Нейрофармакология - Neuropharmacology
Нейрофармакология это исследование того, как лекарственные препараты влияют на клеточные функции нервной системы и нервные механизмы, посредством которых они влияют на поведение.[1] Существует два основных раздела нейрофармакологии: поведенческая и молекулярная. Поведенческая нейрофармакология фокусируется на изучении того, как лекарства влияют на поведение человека (нейропсихофармакология ), включая изучение того, как лекарственная зависимость и зависимость влиять на человеческий мозг.[2] Молекулярная нейрофармакология предполагает изучение нейронов и их нейрохимический взаимодействия, с общей целью разработки лекарств, которые положительно влияют на неврологическую функцию. Оба эти поля тесно связаны, поскольку оба связаны с взаимодействием нейротрансмиттеры, нейропептиды, нейрогормоны, нейромодуляторы, ферменты, вторые посланники, совместные перевозчики, ионные каналы, и рецепторные белки в центральный и периферическая нервная система. Изучая эти взаимодействия, исследователи разрабатывают лекарства для лечения множества различных неврологических расстройств, в том числе боль, нейродегенеративные заболевания, такие как болезнь Паркинсона и Болезнь Альцгеймера, психологические расстройства, наркозависимость и многие другие.
История
Нейрофармакология не появлялась в научной сфере до тех пор, пока в начале 20-го века ученые не смогли составить базовое представление о нервной системе и о том, как нервы взаимодействуют друг с другом. До этого открытия были обнаружены лекарства, которые продемонстрировали какое-то влияние на нервную систему. В 1930-х годах французские ученые начали работать с соединением под названием фенотиазин в надежде синтезировать лекарство, способное бороться с малярией. Хотя использование этого препарата против людей, инфицированных малярией, показало очень мало надежды, было обнаружено, что он обладает седативным действием, а также оказывает положительное воздействие на пациентов с болезнью Паркинсона. Этот метод черного ящика, при котором исследователь вводил лекарство и изучал реакцию, не зная, как связать действие лекарства с реакцией пациента, был основным подходом в этой области, пока в конце 1940-х и начале 1950-х годов ученые не смогли идентифицировать специфические нейротрансмиттеры, такие как норэпинефрин (участвует в сужении кровеносных сосудов и учащении пульса и артериального давления), дофамин (химическое вещество, нехватка которого вызывает болезнь Паркинсона), и серотонин (вскоре будет признан глубоко связанным с депрессией[нужна цитата ]). В 1950-х годах ученые также стали лучше измерять уровни конкретных нейрохимических веществ в организме и, таким образом, соотносить эти уровни с поведением.[3] Изобретение зажим напряжения в 1949 г. позволили изучить ионные каналы и нервную потенциал действия. Эти два важных исторических события в нейрофармакологии позволили ученым не только изучить, как информация передается от одного нейрона к другому, но и изучить, как нейрон обрабатывает эту информацию внутри себя.
Обзор
Нейрофармакология - это очень обширная область науки, которая охватывает многие аспекты нервной системы, от манипуляций с отдельными нейронами до целых областей головного и спинного мозга и периферических нервов. Чтобы лучше понять основу разработки лекарств, нужно сначала понять, как нейроны взаимодействуют друг с другом.
Нейрохимические взаимодействия

Чтобы понять потенциальные успехи в медицине, которые может принести нейрофармакология, важно понимать, как человеческое поведение и мыслительные процессы передаются от нейрона к нейрону и как лекарства могут изменять химические основы этих процессов.
Нейроны известны как возбудимые клетки, потому что на их поверхностной мембране имеется множество белков, известных как ионные каналы, которые позволяют мелким заряженным частицам проходить внутрь и из клетки. Структура нейрона позволяет ему получать химическую информацию. дендриты, распространяется через перикарион (тело клетки) и вниз по его аксон, и, в конечном итоге, переходя к другим нейронам через свои аксонный терминал. Эти потенциалзависимые ионные каналы позволяют быстро деполяризация по всей камере. Эта деполяризация, если она достигнет определенного порога, вызовет потенциал действия. Как только потенциал действия достигает конца аксона, он вызывает приток ионов кальция в клетку. Ионы кальция затем вызывают везикулы, маленькие пакеты, заполненные нейротрансмиттеры, чтобы связываться с клеточной мембраной и выпускать ее содержимое в синапс. Эта клетка известна как пресинаптический нейрон, а клетка, которая взаимодействует с высвобождаемыми нейротрансмиттерами, известна как постсинаптический нейрон. После того, как нейротрансмиттер попадает в синапс, он может либо связываться с рецепторами постсинаптической клетки, либо пресинаптическая клетка может повторно захватить его и сохранить для последующей передачи, либо он может быть разрушен ферментами в синапсе. специфический для этого определенного нейротрансмиттера. Эти три различных действия являются основными областями, в которых действие лекарства может влиять на связь между нейронами.[3]
Есть два типа рецепторов, с которыми нейротрансмиттеры взаимодействуют на постсинаптическом нейроне. Первыми типами рецепторов являются ионные каналы, управляемые лигандами, или LGIC. Рецепторы LGIC - это самые быстрые типы преобразования химического сигнала в электрический. Как только нейротрансмиттер связывается с рецептором, он вызывает конформационные изменения, которые позволяют ионам напрямую проникать в клетку. Второй тип известен как рецепторы, связанные с G-белком, или GPCR. Они намного медленнее, чем LGIC, из-за увеличения количества биохимических реакций, которые должны происходить внутриклеточно. Как только нейротрансмиттер связывается с белком GPCR, он вызывает каскад внутриклеточных взаимодействий, которые могут приводить ко многим различным типам изменений клеточной биохимии, физиологии и экспрессии генов. Взаимодействие нейротрансмиттер / рецептор в области нейрофармакологии чрезвычайно важно, потому что многие лекарства, которые разрабатываются сегодня, связаны с нарушением этого процесса связывания.[4]
Молекулярная нейрофармакология
Молекулярная нейрофармакология включает изучение нейронов и их нейрохимических взаимодействий, а также рецепторов на нейронах с целью разработки новых лекарств, которые будут лечить неврологические расстройства, такие как боль, нейродегенеративные заболевания и психологические расстройства (также известные в данном случае как нейропсихофармакология ). Есть несколько технических терминов, которые необходимо определить, когда связывают нейротрансмиссию с действием рецепторов:
- Агонист - молекула, которая связывается с рецепторным белком и активирует этот рецептор.
- Конкурентный антагонист - молекула, которая связывается с тем же участком рецепторного белка, что и агонист, предотвращая активацию рецептора.
- Неконкурентный антагонист - молекула, которая связывается с рецепторным белком на другом сайте, чем у агониста, но вызывает конформационные изменения в белке, которые не допускают активации.
На следующие взаимодействия нейротрансмиттер / рецептор могут влиять синтетические соединения, которые действуют как одно из трех вышеуказанных. Каналы ионов натрия / калия также можно манипулировать в нейроне, чтобы вызвать ингибирующие эффекты потенциалов действия.
ГАМК
Нейромедиатор ГАМК обеспечивает быстрое синаптическое торможение в центральной нервной системе. Когда ГАМК высвобождается из пресинаптической клетки, она связывается с рецептором (скорее всего, ГАМКА рецептор), который вызывает гиперполяризацию постсинаптической клетки (оставаться ниже порога ее потенциала действия). Это будет противодействовать эффекту любой возбуждающей манипуляции со стороны других взаимодействий нейромедиатора / рецептора.
Это ГАМКА рецептор содержит множество сайтов связывания, которые допускают конформационные изменения и являются основной мишенью для разработки лекарств. Самый распространенный из этих сайтов связывания, бензодиазепин, допускает как агонистические, так и антагонистические эффекты на рецептор. Обычный препарат, диазепам, действует как аллостерический усилитель на этом сайте связывания.[5] Другой рецептор ГАМК, известный как ГАМКB, может быть усилен молекулой баклофена. Эта молекула действует как агонист, поэтому активирует рецептор и, как известно, помогает контролировать и уменьшать спастические движения.
Дофамин
В дофамин нейротрансмиттер опосредует синаптическую передачу, связываясь с пятью специфическими GPCR. Эти пять рецепторных белков разделены на два класса в зависимости от того, вызывает ли ответ возбуждающий или тормозной ответ на постсинаптическую клетку. Существует множество видов наркотиков, легальных и незаконных, которые влияют на дофамин и его взаимодействие с мозгом. При болезни Паркинсона, заболевании, которое снижает количество дофамина в головном мозге, пациенту назначают предшественник дофамина Леводопа, поскольку дофамин не может проникать через гематоэнцефалический барьер а л-допа можно. Некоторые агонисты дофамина также назначаются пациентам с болезнью Паркинсона, имеющей заболевание, известное как синдром беспокойных ног или RLS. Некоторые примеры из них: ропинироль и прамипексол.[6]
Психологические расстройства, подобные Синдром дефицита внимания и гиперактивности (СДВГ) можно лечить такими лекарствами, как метилфенидат (также известный как Риталин ), которые блокируют повторный захват дофамина пресинаптической клеткой, тем самым обеспечивая увеличение дофамина, оставшегося в синаптической щели. Это увеличение синаптического дофамина увеличивает связывание с рецепторами постсинаптической клетки. Этот же механизм также используется другими запрещенными и более сильнодействующими стимуляторами, такими как кокаин.
Серотонин
Нейромедиатор серотонин обладает способностью опосредовать синаптическую передачу через рецепторы GPCR или LGIC. Возбуждающие или тормозящие постсинаптические эффекты серотонина определяются типом рецептора, экспрессируемого в данной области мозга. Наиболее популярные и широко используемые препараты для регуляции уровня серотонина во время депрессии известны как СИОЗС или селективные ингибиторы обратного захвата серотонина. Эти препараты подавляют транспорт серотонина обратно в пресинаптический нейрон, оставляя больше серотонина в синаптической щели.
До открытия СИОЗС были также лекарства, которые подавляли фермент, расщепляющий серотонин. MAOI или ингибиторы моноаминоксидазы увеличивал количество серотонина в синапсе, но имел много побочных эффектов, включая сильную мигрень и высокое кровяное давление. В конечном итоге это было связано с тем, что лекарства взаимодействуют с обычным химическим веществом, известным как тирамин содержится во многих продуктах питания.[7]
Ионные каналы
Ионные каналы, расположенные на поверхностной мембране нейрона, обеспечивают приток ионов натрия и движение ионов калия наружу во время потенциала действия. Избирательная блокировка этих ионных каналов снизит вероятность возникновения потенциала действия. Наркотик рилузол - нейропротекторный препарат, блокирующий ионные каналы натрия. Поскольку эти каналы не могут активироваться, отсутствует потенциал действия, и нейрон не выполняет никакого преобразования химических сигналов в электрические, и сигнал не передается дальше. Этот препарат используется как обезболивающее, а также как успокаивающее средство.[8]
Поведенческая нейрофармакология
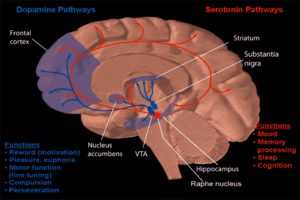
Одна из форм поведенческой нейрофармакологии фокусируется на изучении лекарственной зависимости и того, как наркомания влияет на человеческий разум. Большинство исследований показало, что основная часть мозга, которая усиливает зависимость посредством нейрохимического вознаграждения, - это прилежащее ядро. Изображение справа показывает, как дофамин проецируется в эту область. Хроническое злоупотребление алкоголем может вызвать зависимость и зависимость. Как возникает эта зависимость, описано ниже.
Этиловый спирт
Алкоголь вознаграждает и укрепляет (т. Е. вызывающий привыкание ) свойства опосредуются через его влияние на дофамин нейроны в мезолимбический путь вознаграждения, который соединяет вентральная тегментальная область к прилежащее ядро (NAcc).[9][10] Одним из основных эффектов алкоголя является аллостерическое подавление Рецепторы NMDA и содействие ГАМКА рецепторы (например, улучшенная ГАМКА рецептор-опосредованный хлористый поток через аллостерическая регуляция рецептора).[11] В высоких дозах этанол подавляет большинство лигандные ионные каналы и ионные каналы, управляемые напряжением в нейронах тоже.[11] Алкоголь подавляет натриево-калиевые насосы в мозжечок и, вероятно, именно так это ухудшает работу мозжечка и координацию тела.[12][13]
При остром употреблении алкоголя дофамин выпущен в синапсы мезолимбического пути, что, в свою очередь, усиливает активацию постсинаптического Рецепторы D1.[9][10] Активация этих рецепторов запускает постсинаптические внутренние сигнальные события через протеинкиназа А что в конечном итоге фосфорилат белок, связывающий элемент ответа цАМФ (CREB), вызывая CREB-опосредованные изменения в экспрессия гена.[9][10]
При хроническом употреблении алкоголя потребление этанола аналогичным образом вызывает фосфорилирование CREB через путь рецептора D1, но также изменяет Рецептор NMDA функционируют через механизмы фосфорилирования;[9][10] адаптивный подавление пути рецептора D1 и функции CREB.[9][10] Хроническое потребление также связано с влиянием на фосфорилирование и функцию CREB через постсинаптические сигнальные каскады рецепторов NMDA через Путь MAPK / ERK и CAMK -опосредованный путь.[10] Эти модификации функции CREB в мезолимбическом пути вызвать выражение (т.е. увеличить экспрессию генов) ΔFosB в NAcc,[10] где ΔFosB - это «главный контрольный белок», который при сверхэкспрессии в NAcc является необходимо и достаточно для развития и поддержания аддиктивного состояния (то есть его сверхэкспрессия в прилежащем ядре производит, а затем напрямую модулирует компульсивное потребление алкоголя).[10][14][15][16]
Исследование
болезнь Паркинсона
болезнь Паркинсона это нейродегенеративное заболевание описывается избирательной потерей дофаминергический нейроны, расположенные в черная субстанция. Сегодня наиболее часто применяемым препаратом для борьбы с этим заболеванием является леводопа или L-ДОПА. Этот предшественник дофамина может проникать через гематоэнцефалический барьер, тогда как нейромедиатор дофамин не может. Было проведено обширное исследование, чтобы определить, является ли L-допа лучшим средством лечения болезни Паркинсона по сравнению с другими агонистами дофамина. Некоторые считают, что длительное использование L-допы нарушит нейрозащиту и, таким образом, в конечном итоге приведет к гибели дофаминергических клеток. Хотя доказательств не было, in vivo или же in vitro, некоторые до сих пор считают, что длительный прием агонистов дофамина лучше для пациента.[17]
Болезнь Альцгеймера
Хотя существует множество гипотез, объясняющих причину Болезнь Альцгеймера, знания об этом заболевании далеко не полны для объяснения, что затрудняет разработку методов лечения. Известно, что в головном мозге пациентов с болезнью Альцгеймера как нейрональные никотиновые рецепторы ацетилхолина (nACh), так и рецепторы NMDA подавляются. Таким образом, четыре антихолинэстеразы были разработаны и одобрены Управление по санитарному надзору за качеством пищевых продуктов и медикаментов США (FDA) для лечения в США. Однако это не идеальные препараты, учитывая их побочные эффекты и ограниченную эффективность. Один многообещающий препарат, нефирацетам, разрабатывается для лечения болезни Альцгеймера и других пациентов с деменцией и обладает уникальными действиями по усилению активности как рецепторов nACh, так и рецепторов NMDA.[18]
Будущее
С развитием технологий и нашим пониманием нервной системы разработка лекарств будет продолжаться с увеличением количества лекарств. чувствительность и специфичность. Отношения структура-деятельность являются важной областью исследований в нейрофармакологии; попытка изменить действие или эффективность (то есть активность) биологически активных химических соединений путем изменения их химической структуры.[8]
Смотрите также
Рекомендации
- ^ Юнг А.В.К., Цветков Н.Т., Атанасов А.Г. Когда неврология встречается с фармакологией: анализ литературы по нейрофармакологии. Front Neurosci. 2018 16 ноября; 12: 852. DOI: 10.3389 / fnins.2018.00852.
- ^ Эверитт, Б. Дж.; Роббинс, Т. (2005). «Нейронные системы подкрепления при наркомании: от действий к привычкам и принуждению». Природа Неврологии. 8 (11): 1481–1489. Дои:10.1038 / nn1579. PMID 16251991.
- ^ а б Вробель, С. (2007). «Наука, серотонин и печаль: биология антидепрессантов: серия для общественности». Журнал FASEB. 21 (13): 3404–17. Дои:10.1096 / fj.07-1102 мкФ. PMID 17967927.
- ^ Ловингер, Д. М. (2008). «Коммуникационные сети в нейронах мозга, рецепторах, нейротрансмиттерах и алкоголе. [Обзор]». Исследования алкоголя и здоровье. 31 (3): 196–214.
- ^ Сигель, Э (2002). «Картирование сайта распознавания бензодиазепина на рецепторах ГАМК (А)». Актуальные темы медицинской химии. 2 (8): 833–9. Дои:10.2174/1568026023393444. PMID 12171574.
- ^ Винкельман, JW; Allen, RP; Tenzer, P; Хенинг, W (2007). «Синдром беспокойных ног: нефармакологические и фармакологические методы лечения». Гериатрия. 62 (10): 13–6. PMID 17922563.
- ^ Лопес-Муньос, Ф .; Аламо, К. (2009). «Моноаминергическая нейротрансмиссия: история открытия антидепрессантов с 1950-х годов до наших дней». Текущий фармацевтический дизайн. 15 (14): 1563–1586. Дои:10.2174/138161209788168001. PMID 19442174.
- ^ а б Нарахаши, Т. (2000). «Нейрорецепторы и ионные каналы как основа действия лекарств: прошлое, настоящее и будущее». Журнал фармакологии и экспериментальной терапии. 294 (1): 1–26. PMID 10871290.
- ^ а б c d е «Алкоголизм - запись в базе данных Homo sapiens (человек)». KEGG Pathway. 29 октября 2014 г.. Получено 9 февраля 2015.
В качестве одного из основных медиаторов положительных эффектов алкоголя были идентифицированы дофаминергические проекции вентральной тегментальной области (VTA) на прилежащее ядро (NAc). Острое воздействие алкоголя стимулирует выброс дофамина в NAc, который активирует рецепторы D1, стимулируя передачу сигналов PKA и последующую опосредованную CREB экспрессию гена, тогда как хроническое воздействие алкоголя приводит к адаптивному подавлению этого пути, в частности функции CREB. Снижение функции CREB в NAc может способствовать употреблению наркотиков, вызывающих злоупотребление, для достижения увеличения вознаграждения и, таким образом, может участвовать в регулировании положительных аффективных состояний зависимости. Передача сигналов PKA также влияет на активность рецептора NMDA и может играть важную роль в нейроадаптации в ответ на хроническое воздействие алкоголя.
- ^ а б c d е ж грамм час Kanehisa Laboratories (29 октября 2014 г.). «Алкоголизм - Homo sapiens (человек)». KEGG Pathway. Получено 31 октября 2014.
- ^ а б Маленка Р.К., Нестлер Э.Дж., Хайман С.Е. (2009). «Глава 15: Закрепление и аддиктивные расстройства». В Sydor A, Brown RY (ред.). Молекулярная нейрофармакология: основа клинической неврологии (2-е изд.). Нью-Йорк: McGraw-Hill Medical. п. 372. ISBN 9780071481274.
Несмотря на высокие концентрации, необходимые для его психоактивных эффектов, этанол оказывает специфическое действие на мозг. Первоначальные эффекты этанола являются результатом, прежде всего, облегчения рецепторов GABAA и ингибирования рецепторов глутамата NMDA. В более высоких дозах этанол также подавляет функционирование большинства лигандных и потенциалзависимых ионных каналов. Неизвестно, влияет ли этанол избирательно на эти каналы через прямое низкоаффинное связывание или через неспецифическое разрушение плазматических мембран, которое затем избирательно влияет на эти очень сложные, мультимерные, трансмембранные белки. Этанол аллостерически регулирует рецептор GABAA для усиления потока Cl-, активируемого GABA. Анксиолитическое и седативное действие этанола, а также барбитуратов и бензодиазепинов является результатом усиления ГАМКергической функции. Также считается, что облегчение функции рецептора ГАМК способствует усилению эффектов этих препаратов. Не все рецепторы GABAA чувствительны к этанолу. ... Этанол также действует как антагонист NMDA, аллостерически подавляя прохождение активированных глутаматом токов Na + и Ca2 + через рецептор NMDA. ... Усиливающие эффекты этанола частично объясняются его способностью активировать мезолимбические схемы дофамина, хотя неизвестно, опосредуется ли этот эффект на уровне VTA или NAc. Также неизвестно, вызвана ли эта активация дофаминовых систем главным образом облегчением рецепторов ГАМКА или ингибированием рецепторов NMDA, или обоими. Усиление этанолом также частично опосредовано этанолом высвобождением эндогенных опиоидных пептидов в мезолимбической дофаминовой системе, хотя пока не известно, является ли VTA или NAc преобладающим местом такого действия. Соответственно, антагонист опиоидных рецепторов налтрексон снижает самовведение этанола животными и используется с умеренным эффектом для лечения алкоголизма у людей.
- ^ Форрест, доктор медицины (апрель 2015 г.). «Моделирование действия алкоголя на подробной модели нейрона Пуркинье и более простой суррогатной модели, которая работает в> 400 раз быстрее». BMC Neuroscience. 16 (27): 27. Дои:10.1186 / s12868-015-0162-6. ЧВК 4417229. PMID 25928094.
- ^ Форрест, Майкл (апрель 2015 г.). "Неврология, почему мы падаем в пьяном виде". Наука 2.0. Получено 2 января, 2019.
- ^ Ruffle JK (ноябрь 2014 г.). «Молекулярная нейробиология зависимости: о чем вообще (Δ) FosB?». Am J Злоупотребление алкоголем. 40 (6): 428–437. Дои:10.3109/00952990.2014.933840. PMID 25083822.
ΔFosB как терапевтический биомаркер
Сильная корреляция между хроническим воздействием лекарств и ΔFosB открывает новые возможности для таргетной терапии при зависимости (118) и предлагает методы анализа их эффективности (119). За последние два десятилетия исследования продвинулись от определения индукции ΔFosB к изучению его последующего действия (38). Вероятно, что исследования ΔFosB теперь перейдут в новую эру - использование ΔFosB в качестве биомаркера. Если обнаружение ΔFosB указывает на хроническое воздействие препарата (и, по крайней мере, частично отвечает за зависимость от вещества), то его мониторинг терапевтической эффективности в интервенционных исследованиях является подходящим биомаркером (рис. 2). Здесь обсуждаются примеры терапевтических возможностей. ...
Выводы
ΔFosB является важным фактором транскрипции, участвующим в молекулярных и поведенческих механизмах привыкания после многократного воздействия наркотиков. Образование ΔFosB во многих областях мозга и молекулярный путь, ведущий к образованию комплексов AP-1, хорошо изучены. Установление функционального назначения ΔFosB позволило дополнительно определить некоторые ключевые аспекты его молекулярных каскадов, включая такие эффекторы, как GluR2 (87,88), Cdk5 (93) и NFkB (100). Более того, многие из этих выявленных молекулярных изменений теперь напрямую связаны со структурными, физиологическими и поведенческими изменениями, наблюдаемыми после хронического воздействия лекарств (60,95,97,102). Эпигенетические исследования открыли новые горизонты исследований молекулярной роли ΔFosB, а недавние достижения продемонстрировали роль ΔFosB, действующего на ДНК и гистоны, действительно как «молекулярный переключатель» (34). Благодаря нашему более глубокому пониманию ΔFosB при зависимости, появилась возможность оценить вызывающий привыкание потенциал текущих лекарств (119), а также использовать его в качестве биомаркера для оценки эффективности терапевтических вмешательств (121, 122, 124). Некоторые из этих предложенных вмешательств имеют ограничения (125) или находятся в зачаточном состоянии (75). Однако есть надежда, что некоторые из этих предварительных результатов могут привести к инновационным методам лечения, которые так необходимы при зависимости. - ^ Нестлер EJ (декабрь 2013 г.). «Клеточная основа памяти при зависимости». Диалоги Clin Neurosci. 15 (4): 431–443. ЧВК 3898681. PMID 24459410.
НЕСМОТРЯ НА ВАЖНОСТЬ МНОГОЧИСЛЕННЫХ ПСИХОСОЦИАЛЬНЫХ ФАКТОРОВ, В СВОЕЙ ОСНОВНОЙ ЦЕЛЯХ НАРКОТИКАЛЬНАЯ ЗАВИСИМОСТЬ ВКЛЮЧАЕТ БИОЛОГИЧЕСКИЙ ПРОЦЕСС: способность многократного воздействия наркотика, вызывающего злоупотребление, вызывать изменения в уязвимом мозге, которые вызывают компульсивный поиск и прием наркотиков, и потерю контроля над употреблением наркотиков, которые определяют состояние зависимости. ... Большое количество литературы продемонстрировало, что такая индукция ΔFosB в нейронах NAc D1-типа увеличивает чувствительность животного к лекарству, а также увеличивает естественное вознаграждение и способствует самостоятельному введению лекарства, предположительно за счет процесса положительного подкрепления.
- ^ Робисон А.Дж., Нестлер Э.Дж. (ноябрь 2011 г.). «Транскрипционные и эпигенетические механизмы зависимости». Nat. Преподобный Neurosci. 12 (11): 623–637. Дои:10.1038 / nrn3111. ЧВК 3272277. PMID 21989194.
ΔFosB был непосредственно связан с несколькими зависимостями поведения ... Важно отметить, что генетическая или вирусная сверхэкспрессия ΔJunD, доминантно-отрицательного мутанта JunD, который противодействует ΔFosB- и другой AP-1-опосредованной транскрипционной активности, в NAc или OFC блокирует их. ключевые эффекты воздействия наркотиков14,22–24. Это указывает на то, что ΔFosB необходим и достаточен для многих изменений, вызванных в мозге хроническим воздействием лекарств. ΔFosB также индуцируется в MSN NAc D1-типа при хроническом потреблении нескольких естественных наград, включая сахарозу, пищу с высоким содержанием жира, секс, бег колеса, где он способствует этому потреблению.14,26–30. Это означает, что ΔFosB участвует в регуляции естественного вознаграждения в нормальных условиях и, возможно, во время состояний, подобных патологическому привыканию. ... ΔFosB служит одним из основных контрольных белков, регулирующих эту структурную пластичность.
- ^ Shin, J. Y .; Park, H.J .; Ahn, Y.H .; Ли, П. Х. (2009). «Нейропротекторное действие l-допа на дофаминергические нейроны сравнимо с прамипексолом в модели болезни Паркинсона на животных, получавших МРТР: исследование прямого сравнения». Журнал нейрохимии. 111 (4): 1042–50. Дои:10.1111 / j.1471-4159.2009.06381.x. PMID 19765187.
- ^ Нарахаши, Т; Маршалек, Вт; Моригути, S; Ага, JZ; Чжао, X (2003). «Уникальный механизм действия препаратов Альцгеймера на никотиновые ацетилхолиновые рецепторы и рецепторы NMDA в головном мозге». Науки о жизни. 74 (2–3): 281–91. Дои:10.1016 / j.lfs.2003.09.015. PMID 14607256.
внешняя ссылка
 СМИ, связанные с Нейрофармакология в Wikimedia Commons
СМИ, связанные с Нейрофармакология в Wikimedia Commons
