капитаны.[11]
Хотя у большинства насекомых есть три простых глаза, или глазки только два глазка присутствуют у всех видов чешуекрылых, за исключением нескольких бабочек, по одному с каждой стороны головы у края сложного глаза. У некоторых видов органы чувств, называемые хетосема находятся около глазков.[11][21] Глазки не гомологичный к простым глазам гусениц, которые иначе называются стволовые.[5] Глазки чешуекрылых в некоторых семьях уменьшены внешне; там, где они присутствуют, они не сфокусированы, в отличие от стемм личинок, которые полностью сфокусированы. Польза глазков в настоящее время не изучена.[5]
Бабочки и мотыльки умеют видеть ультрафиолетовый (УФ) свет, а также окраска и узор крыльев чешуекрылых в основном наблюдаются в этих длинах волн света.[13] Узоры, видимые на их крыльях в ультрафиолетовом свете, значительно отличаются от видимых при нормальном свете. Ультрафиолетовые лучи действуют как визуальные подсказки, которые помогают различать виды с целью спаривания. Были проведены исследования рисунков крыльев Lepidoptera (в основном бабочек), освещенных УФ-светом.[13]
Пальпи

Обычно губные щупики выступающие, трехчлениковые, выходящие из-под головы и загибающиеся вверх перед лицом.[7] Морфология лабиальных щупиков сильно различается у разных семейств Lepidoptera; иногда щупики отдельные, а иногда снисходительный и образуют клюв, но они всегда могут двигаться независимо. В других случаях лабиальные щупики могут быть не прямостоячими, а правильный (выдвигается вперед по горизонтали).[11][13] Щупики состоят из короткого базального сегмента, сравнительно длинного центрального сегмента и узкой конечной части. Первые два сегмента имеют плотный масштаб и могут быть волосатый; конечный сегмент голый. Конечный сегмент может быть тупым или заостренным; он может выступать прямо или под углом из второго сегмента, внутри которого он может быть скрыт.[11]
Ротовой аппарат


В то время как нижние челюсти или челюсти (жевательные части рта) присутствуют только на стадии гусеницы, ротовые органы большинства взрослых чешуекрылых в основном состоят из сосущих; эта часть называется хоботком или хаустеллум. Некоторые виды чешуекрылых имеют уменьшенный ротовой аппарат и не питаются во взрослом состоянии. Другие, такие как базальный семейство Micropterigidae, имеют жевательный ротовой аппарат.[21]
Хоботок (множественное число - хоботки ) формируется из верхнечелюстные галеи и приспособлен для сосания нектар.[3] Он состоит из двух трубок, соединенных крючками и отделяемых для очистки. Каждая трубка вогнута внутрь, образуя центральную трубку, через которую всасывается влага. Всасывание осуществляется за счет сжатия и расширения мешка в голове.[11] Хоботок свернут под головой, когда насекомое покоится, и расширяется только при кормлении.[16] Верхнечелюстные щупики уменьшены и даже рудиментарны. Они бросаются в глаза и имеют пять сегментов в некоторых из наиболее базальных семейств и часто складчатые.[7]
Форма и размеры хоботка эволюционировали, чтобы дать разным видам более широкий и, следовательно, более выгодный рацион.[3] Существует аллометрический масштабная взаимосвязь между массой тела Lepidoptera и длиной хоботка[22] от которого интересным адаптивным отклонением является необычно длинноязычная бабочка сфинкс. Xanthopan morganii praedicta. Чарльз Дарвин предсказал существование и длину хоботка этой бабочки до ее открытия, основываясь на своих знаниях о мадагасканской звездной орхидее с длинными шпорами. Angraecum sesquipedale.[23]
В первую очередь два кормление гильдий Lepidoptera - нектароядные, которые получают большую часть своих потребностей в питательных веществах из цветочного нектара, и те из гильдии плодоядных, которые питаются в основном соками гниющих фруктов или сбраживающим соком деревьев. Морфология хоботков обеих кормовых гильдий существенно различается. Ястребиные (семейство) Сфингиды ) имеют удлиненные хоботки, которые позволяют им питаться и опылять цветки с длинными трубчатыми венчиками. Помимо этого, ряд таксонов (особенно совок бабочки) развили различную морфологию хоботка. У некоторых видов совок развился пронзительный ротовой аппарат; на конце хоботка имеются склеротизированные чешуйки, которыми можно протыкать и высасывать кровь или фруктовые соки. Хоботки у некоторых Геликоний виды эволюционировали, чтобы потреблять твердые вещества, такие как пыльца.[24] Некоторые другие бабочки, в основном совки, модифицировали хоботки в соответствии со своим режимом питания - слезотечение (питаясь слезами спящих птиц). У хоботков часто есть острые вершины, а также множество зазубрин и шпор на стебле.[25][26]

Сканирующая электронная микрофотография хоботка моли из семейства Pyralidae

А нимфалида бабочка сосать банан

Сара длиннокрылая (Геликоний сара ), один из многих Геликоний виды, которые, как известно, питаются пыльцой, с пыльцой на хоботке

Xanthopan morganii, африканец сфингид, имеет хоботок длиной в фут, приспособленный для питания орхидей. Angraecum sesquipedale

Слезные Lepidoptera, такие как две бабочки Julia (Дриас Юлия ) пьют слезы черепах в Эквадоре, имеют крючки и зазубрины на кончике хоботка





Грудная клетка

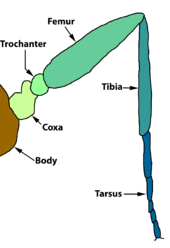
Грудь, которая развивается из 2, 3 и 4 сегментов личинки, состоит из трех незаметно разделенных сегментов, а именно: переднегрудь, заднегрудь, и среднегрудь.[11] Органы передвижения насекомых - ноги и крылья - несут на грудной клетке. Передние ноги отходят от переднегруди, передние крылья и средняя пара ног опираются на среднегрудь, а задние и задние конечности - от заднегруди. В некоторых случаях крылья рудиментарны.[11][27]
Верхняя и нижняя части грудной клетки (терга и стерна соответственно) состоят из сегментарных и внутрисегментарных склериты которые проявляют вторичную склеротизацию и значительные модификации у чешуекрылых. Переднегрудь - самый простой и маленький из трех сегментов, тогда как среднегрудь - наиболее развитая.[27]
Между головой и грудной клеткой находится перепончатая шея или шейка матки. Он состоит из пары боковых шейных склеритов и состоит как из головных, так и из грудных элементов.[10]:71[27] Между головой и грудной клеткой находится пучковая чешуя, называемая переднеспинка. По обе стороны от щита шкала, называемая лопатка.[11] в Noctuoidea, заднегрудь модифицируется парой барабанные органы.[7]
Нога
Передние ноги в Papilionoidea демонстрируют редукцию различных форм: семейство бабочек Нимфалиды У бабочек с кистоногими ногами, как их обычно называют, только две задние пары ног полностью функциональны, а передняя пара сильно сокращена и не способна ходить или садиться. У Lycaenidae предплюсна несегментирован, поскольку тарсомеры сливаются, и, когти предплюсны отсутствуют. Аролиарная подушечка (подушечка, выступающая между когтями предплюсны у некоторых насекомых) и пульвиллы (единственное число: pulvillus, подушечка или мочка под каждой лапкой предплюсны) уменьшены или отсутствуют в Papilionidae. Коготки предплюсны также отсутствуют у Riodinidae.[28]
У чешуекрылых все три пары ног покрыты чешуей.[13] У чешуекрылых также есть органы обоняния на ногах, которые помогают «пробовать» или «нюхать» пищевые растения.[6]
Крылья
Взрослые чешуекрылые имеют две пары перепончатых крыльев, обычно полностью покрытых мелкой чешуей. Крыло состоит из верхней и нижней мембран, которые соединены мельчайшими волокнами и усилены системой утолщенных полых ребер, часто, но неправильно называемых «венами», поскольку они также могут содержать трахеи, нервные волокна и кровеносные сосуды.[11][29] Перепонки покрыты мелкими чешуйками с неровными концами или волосками, прикрепленными крючками. Крылья двигаются за счет быстрого сокращения мышц и расширения грудной клетки.[11]
Крылья образуются из мезо- и заднегрудного сегментов и сходны по размеру в базальных группах. В более производных группах мезо-грудные крылья крупнее, с более мощной мускулатурой у их оснований и более жесткими структурами вен на нижних конечностях. реберный край.[7]
Помимо обеспечения основной функции полета, крылья также выполняют второстепенные функции: самооборона, камуфляж и терморегуляция.[3][30] В некоторых семьях чешуекрылых, таких как Psychidae и Lymantriidae крылья уменьшены или даже отсутствуют (часто у самок, но не у самцов).[7]
Форма
Форма крыльев чешуекрылых очень разнообразна. В случае Papilionoidea Коста может быть прямым или сильно изогнутым. Иногда бывает вогнутый на заднем крыле. Иногда это зазубрины или мелкие зубцы на переднем крыле. Вершина может быть закругленной, заостренной или серповидный (изготовлено и вогнутое внизу). В термен имеет тенденцию быть прямым или вогнутым на переднем крыле, тогда как оно обычно более или менее выпукло на заднем. Термен часто зубчатый или же зубчатый, т.е. производятся на каждой жилке и вогнуты между ними. Спинка обычно прямая, но может быть вогнутой.[11]
Заднее крыло часто хвостатый, т.е. жилки около конца торнус иметь один или несколько хвостов. Сам торнус часто образованный и часто лопастный.[11] Вдоль задних крыльев в двойной ряд плотно уложены чешуйки. Нижняя сторона чешуек выступает и образует регулярную узкую бахрому, называемую реснички.[11]

Шлейфовые мотыльки (семейство Pterophoridae ) имеют раздвоенные крылья

У многоперых мотыльков (семейство Alucitidae ) крылья разделены по каждой жилке

Microlepidoptera Гелехиоидея, Такие как Palumbina guerinii, имеют волосовидную бахрому на задних крыльях

Хвостатые задние крылья мадагаскарской моли (Chrysiridia rhipheus семья Uraniidae )

Lycaenids, такие как головоломка обезьяны (Ратинда любовь ) имеют нитевидные хвосты, которые пытаются объяснить гипотезой "ложной головы"[31]

Гиалиновый заплатки на крыльях колибри, ястреб-моль, снежно-ягодный ясенок (Hemaris diffinis )

Pachyerannis obliquaria, брачная пара - вверху крылатый самец, внизу маленькая бескрылая самка







Жилкование


Через двухслойное перепончатое крыло проходят трубчатые жилки. Вены связаны с гемоцель и теоретически позволить гемолимфе течь через них. Кроме того, по венам могут проходить нерв и трахея.[29]
Жилкование чешуекрылых простое, с небольшим количеством поперечин.[15]:88 Жилкование крыльев Lepidoptera является диагностическим средством для различения таксонов, а также родов и семейств.[13] Терминология основана на Система Комстока-Нидхема что дает морфологическое описание крыло насекомого жилкование.[32] У базальных чешуекрылых жилкование переднего крыла сходно с жилкованием заднего крыла; состояние, называемое «гомоневрным». В Micropterigidae (Zeugloptera) имеют жилкование, напоминающее самые примитивные ручейники (Trichoptera). Все остальные чешуекрылые, подавляющее большинство (около 98%), являются «гетероневральными», жилкование заднего крыла отличается от жилкования переднего и иногда сокращается. Бабочки из семей Nepticulidae, Opostegidae, Gracillariidae, Tischeriidae, и Bucculatricidae, среди прочего, часто имеют сильно уменьшенное жилкование на обоих крыльях.[13]:635[32]:56 Гомоневральные бабочки, как правило, имеют форму соединения крыльев «jugum», в отличие от расположения «уздечка – retinaculum» в случае более продвинутых семей.

Жилкование крыльев насекомых с названиями по системе Комстока – Нидхема.

Гомоневрозное жилкование в Sabatinca lucilia (Micropterigidae )

Гетероневрозное жилкование в Гонептерикс рамни (Pieridae )

Сниженное жилкование в Synanthedon tipuliformis (Sesiidae )




Крыло сцепления

Lepidoptera разработали широкий спектр морфологических механизмов соединения крыльев у имаго, которые делают эти таксоны «функционально двукрылыми» (двукрылыми).[33] Все формы, кроме самых базальных, демонстрируют это соединение крыльев.[34] Существует три различных типа механизмов - скуловой, уздечно-ретинакулярный и амплексиформный.[35]
Более примитивные группы имеют увеличенную лопастную область около базального заднего края (то есть у основания переднего крыла), называемую jugum, который во время полета складывается под заднее крыло.[7][34] У других групп есть уздечка на заднем крыле, которое цепляется за ретинакулум на переднем крыле.[7]
В целом бабочки (за исключением мужчин Euschemoninae ) И в Bombycoidea мотыльки (за исключением Сфингиды ) уздечка и удерживатель не соединяют крылья. Вместо этого увеличенная плечевая область заднего крыла широко перекрывается передним крылом. Несмотря на отсутствие определенной механической связи, крылья перекрываются и работают синфазно. Силовой ход переднего крыла в унисон толкает заднее крыло. Этот тип сцепления является разновидностью уздечки уздечки, но при которой уздечка и удерживатель полностью утрачены.[33][36]
Напольные весы

Крылья чешуекрылых ежеминутно масштабированный, который дает название этому заказу; название Чешуекрылые был придуман в 1735 году Карл Линней для группы «насекомых с четырьмя чешуйчатыми крыльями». Это получено из Древнегреческий лепис (λεπίς) означает «(рыба) чешуя» (и относится к лепеин «очистить») и птерон (πτερόν) означает «крыло».[37]
Чешуя также покрывает голову, части грудной клетки и живота, а также части гениталий. Морфология чешуек была изучена Дж. К. Дауни и А. К. Аллином (1975).[38] и чешуйки были разделены на три группы, а именно волосовидные или пилиформ, лезвие, или пластинчатый и другие вариативные формы.[8]
Примитивные бабочки (не Glossata и Eriocranidae) имеют «твердые» чешуйки без отверстий, т.е. просвет.[8]
Несколько таксонов Трихоптера (ручейники), которые являются сестринской группой чешуекрылых, имеют волосовидные чешуйки, но всегда на крыльях и никогда на теле или других частях насекомого.[13] У ручейников также есть хвостовые церки на брюшке, что отсутствует у чешуекрылых.[7] По словам Скобла (2005),[8] «морфологически чешуйки являются макротрихиями и, таким образом, гомологичны крупным волоскам (и чешуйкам), покрывающим крылья Trichoptera (ручейников)».
Структура
Несмотря на большое разнообразие форм масштабов, все они имеют схожую структуру. Весы, как и другие макрохеты, возникают из специальных трихогенных (продуцирующих волосы) клеток и имеют гнездо, заключенное в особую клетку «тормоген»;[15]:9 это расположение обеспечивает стебель или цветоножка при помощи которых чешуйки прикрепляются к субстрату. Чешуя может быть волосистой (волосовидной) или приплюснутой. Корпус или «лезвие» типичной плоской чешуи состоит из верхней и нижней ламелей с воздушным пространством между ними. Поверхность тела гладкая и известна как низший ламели. Верхняя поверхность, или начальство пластинка, имеет поперечные и продольные гребни и ребра. Ламели удерживаются распорками, называемыми трабакулы и содержат пигменты, придающие цвет. Чешуя немного свободно прилегает к крылу и легко отрывается, не причиняя вреда бабочке.[8][13][39]
Цвет
Чешуя на крыльях бабочки пигментирована меланины которые могут производить черный и коричневый цвета. Белый цвет в семействе бабочек Pieridae является производным от мочевая кислота, выделительный продукт.[13][40]:84 Яркие синие, зеленые, красные и переливчатость обычно создаются не пигментами, а микроструктурой чешуек. Этот структурная окраска это результат когерентное рассеяние света от фотонный кристалл характер чешуи.[41][42][43] Специализированные шкалы, которые обеспечивают структурную окраску отраженного света, в основном создают ультрафиолетовые узоры, которые можно различить в той части ультрафиолетового спектра, которую видят глаза чешуекрылых.[13] Видимый структурный цвет часто зависит от угла зрения. Например, в Морфо карповый, цвет спереди ярко-синий, но если смотреть под углом, он очень быстро становится черным.[44]
Радужная структурная окраска крыльев многих видов ликаенид и папилионид, таких как Parides sesostris и Тейнопальпус империалис и ликаениды, такие как Callophrys rubi, Cyanophrys remus, и Mitoura gryneus, был изучен.[45] Они проявляют самые сложные известные структуры фотонных масштабов - регулярные трехмерные периодические решетки, которые встречаются в просвете некоторых масштабов.[46] В случае с Kaiser-i-Hind (Тейнопальпус империалис ), трехмерная фотонная структура была исследована с помощью трансмиссионной электронной томографии и компьютерного моделирования, чтобы выявить встречающиеся в природе «хиральные тетраэдрические повторяющиеся единицы, упакованные в триклинную решетку»,[47][48] причина перелива.

Структурный синий цвет в морфо Cypris, нимфалида

Когда то же самое Морфо карповый Видно, что образец оканчивается, синий цвет становится черным.

Белый цвет у пиерид, таких как Делиас Эухарис является производным мочевой кислоты, продуктом выделения.

Зеленая переливчатость ласточкина хвоста Kaiser-i-Hind (Тейнопальпус империалис ) привел к открытию трехмерных фотонный кристалл структура.

Окраска крыльев некоторых чешуекрылых позволяет маскироваться, как это видно на примере геометрической бабочки. Колостигия водянистая.





Функция


Чешуя играет важную роль в естественной истории Lepidoptera. Весы позволяют развить яркие или нечеткие узоры которые помогают организму защитить себя камуфляж, мимикрия, и предупреждение. Помимо предоставления изоляция, темные узоры на крыльях позволяют поглощать солнечный свет и, вероятно, участвуют в терморегуляция. Помогают яркие и характерные цветовые узоры у бабочек, неприятные хищникам. общаться их токсичность или несъедобность, что предотвращает хищничество. В Бейтсовская мимикрия, окраска крыльев помогает съедобным чешуекрылым имитировать несъедобные модели, а в Мюллерова мимикрия несъедобные бабочки похожи друг на друга, чтобы сократить количество особей, отобранных неопытными хищниками.[8]
Первоначально весы могли развиться для обеспечения изоляции. Чешуя на грудной клетке и других частях тела может способствовать поддержанию высокой температуры тела, необходимой во время полета. Однако «твердые» чешуйки базальных бабочек не так эффективны, как чешуйки их более продвинутых родственников, поскольку наличие просвета добавляет воздушные слои и увеличивает изоляционные свойства.[8] Весы также помогают увеличить отношение подъемной силы к лобовому сопротивлению в полете.[8]
Для новоявленных взрослых большинства мирмекофил Lycaenidae листопадные восковые чешуи обеспечивают некоторую защиту от хищников, когда они выходят из гнезда.[8] В случае бабочки-мотылька (Liphyra brassolis ), гусеницы - нежеланные гости в гнездах древесных муравьев, питаясь их личинками. Взрослые особи, выходящие из куколок, покрыты мягкими рыхлыми липкими чешуйками, которые стираются и прилипают к муравьям, когда они выходят из гнезда после вылупления.[49]

Андрокония
Самцы чешуекрылых обладают особой чешуей, называемой андрокония (единственное число - андроконий), которые возникли в результате половой отбор в целях распространения феромоны для привлечения подходящих товарищей. Андроконии могут быть распределены на крыльях, теле или ногах или встречаться в виде пятен, называемых «клеймами», «половыми марками» или «стигматами» на крыльях, обычно в инвагинации верхней поверхности передних крыльев, иногда скрытой другими чешуйками. Андроконии также встречаются в складках крыльев. Эти марки иногда состоят из пучков волос, которые способствуют распространению феромона. Роль андроконии в ухаживании за бабочками-пирид и нимфалидами, такими как Pyronia tithonus и Дриас Юлия, доказано экспериментально.[15]:16–17[50][51][52][53]
Последовательные крупные планы чешуи павлиньего крыла
Фотографические и световые микроскопические изображения Уменьшенный вид Аглаис ио. Крупный план чешуи того же экземпляра. Большое увеличение цветных чешуек (вероятно, другого вида). Электронно-микроскопические изображения Кусок крыла Весы крупным планом Единая шкала Микроструктура чешуи Увеличение Прибл. × 50 Прибл. × 200 ×1000 ×5000







Брюшная полость
В брюшная полость или тело состоит из девяти сегментов. У личинки он колеблется от 5 до 13. Одиннадцатый сегмент личинки держит пару анальных кламмеров, которые в некоторых местах выступают вперед. таксоны и представляют гениталии.[11]
У многих семейств моли есть специальные органы, помогающие обнаруживать эхолокация летучих мышей. Эти органы известны как тимпана (единственное число - типанум). В Pyraloidea и почти все Геометроидеа имеют барабанную перепонку, расположенную на передней стернит живота. В Noctuoidea также есть тимпана, но в их случае тимпана расположена на нижней стороне заднегрудь, структура и положение которых являются уникальными и таксономически отличительными признаками надсемейства.[21][54]
У самок некоторых бабочек есть орган, излучающий запах, расположенный на кончике брюшка.[6]
Гениталии

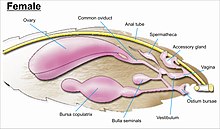
В гениталии являются сложными и служат основой для видовой дискриминации в большинстве семей, а также для идентификации семей.[7] Гениталии возникают из десятого или самого дистального сегмента брюшной полости. Чешуекрылые имеют одни из самых сложных половых структур среди всех насекомых, с большим разнообразием сложных шипов, щетинок, чешуек и пучков у самцов, кламмеров различной формы и модификаций ductus bursae у женщин, через которые сохраненная сперма передается внутри женщины прямо или косвенно во влагалище для оплодотворения.[13][55][56]
Расположение гениталий важно при ухаживании и спаривании, поскольку они предотвращают перекрестное спаривание и гибридизацию. Уникальность гениталий вида привела к использованию морфологического исследования гениталий в качестве одного из наиболее важных ключей в таксономической идентификации таксонов ниже уровня семейства. С появлением анализа ДНК изучение гениталий стало лишь одним из методов, используемых в систематике.[6]
У большинства чешуекрылых существует три основных конфигурации гениталий, основанные на том, как эволюционировало расположение у самок отверстий для копуляции, оплодотворения и откладки яиц:
- Экзопорианец: Hepialidae и родственные семьи имеют внешнюю бороздку, по которой сперма переносится от копулятивного отверстия (гонопора) к (яйцекладу), и называются экзопорианскими.[7]
- Монотризский: У примитивных групп есть одно генитальное отверстие в конце брюшка, через которое происходит совокупление и откладка яиц. Этот символ используется для обозначения Monotrysia.[7]
- Дитрисян: Остальные группы имеют внутренний проток, по которому идет сперма и образующий Ditrysia, с отдельными отверстиями для копуляции и откладки яиц.[7]
Гениталии самца и самки у любого конкретного вида приспособлены друг к другу, как замок (самец) и ключ (самка).[6] У мужчин девятый сегмент брюшка делится на спинной "тегумен" и вентральный «Виникулум».[56] Они образуют кольцевидную структуру для прикрепления половых органов и пары латеральных обхватывающих органов (кламмеров или «арп»). У самца есть средний трубчатый орган (называемый эдеагус ), который проходит через разворачивающуюся оболочку (или «vesica») для оплодотворения самки.[55] У самцов у всех чешуекрылых парные семявыносящие протоки; парные семенники обособлены у базальных таксонов и слиты у продвинутых форм.[55]
В то время как расположение внутренних половых протоков и отверстий гениталий самок зависит от таксономической группы, к которой принадлежит насекомое, внутренняя репродуктивная система самок всех чешуекрылых состоит из парных яичников и дополнительных желез, которые производят желтки и скорлупу яиц. У самок насекомых есть система сосудов и протоков, в которых сперма поступает, транспортируется и хранится. Яйцеводы женщины соединяются вместе, образуя общий проток (называемый «oviductus communis»), который ведет к влагалищу.[55][56]
Когда происходит совокупление, самец бабочки или мотылька кладет капсулу спермы (сперматофор ) в сосуде самки (называемом бурса тела). Сперма при выходе из капсулы попадает прямо в или через небольшую трубку в специальное семяприемник (сперматека ), где сперма хранится до тех пор, пока она не попадет во влагалище для оплодотворения во время кладки яиц, что может произойти через часы, дни или месяцы после спаривания. Яйца проходят через яйцеклад. Яйцеклад может быть на конце модифицированного яйцеклада или окружен парой широких щетинистых анальных сосочков.[55][56]
Бабочки Парнасины (семейство Papilionidae) и некоторые Acraeini (семейство Nymphalidae) добавляют посткопуляционную пробку, называемую сфрагис к брюшку самки после совокупления, не позволяя ей снова спариваться.[13]
Самцы многих видов Papilionoidea обладают вторичными половыми признаками. Они состоят из органов, излучающих запахи, кистей, а также марок или мешочков со специальной чешуей. По-видимому, они выполняют функцию убеждения самки в том, что она спаривается с самцом правильного вида.[11]
Было зарегистрировано, что три вида боярышника издают щелчки ультразвуком, потирая свои гениталии; самцы производят, натирая твердые чешуйки на внешней стороне застежки в то время как самки издают звук, сокращая свои гениталии, что вызывает трение чешуей о живот. Функция создания этого шума не ясна, и выдвигаемые предложения включают подавление эхолокации летучих мышей и рекламу того, что добыча летучей мыши - колючие и отличные летчики.[57]

Citheronia regalis с застежками закрытыми

Citheronia regalis с открытыми застежками

Крупным планом - затвердевший сфрагис, выступающий на 2-3 мм позади брюшной полости. Парнас




Cloaca
У чешуекрылых насекомых есть клоака в конце живота. Он может быть полным, включая анус, то яйцеклад и копулятивная пора, как и в случае Дакнонифа, Zeugloptera и большинство Монотризия; или неполный, включающий только задний проход и яйцеклад, как это встречается у некоторых Monotrysia, Psychidae, а в некоторых Choreutidae и Cossidae.[58]
Разработка

Оплодотворенная яйцеклетка созревает и из нее вылупляется гусеница. Гусеница - это питательная стадия жизненного цикла чешуекрылых. Гусеница должна иметь возможность питаться и избегать того, чтобы ее поедали, и большая часть ее морфологии эволюционировала, чтобы облегчить эти две функции.[59]:108 После роста и шелушение гусеница входит в сидячий стадия развития, называемая куколкой (или куколкой), вокруг которой она может образовывать оболочку. Насекомое развивается во взрослую особь на стадии куколки; когда куколка готова, вылупляется и взрослая стадия или имаго бабочки или мотылька.
Яйцо

Как и большинство насекомых, чешуекрылые яйцекладущий или «яичные несушки».[40] Яйца чешуекрылых, как и яйца других насекомых, центролецитальный в том, что яйца имеют центральную желток окружен цитоплазма. Желток обеспечивает жидкое питание для эмбрион гусеница, пока она не вырвется из панциря.[60] Цитоплазма окружена вителиновый конверт и белковая мембрана, называемая хорион защищает яйцо внешне.[60][61] Ядро зиготы расположено сзади.[61]
У некоторых видов чешуекрылых внутри хориона, прилегающего к желточному слою, присутствует восковой слой, который, как считается, образовался для предотвращения высыхания. У насекомых хорион имеет слой воздушных пор в твердом материале, который обеспечивает очень ограниченную способность к дыхательной функции. У чешуекрылых слой хориона над этим слоем воздушных пор является ламеллярным с последовательными слоями белка, расположенными в определенном направлении и ступенчатыми так, чтобы образовывать спиральное расположение.[61]
Верхняя часть яйца вдавлена и образует небольшую центральную полость, называемую микропиле через которую оплодотворяется яйцеклетка.[11] В яйцах шаровидной, конической или цилиндрической формы микропиле располагается сверху; у тех яиц, которые имеют уплощенную или двояковыпуклую форму, микропиле располагается на внешнем крае или краю.[18][62]
Яйца чешуекрылых обычно округлые и маленькие (1 мм), хотя они могут достигать 4 мм в случае Сфингиды и Saturniidae.[13]:640 Обычно они довольно простые по цвету, белые, бледно-зеленые, голубовато-зеленые или коричневые. Яйца бабочек и моли бывают разных форм; одни сферические, другие полусферические, конические, цилиндрические или линзовидные (линзовидные). Некоторые из них имеют форму бочонка или блина, а другие - тюрбана или сыра. Они могут быть наклонными или вдавленными на обоих концах, ребристыми или орнаментированными, пятнистыми или дефектными.[18][62]
Яйца откладываются поодиночке, небольшими группами или массой, и неизменно на источнике пищи или рядом с ним. Известно, что моли в неволе откладывают яйца в клетки, в которых они были изолированы.[18][62] На размер яиц Lepidoptera влияет ряд факторов. Lepidoptera виды, которые перезимовать на стадии яйца обычно есть более крупные яйца, чем у видов, у которых их нет. Точно так же виды, питающиеся древесными растениями на личиночной стадии, имеют более крупные яйца, чем виды, питающиеся травянистыми растениями. Яйца, откладываемые старшими самками некоторых видов бабочек, были меньше по размеру, чем их более молодые собратья. При отсутствии полноценного питания самки кукурузной мотылька ( Остриния spp.) были зарегистрированы для откладывания кладок с размером яиц ниже нормального.[61]
Во время побега только что вылупившиеся личинки многих видов иногда поедают хорион, чтобы вылупиться. В качестве альтернативы, скорлупа яйца может иметь слабую линию вокруг шляпки, которая уступает место, позволяя личинке выйти.[61] Яичная скорлупа и небольшое количество желтка, попавшие в амниотические оболочки, составляют первую пищу для большинства личинок чешуекрылых.

Яйца пионерские (Anaphaeis aurota семья Pieridae)

Яйца малиновой розы (Атрофаневра Гектор семейство Papilionidae)

Яйцо мальвы шкиперское (Carcharodus alceae семейство Hesperiidae)

Яйцо крупное медное (Lycaena dispar семейство Lycaenidae)





Бок-о-бок яйца двустворчатого чешуекрылого, кипарисовика листороса (Archips goyerena семья Tortricidae )

Прямостоячие яйца дитризового чешуекрылого, лунного мотылька (Actias luna семейство Saturniidae) в неволе на бумаге

Яйца сосновой моли (Bupalus piniaria семейство Geometridae)

Яйца моли лакейной (Малакосома невстрия семейство Lasiocampidae)




Гусеница
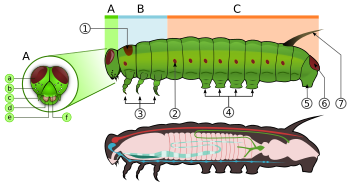
Гусеницы, "характерные" полипод личинки с цилиндрическими телами, короткими грудными ногами и брюшными ложноножками (ложноножками) ».[63] У них закаленные (склеротизированный ) головная капсула, нижние челюсти (ротовые части) для жевания и мягкое трубчатое сегментированное тело, которое может иметь волосовидные или другие выступы, три пары настоящих ног и дополнительные бездельники (до пяти пар).[2] Тело состоит из тринадцати сегментов, три из которых грудные (Т1, Т2 и Т3) и десять брюшных (от A1 до A10).[21]




У всех настоящих гусениц есть перевернутая Y-образная линия, идущая от макушки вниз. Между Y-образной линией лежит фронтальный треугольник или лоб. В наличник, расположенный ниже лба, лежит между двумя усики. В верхняя губа находится ниже наличника. В центре верхней губы есть небольшая выемка, с которой зацепляется край листа, когда гусеница ест.[64]
Личинки имеют шелк железы, которые расположены на губа. Эти железы модифицированы слюнные железы. Они используют эти шелковые железы для изготовления шелка для коконов и убежищ.[21] Под верхней губой расположены нижние челюсти.[64] На каждой стороне головы обычно по шесть стволовые чуть выше нижних челюстей. Эти стемматы расположены полукругом. Под стволом есть небольшая пара антенн, по одной с каждой стороны.[21][64]
Грудь несет три пары ног, по одной паре на каждом сегменте. Переднегрудь (T1) имеет функциональное дыхальце, которое фактически происходит от среднегруди (T2), в то время как заднегрудь имеет уменьшенное дыхальце, которое не открыто снаружи и находится под кутикулой.[59]:114 Грудные ноги состоят из тазика, вертела, бедренной кости, предплюсны и когтя и имеют неизменную форму во всем отряде. Однако у некоторых минеров они уменьшены, а у некоторых - удлинены. Notodontidae. У Micropterigidae ноги трехчлениковые, тазик, вертел и бедро слиты.[59]:114
Третий, шестой и десятый сегменты брюшка могут иметь по паре более мясистых ног.[21] Грудные ноги известны как настоящие ноги, а брюшные ноги - ложные.[64] Настоящие ноги мало различаются у чешуекрылых, за исключением уменьшения у некоторых листовых минеров и удлинения в семействе. Notodontidae.[59]:114 Ножки содержат несколько маленьких крючков на конце, которые известны как крючки. Семейства чешуекрылых различаются по количеству и расположению ложноножек. Некоторые личинки, например, глисты (Geometridae ) и петлители (Plusiinae ) имеют пять пар ложноножек или меньше, а другие любят Lycaenidae и гусеницы слизняков (Limacodidae ) вообще отсутствуют ложные ножки.[21][64] У некоторых гусениц, добывающих листья, на брюшной стенке присутствуют вязаные крючки, которые представляют собой уменьшенные ложные ножки, в то время как у других видов, добывающих листья, петли полностью отсутствуют.[65] Брюшные дыхальца расположены с каждой стороны тела на первых восьми брюшных сегментах.[64]
Гусеницы имеют разные виды выступов; щетинки (волоски), шипы, бородавки, бугорки и рога. Волосы бывают разных цветов и могут быть длинными или короткими; одиночные, скоплениями или пучками; тоньше на острие или с булавой на конце. Позвоночник может быть Chalaza (имеющий одну точку) или скол (имеющий несколько точек). Бородавки могут быть небольшими шишками или короткими выступами на теле. Бугорки - это мясистые выступы тела, короткие и бугристые или длинные и похожие на нити. Обычно они встречаются парами или кластером на одном или нескольких сегментах. Рога короткие, мясистые, остроконечные. Обычно они находятся на восьмом брюшном сегменте.[65]
Большое количество видов семейств Saturniidae, Limacodidae, и Megalopygidae есть жалящие гусеницы с ядовитыми щетинками, называемыми мочеиспускание, а в случае Лономия - а Бразильский род сатурниид - может убить человека из-за своей мощи антикоагулянт яд.[13]:644 Гусеницы многих таксонов, которые секвестировали токсичные химические вещества из растений-хозяев или имеют острые мочеиспускательные волосы или шипы, проявляют апосематический окраска и маркировка.[66]
Гусеницы подвергаются шелушение и имеют ряд личинок возрастов, обычно пять, но различаются между видами. Новая кутикула мягкая и позволяет гусенице увеличиваться в размерах и развиваться, прежде чем она станет твердой и неэластичной. При последнем шелушении старая кутикула расщепляется и сворачивается в небольшой клубок на заднем конце куколки и называется личинкой. экзувия.[67]:31

Два возрастов папилионида простой мормон с разными схемами камуфляжа - напоминающими птичий помет и растительность

Личинки нотодонтидных молей, например, Stauropus fagi, имеют удлиненные грудные ноги.

Личинка Lonomia obliqua, а сатурниид мотылек из Бразилии, имеет мочеиспускание со смертельным антикоагулянт яд.

Седловидный мотылек (Ахария стимула ) личинки имеют апосематическую окраску в виде седла.

Нижняя сторона слизняковых гусениц Phobetron pithecium (семейство Limacodiae) с отсутствием ложноножек

Гусеница минера осина обыкновенная (Phyllocnistis populiella )

Стрельба из красного дерева (Hypsipyla grandella ) убытки красное дерево в Бразилии.

Гусеница мешочницы (возможно Hyalarcta huebneri семья Psychidae ) выходит из корпуса








Хризалис или куколка



Кокон - это оболочка, сплетенная из шелк многими гусеницами моли и многими другими голометаболический насекомое личинки как защитное покрытие для куколки. Большинство личинок Lepidoptera либо кокон и окукливаться внутри них или окуклится в клетке под землей,[21] за исключением бабочек и современных моли, таких как совки, куколки которого обнажены.[13] Куколки бабочек обычно коричневые и гладкие, тогда как куколки бабочек часто красочные и их форма сильно различается.[21] У бабочек обнаженную куколку часто называют куколка, полученный из Греческий термин «куколка»: χρυσός (Chrysós) за золото, имея в виду золотистый цвет некоторых куколок.[68]
Гусеницы многих бабочек прикрепляются шелковой пуговицей к нижней стороне ветки, камня или другой выступающей поверхности. Они остаются прикрепленными к шелковой подушечке с помощью крючкового процесса, называемого кремастером. Большинство куколок свешиваются вниз головой, но в семействах Papilionidae, Pieridae и Lycaenidae куколка удерживается в более вертикальном положении за счет шелкового пояса вокруг середины куколки.[21]
Куколки большинства Lepidoptera обнаруживать, с придатками, сращенными или приклеенными к телу, а остальные имеют исключать куколки, у которых усики, ноги и крылья свободны и не приклеены к телу.[69]
На стадии куколки морфология имаго развивается за счет развития личиночных структур.[40]:151 Общий вид взрослого человека виден до того, как затвердеет внешняя поверхность - голова, опирающаяся на грудную клетку, глаза, усики (вынесенные вперед над головой), крылья, расположенные над грудной клеткой, и шесть ног между крыльями и грудной клеткой. брюшная полость.[70] Среди особенностей, заметных в области головы куколки, можно выделить склериты, швы, пилиферы, нижние челюсти, окуляры, усики, щупики и верхние челюсти. Куколочная грудная клетка состоит из трех грудных сегментов, ног, крыльев, тегул, крыловидных борозд и подмышечных бугорков. На брюшке куколки десять сегментов, шипы, щетинки, рубцы личиночных ложноножек и бугорков, анальное и генитальное отверстия, а также дыхальца.Куколки мотыльков имеют фланцевые пластины, а у специализированных чешуекрылых - кремовую пластину.[14]:23–29
В то время как куколка обычно неподвижна и неподвижна, куколки примитивных семейств моли Micropterigidae, Agathiphagidae, и Heterobathmiidae имеют полностью функциональные нижние челюсти.[59]:131 Они служат главным образом для того, чтобы взрослые особи могли вырваться из кокона.[14]:34 Кроме того, все придатки и тело отделены от кожи куколки и обладают некоторой степенью независимого движения. Все другие надсемейства чешуекрылых более специализированы, имеют нефункциональные нижние челюсти, придатки и тело, прикрепленные к коже куколки, и теряют некоторую степень независимого движения.[14]:20
Куколки некоторых бабочек умеют шевелить брюшком. Три хвостовых сегмента куколочного брюшка (8-10 сегменты) фиксированы; остальные сегменты в некоторой степени подвижны. В то время как более развитые чешуекрылые могут изгибать только последние два или три сегмента на конце брюшка, большее количество базальных таксонов, таких как Micropterigidae может пошевелить оставшимися семью сегментами живота; это, по-видимому, помогает им выступать передним концом куколки до того, как восторг.[14]:28[67] Куколки Hepialidae могут двигаться вперед и назад в личиночном туннеле, извиваясь, чему способствуют не только шипы, но и выступы на спине.[67] Считается, что извивающиеся брюшные мышцы могут напугать хищников и отпугнуть их. В случае нескольких ястребиная моль, Такие как Theretra latreillii шевеление живота сопровождается дребезжащим или щелкающим звуком, который усиливает эффект вздрагивания.[67]
У некоторых видов, например Геликоний харитония, спаривание может происходить внутри куколки самок самцами.[71]

Папилионидные хризалиды обычно прикрепляются к субстрату кремастером и держат голову вверх с помощью шелк пояс.

Подвешенная куколка нимфалида золотистого цвета Ядро евплои

Actias luna (семья Saturniidae ) выходящий из кокона

Специализированная куколка сфингидной бабочки, Agrius convolvuli, может шевелить животом, издавая щелкающий звук, который может испугать.



Защита и хищничество



Чешуекрылые обладают мягким телом, хрупкими и почти беззащитными, в то время как незрелые стадии движутся медленно или неподвижны, поэтому все стадии подвергаются воздействию хищничество птицами, мелкими млекопитающими, ящерицами, амфибиями, беспозвоночными хищниками (особенно паразитоид и паразитический осы и мухи ), а также грибки и бактерии. Для борьбы с этим Lepidoptera разработали ряд стратегий защиты и защиты, которые включают: камуфляж, апосематизм, мимикрия, а также разработка моделей угроз и их отображения.[72]
Камуфляж - важная стратегия защиты, которая обеспечивается изменениями формы, цвета и отметин тела. Некоторые чешуекрылые сливаются с окружающей средой, поэтому хищники их не заметят. Гусеницы могут иметь оттенки зеленого, соответствующие цвету растения-хозяина. Другие напоминают несъедобные предметы, например веточки или листья. Личинки некоторых видов, например, простой мормон и Западный тигровый ласточкин хвост похожи на птичий помет.[72][73]
Некоторые виды Lepidoptera изолируют или производят токсины, которые хранятся в тканях их тела, что делает их ядовитыми для хищников; примеры включают бабочка монарх в Америке и Атрофаневра вид в Азии. Хищники, которые едят ядовитых чешуекрылых, могут заболеть и сильно рвать, и поэтому научатся избегать этих видов. Хищник, который ранее ел ядовитого чешуекрылого, может в будущем избегать других видов с аналогичными отметинами, тем самым спасая и многие другие виды.[72][74] Токсичные бабочки и личинки, как правило, приобретают яркую окраску и яркие узоры, что является индикатором их токсичности для хищников. Это явление известно как апосематизм.[75]
Апосематизм привел также к развитию комплексов мимикрии Бейтсовская мимикрия, где съедобные виды имитируют апосематические таксоны, и Мюллерова мимикрия, где несъедобные виды, часто относящиеся к родственным таксонам, эволюционировали, чтобы походить друг на друга, чтобы получить выгоду от сокращения количества выборок хищниками во время обучения. Точно так же взрослый Sesiidae виды (также известные как яснокрылые мотыльки) имеют общий вид, достаточно похожий на Оса или же шершень чтобы уменьшить вероятность того, что бабочки станут хищниками за счет мимикрии Бейтса.[76]
Пятна для глаз являются разновидностью автомимикрия используется некоторыми чешуекрылыми. У бабочек пятна состоят из концентрических колец чешуек разного цвета. Предполагаемая роль глазных пятен - отвлекать внимание хищников. Их сходство с глазами вызывает у хищника инстинкт атаковать эти узоры крыльев.[77] Предполагается, что роль нитевидных хвостов у Lycaenidae сбивает хищников с толку относительно реального расположения головы, что дает им больше шансов спастись живыми и относительно невредимыми.[78]
Некоторые гусеницы, особенно представители Papilionidae, содержат осметерий, Y-образный выступ железа найдено в переднегрудной сегмент личинки. При угрозе гусеница издает неприятный запах из органа, чтобы отпугнуть хищников.[79][80]
Смотрите также
- Различия между бабочками и молью
- Словарь терминов энтомологии
- Морфология насекомых
- Чешуекрылые
- Морфология (биология)
Сноски
- ^ Kristensen, Niels P .; Скобл, М. Дж .; Карсхольт, Оле (2007). Z.-Q. Чжан; W. A. Shear (ред.). Трехсотлетие Линнея: прогресс в таксономии беспозвоночных (PDF). Zootaxa. 1668. С. 699–747. Дои:10.11646 / zootaxa.1668.1.30. ISBN 978-0-12-690647-9.
Глава: «Филогения и систематика чешуекрылых: состояние инвентаризации разнообразия моли и бабочек»
- ^ а б Дагдейл, Дж. С. (1996). "Естественная история и идентификация питающихся подстилкой личинок чешуекрылых (Insecta) в буковых лесах, долина Оронго-Ронго, Новая Зеландия, с особым упором на рацион мышей (Mus musculus)" (PDF). Журнал Королевского общества Новой Зеландии. 26 (4): 251–274. Дои:10.1080/03014223.1996.9517513.[постоянная мертвая ссылка ]
- ^ а б c d Скобл, М. Дж. (1995). "Ротовой аппарат". Чешуекрылые: форма, функции и разнообразие. Oxford University Press. С. 6–19. ISBN 978-0-19-854952-9.
- ^ Боррор, Дональд Дж .; Триплхорн, Чарльз А .; Джонсон, Норман Ф. (1989). Введение в изучение насекомых (6, иллюстрированное изд.). Публикации колледжа Сондерс. ISBN 978-0-03-025397-3. Получено 16 ноября 2010. (Нет предварительного просмотра.)
- ^ а б c Скобл (1995). Раздел «Сенсация», (стр. 26–38).
- ^ а б c d е ж грамм час Хоскинс, Адриан. "Анатомическая голова бабочки (и другие страницы)". Узнайте о бабочках. Получено 15 ноября 2010.
- ^ а б c d е ж грамм час я j k л м п Пауэлл, Джерри А. (2009). «Чешуекрылые». In Resh, Vincent H .; Cardé, Ring T. (ред.). Энциклопедия насекомых (2-е изд.). Академическая пресса. С. 661–663. ISBN 978-0-12-374144-8.
- ^ а б c d е ж грамм час я Скобл (1995). Раздел «Весы», (стр. 63–66).
- ^ Маллет, Джим (12 июня 2007 г.). «Подробная информация о проектах таксома чешуекрылых и бабочек». Проект таксома Lepidoptera. Университетский колледж Лондона. Получено 14 ноября 2010.
- ^ а б c Жилло, Седрик (1995). «Бабочки и мотыльки». Энтомология (2-е изд.). ISBN 978-0-306-44967-3.
- ^ а б c d е ж грамм час я j k л м п о п q р s т Эванс, В. Х. (1932). "Вступление". Идентификация индийских бабочек (2-е изд.). Мумбаи: Бомбейское общество естественной истории. С. 1–35.
- ^ «Чешуекрылые». Энциклопедия Britannica Online. 2011. Получено 12 февраля 2011.
- ^ а б c d е ж грамм час я j k л м п о п q р s Хеппнер, Дж. Б. (2008). «Бабочки и мотыльки». В Capinera, Джон Л. (ред.). Энциклопедия энтомологии. Виртуальная справочная библиотека Gale. 4 (2-е изд.). Ссылка Springer. п. 4345. ISBN 978-1-4020-6242-1.
- ^ а б c d е ж Мошер, Эдна (2009) [1918]. Классификация чешуекрылых по признакам куколки (переиздание ред.). БиблиоБазар, ООО. ISBN 978-1-110-02244-1.
- ^ а б c d е Кристенсен, Нильс П. (2003). Чешуекрылые, мотыльки и бабочки: морфология, физиология и развитие, Том 2. Том 4, часть 36 Handbuch der Zoologie. Вальтер де Грюйтер. ISBN 978-3-11-016210-3.
- ^ а б c d Скобл (1995). Раздел «Голова взрослого - питание и ощущение» (стр. 4–22).
- ^ Хеппнер, Джон Б. (2008). «Мотылек шелкопряда (Lepidoptera: Bombycidae)». В Capinera, Джон Л. (ред.). Энциклопедия энтомологии. Springer Нидерланды. С. 3375–3376. Дои:10.1007/978-1-4020-6359-6_4198. ISBN 9781402062421.
- ^ а б c d Голландия, У. Дж. (1903). "Вступление" (PDF ). Книга Мотыльков. Лондон: Хатчинсон и Ко. ISBN 978-0-665-75744-0.
- ^ Мерлин, Кристина; Gegear, Роберт Дж .; Репперт, Стивен М. (2009). «Антенные циркадные часы координируют ориентацию солнечного компаса у мигрирующих бабочек-монархов». Наука. 325 (5948): 1700–1704. Bibcode:2009Научный ... 325.1700M. Дои:10.1126 / science.1176221. ЧВК 2754321. PMID 19779201.
- ^ Робинсон, Г.С. (1988). «Филогения Tineoidea (Lepidoptera)». Систематика и эволюция насекомых. Брилл. 19 (2): 117–129. Дои:10.1163 / 187631289x00113.. «... у многих Amphitheridae (s.l.) сложный глаз мужчины частично или полностью разделен по горизонтали».
- ^ а б c d е ж грамм час я j k Триплхорн, Чарльз А .; Джонсон, Норман Ф. (2005). Введение Боррора и Делонга в изучение насекомых. Белмонт, Калифорния: Томсон Брукс / Коул. ISBN 978-0-03-096835-8.
- ^ Agosta, Salvatore J .; Янзен, Даниэль Х. (2004). «Распределение размеров тела крупных костариканских бабочек сухих лесов и лежащая в основе взаимосвязь между морфологией растений и опылителей». Ойкос. 108 (1): 183–193. Дои:10.1111 / j.0030-1299.2005.13504.x.
- ^ Кунте, Крушнамег (2007). «Аллометрия и функциональные ограничения на длину хоботка у бабочек». Функциональная экология. 21 (5): 982–987. Дои:10.1111 / j.1365-2435.2007.01299.x.
- ^ Krenn, H.W .; Пенц, К. М. (1 октября 1998 г.). "Ротовые органы Геликоний бабочки (Lepidoptera: Nymphalidae): поиск анатомических приспособлений к питанию пыльцой ". Международный журнал морфологии и эмбриологии насекомых. 27 (4): 301–309. Дои:10.1016 / S0020-7322 (98) 00022-1.
- ^ Маккензи, Дебора (20 декабря 2006 г.). «Мотыльки пьют слезы спящих птиц». Новый ученый. Reed Business Information. Получено 10 февраля 2012.
- ^ Хилгартнер, Роланд; Раойлисон, Мамизоло; Бюттикер, Вильгельм; Лиз, Дэвид С.; Кренн, Харальд В. (22 апреля 2007 г.). «Малагасийские птицы как хозяева для глазных бабочек». Письма о биологии. 3 (2): 117–120. Дои:10.1098 / rsbl.2006.0581. ЧВК 2375961. PMID 17251126.
- ^ а б c Scoble (1995) Глава 3: «Грудная клетка взрослого - исследование функции и эффекта» (стр. 39–40).
- ^ Скобл, М. Дж .; Айелло, Аннетт (1990). «Бабочки, похожие на мотыльков (Hedylidae: Lepidoptera): резюме, с комментариями к яйцу» (PDF). Журнал естественной истории. 24 (1): 159–164. Дои:10.1080/00222939000770101.[постоянная мертвая ссылка ]
- ^ а б Чепмен, Р. Ф. (1998). «Торакс». Насекомые: строение и функции (4-е изд.). Издательство Кембриджского университета. п.45. ISBN 978-0-521-57890-5..
- ^ Кришна, Анирудх; Не, Сяо; Уоррен, Эндрю Д.; Льоренте-Бускетс, Хорхе Э .; Briscoe, Adriana D .; Ли, Джэхо (2020). «Инфракрасные оптические и тепловые свойства микроструктур крыльев бабочки». Труды Национальной академии наук. 117 (3): 1566–1572. Дои:10.1073 / pnas.1906356117. ISSN 0027-8424. ЧВК 6983360. PMID 31919285.
- ^ Роббинс, Роберт К (1981). «Гипотеза« ложной головы »: хищничество и изменение формы крыльев бабочек Lycaenid». Американский натуралист. 118 (5): 770–775. Дои:10.1086/283868.
- ^ а б Скобл (1995). Раздел «Крылья». Стр. 55.
- ^ а б Дадли, Роберт (2002). Биомеханика полета насекомых: форма, функции, эволюция. Princeton University Press. ISBN 978-0-691-09491-5.
- ^ а б Акции, Ян (2008). «Крыло сцепления». В Capinera, Джон Л. (ред.). Энциклопедия энтомологии. Виртуальная справочная библиотека Gale. 4 (2-е изд.). Ссылка Springer. п. 4266. ISBN 978-1-4020-6242-1.
- ^ Скобл (1995). Раздел «Сцепление крыла», (стр. 56–60).
- ^ Горб, Станислав (2001). «Блокировка частей тела». Приспособления для прикрепления кутикулы насекомых. Springer. п. 305. ISBN 978-0-7923-7153-3.
- ^ Харпер, Дуглас. «Чешуекрылые». Интернет-словарь этимологии. Получено 21 ноября 2010. из "Чешуекрылые "на сайте Dictionary.com.
- ^ Downey, J.C .; Аллин, A.C. (1975). «Морфология и номенклатура крыла». Бык. Аллин Мус. 31: 1–32.
- ^ Чепмен (1988). Раздел «Крылья и полет» (с. 190).
- ^ а б c Gullan, P.J .; Крэнстон, П. С. (2005). Насекомые: очерк энтомологии (3-е изд.). Wiley-Blackwell. ISBN 978-1-4051-1113-3.
- ^ Мейсон, К. У. (1927). «Структурные цвета у насекомых - II». Журнал физической химии. 31 (3): 321–354. Дои:10.1021 / j150273a001.
- ^ Вукусич, П. (2006). «Структурная окраска чешуекрылых» (PDF). Текущая биология. 16 (16): R621 – R623. Дои:10.1016 / j.cub.2006.07.040. PMID 16920604. S2CID 52828850.
- ^ Prum, R.O .; Quinn, T .; Торрес, Р. Х. (2006). «Анатомически разнообразные чешуйки бабочек дают структурные цвета за счет когерентного рассеяния». Журнал экспериментальной биологии. 209 (4): 748–765. Дои:10.1242 / jeb.02051. PMID 16449568.
- ^ Киношита, Шу-ичи (2008). Структурные цвета в царстве природы. World Scientific. стр.52 –53. ISBN 978-981-270-783-3.
- ^ Michielsen, K .; Ставенга, Д. Г. (2008). «Гироидные кутикулярные структуры в чешуях крыльев бабочки: биологические фотонные кристаллы». Журнал интерфейса Королевского общества. 5 (18): 85–94. Дои:10.1098 / rsif.2007.1065. ЧВК 2709202. PMID 17567555.
- ^ Поладиан, Леон; Уикхэм, Шелли; Ли, Кван; Большой, Мэриэнн К. Дж. (2009). «Радужность фотонных кристаллов и ее подавление в чешуе бабочки». Журнал интерфейса Королевского общества. 6 (Приложение 2): S233 – S242. Дои:10.1098 / rsif.2008.0353.focus. ЧВК 2706480. PMID 18980932.
- ^ Argyros, A .; Manos, S .; Large, M. C. J .; McKenzie, D. R .; Cox, G.C .; Дварт, Д. М. (2002). «Электронная томография и компьютерная визуализация трехмерного« фотонного »кристалла в масштабе крыла бабочки». Микрон. 33 (5): 483–487. Дои:10.1016 / S0968-4328 (01) 00044-0. PMID 11976036.
- ^ Гираделла, Хелен (1991). «Свет и цвет на крыле: структурные цвета у бабочек и мотыльков». Прикладная оптика. 30 (24): 3492–3500. Bibcode:1991ApOpt..30.3492G. Дои:10.1364 / AO.30.003492. PMID 20706416.
- ^ Винтер-Блит, М.А. (1957). Бабочки Индийского региона (Перепечатка 2009 года издательством Today & Tomorrows Publishers, ред. Нью-Дели). Мумбаи, Индия: Бомбейское общество естественной истории. ISBN 978-81-7019-232-9.
- ^ «Андроконий». Энциклопедия Britannica Online. Получено 30 октября 2010.
- ^ Холл, Джейсон П. У .; Харви, Дональд Дж. (2002). «Обследование андрокониальных органов у Riodinidae (Lepidoptera)» (PDF). Зоологический журнал Линнеевского общества. 136 (2): 171–197. Дои:10.1046 / j.1096-3642.2002.00003.x.
- ^ Комсток, Джон Генри (2008) [1920]. Введение в энтомологию. Прочтите книги, первоначально опубликованные издательством Comstock Publishing Company. ISBN 978-1-4097-2903-7.
- ^ Скотт, Джеймс А (1997). Бабочки Северной Америки: естественная история и полевой справочник. Стэнфорд, Калифорния: Издательство Стэнфордского университета. ISBN 978-0804720137. OCLC 49698782.
- ^ Скобл (2005). Глава «Высшая Дитрисия», стр. 328.
- ^ а б c d е «Чешуекрылое». Энциклопедия Britannica Online. Получено 16 ноября 2010.
- ^ а б c d Скобл (1995). Раздел «Живот взрослого», (стр. 98–102).
- ^ Уотсон, Трэйси (3 июля 2013 г.). "Ястребиные соколы поражают летучих мышей звуковыми взрывами из их гениталий". Природа. Дои:10.1038 / природа.2013.13333. S2CID 180859622. Получено 5 июля 2013.
- ^ Дагдейл, Дж. (1974). «Конфигурация женских половых органов в классификации Lepidoptera». Новозеландский зоологический журнал. Королевское общество Новой Зеландии. 1 (2): 132. Дои:10.1080/03014223.1974.9517821. Получено 3 мая 2020.
- ^ а б c d е Скобл (1995). Глава «Незрелые стадии», (стр. 104–133).
- ^ а б Нация, Джеймс Л. (2002). Физиология и биохимия насекомых. CRC Press. ISBN 978-0-8493-1181-9.
- ^ а б c d е Чепмен (1998). Раздел «Яйцо и эмбриология» (стр. 325–362).
- ^ а б c Холланд, У. Дж. (1898). "Вступление" (PDF ). Книга бабочек. Лондон: Хатчинсон и Ко. ISBN 978-0-665-13041-0.
- ^ П. Дж. Гуллан; П. С. Крэнстон (2010). «Образцы и фазы жизненного цикла». Насекомые: очерк энтомологии (4-е изд.). Wiley-Blackwell. стр.156 –164. ISBN 978-1-4443-3036-6.
- ^ а б c d е ж Вагнер, Дэвид Л. (2005). Гусеницы востока Северной Америки. Princeton University Press. ISBN 978-0-691-12144-4.
- ^ а б Миллер, Джеффри К. (3 августа 2006 г.). «Морфология гусеницы». Гусеницы лесов и редколесий северо-запада Тихого океана. Центр исследования дикой природы северных прерий. Получено 16 ноября 2010.
- ^ Макослейн, Хизер Дж. (2008). «Апосематизм». В Capinera, Джон Л. (ред.). Энциклопедия энтомологии. Виртуальная справочная библиотека Gale. 4 (2-е изд.). Ссылка Springer. ISBN 978-1-4020-6242-1.
- ^ а б c d Общие, И. Ф. Б. (1990). Бабочки Австралии. Brill Publishers. ISBN 978-90-04-09227-3.
- ^ Харпер, Дуглас. "Хризалис". Интернет-словарь этимологии. Dictionary.com. Получено 16 ноября 2010.
- ^ Штер, Фредерик В. (2009). «Куколка и пупарий». In Resh, Vincent H .; Cardé, Ring T. (ред.). Энциклопедия насекомых (2-е изд.). Академическая пресса. С. 970–973. ISBN 978-0-12-374144-8.
- ^ Фигье, Луи (1868). Мир насекомых: популярное описание отрядов насекомых, вместе с описанием привычек и хозяйствования некоторых из наиболее интересных видов. Нью-Йорк: D. Appleton & Co.
- ^ Сураков, Андрей (2008). «Спаривание куколки у зебры длиннокрылых (Геликоний харитония): Фотографические свидетельства ". Новости общества лепидоптерологов. 50 (1): 26–32.
- ^ а б c «Защитные механизмы гусеницы и бабочки». EnchantedLearning.com. Получено 7 декабря 2009.
- ^ Латимер, Джонатан П .; Карен Стрэй Нолтинг (2000). Бабочки. Houghton Mifflin Harcourt. п.12. ISBN 978-0-395-97944-0.
Махаон тигровый.
- ^ Кричер, Джон (1999). "6". Неотропический компаньон. Princeton University Press. С. 157–158. ISBN 978-0-691-00974-2.
- ^ Santos, J.C .; Каннателла, Д. К. (2003). «Множественные, повторяющиеся причины апосематизма и специализации диеты на ядовитых лягушках» (PDF). Труды Национальной академии наук. 100 (22): 12792–12797. Bibcode:2003ПНАС..10012792С. Дои:10.1073 / pnas.2133521100. ЧВК 240697. PMID 14555763. Архивировано из оригинал (PDF) 24 июля 2011 г.. Получено 19 февраля 2011.
- ^ Насекомые и пауки мира. 10. Корпорация Маршалла Кавендиша. Маршалл Кавендиш. Январь 2003. С. 292–293. ISBN 978-0-7614-7344-2.CS1 maint: другие (связь)
- ^ Кэрролл, Шон (2005). Бесконечные самые прекрасные формы: новая наука Evo Devo и создание животного мира. W. W. Norton & Co., стр.205 –210. ISBN 978-0-393-06016-4.
Защита глазных пятен бабочки.
- ^ Хеффернан, Эмили (2004). Симбиотические отношения между Anthene emolus (Lycaenidae) и Oecophylla smaragdina (Formicidae): обязательный мутуализм в тропических лесах Малайзии (PDF) (Магистр Тезис). Университет Флориды.
- ^ «Осметериум». Merriam-Webster, Incorporated. Получено 9 декабря 2009.
- ^ Хэдли, Дебби. «Осметериум». About.com Руководство. Получено 9 декабря 2009.
внешняя ссылка
- СЭМ изображение чешуи бабочки и ее цветоножки (третье сверху).
- Изысканные кораблики - фоторепортаж о яйцах чешуекрылых. Национальная география.
- Необычное зрение - фоторепортаж о мотыльках. Национальная география.