Черная субстанция - Substantia nigra - Wikipedia
| Черная субстанция | |
|---|---|
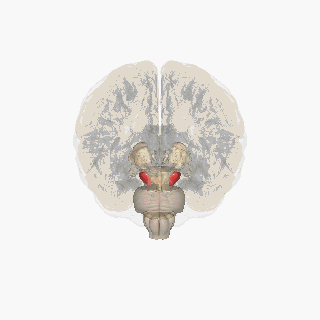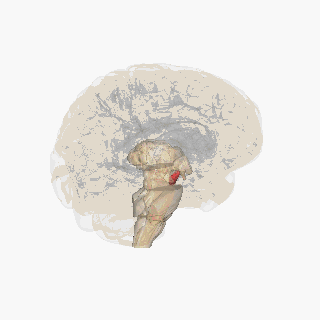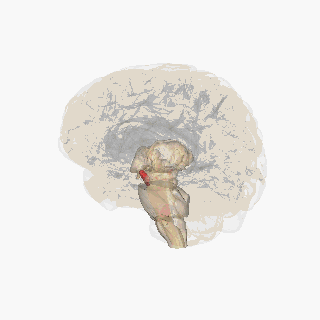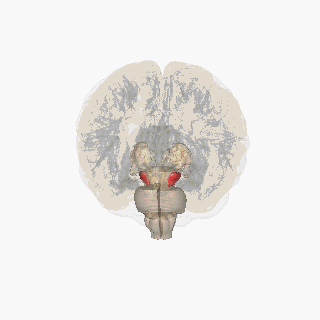 Черная субстанция выделена красным. | |
 Раздел через верхний холмик с изображением черной субстанции. | |
| Подробности | |
| Часть | Средний мозг, Базальный ганглий |
| Идентификаторы | |
| латинский | Черная субстанция |
| MeSH | D013378 |
| NeuroNames | 536 |
| НейроЛекс Я БЫ | birnlex_789 |
| TA98 | A14.1.06.111 |
| TA2 | 5881 |
| FMA | 67947 |
| Анатомические термины нейроанатомии | |
В черная субстанция (SN) это базальный ганглий структура, расположенная в средний мозг что играет важную роль в награда и движение. Черная субстанция является латинский для «черного вещества», что отражает тот факт, что части черной субстанции кажутся темнее, чем соседние области из-за высокого уровня нейромеланин в дофаминергический нейроны.[1] болезнь Паркинсона характеризуется потерей дофаминергических нейронов в черной субстанции pars compacta.[2]
Хотя черная субстанция выглядит как непрерывная полоса в срезах мозга, анатомические исследования показали, что на самом деле она состоит из двух частей с очень разными связями и функциями: pars compacta (SNpc) и pars reticulata (СНпр). Компактная часть служит в основном выступом к цепи базальных ганглиев, снабжая полосатое тело с дофамином. Pars reticulata передает сигналы от базальный ганглий к множеству других структур мозга.[3]
Структура
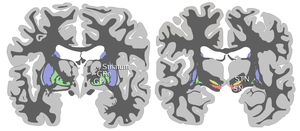

Черная субстанция вместе с четырьмя другими ядрами является частью базальный ганглий. Это самый большой ядро в среднем мозге, лежит дорсальнее церебральные ножки. У людей есть две черные субстанции, по одной с каждой стороны от средней линии.
SN разделен на две части: pars reticulata (СНпр) и pars compacta (SNpc), который лежит медиальнее ретикулатной части. Иногда упоминается третья область, латеральная часть, хотя обычно ее классифицируют как часть ретикулатной части. (SNpr) и внутренний бледный шар (GPi) разделены внутренняя капсула.[4]
Pars reticulata
Pars reticulata имеет сильное структурное и функциональное сходство с внутренней частью бледного шара. Их иногда считают частями одной и той же структуры, разделенной белым веществом внутренней капсулы. Как и у бледного шара, нейроны pars reticulata в основном ГАМКергический.[нужна цитата ]
Афферентные связи
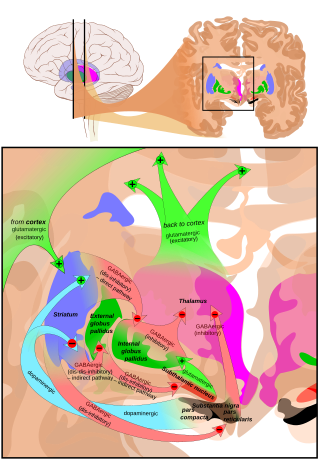
Главный вход в SNpr происходит от полосатое тело. Он идет двумя путями, известными как непосредственный и косвенные пути. Прямой путь состоит из аксонов от средних колючих клеток полосатого тела, которые проецируются непосредственно в pars reticulata. Непрямой путь состоит из трех звеньев: проекции колючих клеток полосатого тела на внешнюю часть бледный шар; а ГАМКергический проекция из бледного шара на субталамическое ядро, а глутаматергический проекция от субталамического ядра к pars reticulata.[5] Таким образом, активность полосатого тела через прямой путь оказывает ингибирующее действие на нейроны в (SNpr), но возбуждающее действие через непрямой путь. Прямые и непрямые пути берут начало от разных подгрупп шиповатых клеток полосатого тела: они тесно переплетены, но экспрессируют разные типы дофаминовых рецепторов, а также демонстрируют другие нейрохимические различия.
Эфферентные связи
Значительные выступы происходят в таламусе (вентрально-латеральное и вентральное переднее ядра), верхний холмик и другие каудальные ядра из pars reticulata (нигроталамический путь),[6] которые используют ГАМК в качестве своего нейромедиатора. Кроме того, эти нейроны образуют до пяти коллатералей, которые разветвляются как внутри pars compacta, так и pars reticulata, вероятно, модулируя дофаминергическую активность в pars compacta.[7]
Функция
Черная субстанция играет важную роль в работе мозга, в частности, в движение глаз, двигательное планирование, стремление к вознаграждению, учусь, и зависимость. Многие эффекты черной субстанции опосредованы полосатое тело. Ниграл дофаминергический вход в полосатое тело через нигростриальный путь тесно связан с функцией полосатого тела.[8] Взаимозависимость между полосатым телом и черной субстанцией можно увидеть следующим образом: когда черная субстанция электрически стимулируется, движения не происходит; тем не менее, симптомы дегенерации черной субстанции из-за болезни Паркинсона являются ярким примером влияния черной субстанции на движение. Помимо функций, опосредованных полосатым телом, черная субстанция также служит основным источником ГАМКергический подавление различных мишеней мозга.
Pars reticulata
В pars reticulata черной субстанции является важным центром обработки в базальных ганглиях. ГАМКергические нейроны в сетчатой части передают окончательные обработанные сигналы базальный ганглий к таламус и верхний холмик. Кроме того, pars reticulata также подавляет дофаминергическую активность в pars compacta через коллатерали аксонов, хотя функциональная организация этих связей остается неясной.
ГАМКергические нейроны ретикулатной части спонтанно возбуждаются. потенциалы действия. У крыс частота потенциалов действия составляет примерно 25 Гц.[9] Целью этих потенциалов спонтанного действия является подавление мишеней базальных ганглиев, а уменьшение подавления связано с движением.[10] Субталамическое ядро дает возбуждающий сигнал, который модулирует скорость возбуждения этих спонтанных потенциалов действия. Однако поражение субталамического ядра приводит только к 20% снижению частоты активации pars reticulata, что позволяет предположить, что генерация потенциалов действия в pars reticulata в значительной степени автономна.[11] на примере роли pars reticulata в саккадическое движение глаз. Группа ГАМКергических нейронов от pars reticulata проецируется в верхний холмик, демонстрируя высокий уровень устойчивой ингибирующей активности.[12] Прогнозы из хвостатое ядро к верхнему бугорку также модулируют саккадические движения глаз. Измененные модели стрельбы pars reticulata, такие как стрельба одиночными шипами или очередями, встречаются у болезнь Паркинсона[13] и эпилепсия.[14]
Pars compacta
Самая выдающаяся функция pars compacta - блок управления двигателем,[15] хотя роль черной субстанции в двигательном контроле косвенная; электрическая стимуляция черной субстанции не приводит к движению из-за посредничества полосатого тела во влиянии движения. Компактная часть посылает возбуждающий сигнал в полосатое тело через путь D1, который возбуждает и активирует полосатое тело, что приводит к высвобождению ГАМК на бледный шар, чтобы подавить его ингибирующие эффекты на ядро таламуса. Это приводит к возбуждению таламокортикальных путей и передаче сигналов двигательных нейронов к коре головного мозга, что позволяет инициировать движение, которое отсутствует при болезни Паркинсона. Однако отсутствие нейронов pars compacta оказывает большое влияние на движение, о чем свидетельствуют симптомы болезни Паркинсона. Моторная роль компактной части может включать мелкую моторику, что было подтверждено на животных моделях с поражениями в этой области.[16]
Компактная часть активно участвует в усвоении ответов на раздражители. У приматов активность дофаминергических нейронов в нигростриатном пути увеличивается при предъявлении нового стимула.[17] Дофаминергическая активность снижается при повторном предъявлении стимула.[17] Тем не менее, поведенчески значимое предъявление стимула (т.е. вознаграждения) продолжает активировать дофаминергические нейроны в компактной части черной субстанции. Дофаминергические проекции от вентральная тегментальная область (нижняя часть «среднего мозга» или среднего мозга) к префронтальной коре (мезокортикальный путь) и к прилежащему ядру (мезолимбический путь - «мезо», что означает «из среднего мозга» ... в частности, вентральная тегментальная область ) связаны с вознаграждением, удовольствием и вызывающим привыкание поведением. Компактная часть также важна в пространственном обучении, наблюдениях за окружающей средой и положением в пространстве. Повреждения компактной части тела приводят к дефициту обучения повторению идентичных движений,[18] и некоторые исследования указывают на его участие в системе памяти, зависящей от спинного стриатума и ответной реакции, которая функционирует относительно независимо от гиппокамп, который традиционно считается подчиненным пространственным или эпизодическая память функции.[19]
Компактная часть также играет роль в временная обработка и активируется во время воспроизведения времени. Поражения в pars compacta приводит к временному дефициту.[20] В последнее время подозревали, что pars compacta регулирует цикл сна и бодрствования,[21] что согласуется с такими симптомами, как бессонница и Быстрый сон расстройства, о которых сообщают пациенты с болезнь Паркинсона. Даже в этом случае частичный дефицит дофамина, который не влияет на моторный контроль, может привести к нарушениям цикла сна-бодрствования, особенно к REM-подобным паттернам нервной активности во время бодрствования, особенно в гиппокамп.[22]
Клиническое значение
Черная субстанция имеет решающее значение в развитии многих заболеваний и синдромов, в том числе: паркинсонизм и болезнь Паркинсона.
болезнь Паркинсона
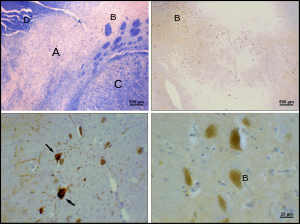
Болезнь Паркинсона - это нейродегенеративное заболевание частично характеризуется гибелью дофаминергических нейронов в SNpc. Основные симптомы болезни Паркинсона включают: тремор, акинезия, брадикинезия, и жесткость.[23] Другие симптомы включают нарушение осанки, усталость, нарушения сна, и в депрессии.[24]
Причина гибели дофаминергических нейронов в SNpc неизвестна. Тем не менее, некоторые вклады в уникальную восприимчивость дофаминергических нейронов в компактной части pars были идентифицированы. Во-первых, дофаминергические нейроны обнаруживают аномалии в митохондриальный комплекс 1, вызывая агрегацию альфа-синуклеин; это может привести к неправильной обработке белка и гибели нейронов.[25] Во-вторых, дофаминергические нейроны компактной части содержат меньше кальбиндин чем другие дофаминергические нейроны.[26] Кальбиндин белок, участвующий в кальций ионный транспорт внутри клеток, а избыток кальция в клетках токсичен. В кальбиндин Теория объяснила бы высокую цитотоксичность болезни Паркинсона в черной субстанции по сравнению с вентральной тегментальной областью. Независимо от причины гибели нейронов пластичность компактной части очень высока; Симптомы паркинсонизма не проявляются до тех пор, пока не погибнет до 50–80% дофаминергических нейронов pars compacta.[27] Большая часть этой пластичности происходит на нейрохимическом уровне; Транспортные системы дофамина замедляются, что позволяет дофамину задерживаться на более длительные периоды времени в химических синапсах в полосатом теле.[27]
Menke, Jbabdi, Miller, Matthews и Zari (2010) использовали визуализацию тензора диффузии, а также картографирование T1 для оценки объемных различий в SNpc и SNpr у участников с болезнью Паркинсона по сравнению со здоровыми людьми. Эти исследователи обнаружили, что участники с болезнью Паркинсона постоянно имели меньшую черную субстанцию, особенно в SNpr. Поскольку SNpr связан с задним таламусом, вентральным таламусом и, в частности, с моторной корой, и поскольку участники с болезнью Паркинсона сообщают о меньших SNprs (Menke, Jbabdi, Miller, Matthews and Zari, 2010), небольшой объем этой области может быть причиной двигательных нарушений у пациентов с болезнью Паркинсона. Этот небольшой объем может быть причиной более слабых и / или менее контролируемых двигательных движений, что может привести к тремору, который часто испытывают люди с болезнью Паркинсона.[28]
Окислительный стресс и окислительное повреждение SNpc, вероятно, являются ключевыми факторами в этиологии Болезнь Паркинсона как люди стареют.[29] Повреждения ДНК вызванные окислительным стрессом, могут быть отремонтирован процессами, модулируемыми альфа-синуклеин.[30] Альфа-синуклеин экспрессируется в черной субстанции, но ее Ремонт ДНК функция, похоже, нарушена в Тело Леви подшипник включения нейроны.[30] Эта потеря может вызвать гибель клеток.
Шизофрения
Повышенный уровень дофамина уже давно участвует в развитии шизофрения.[31] Однако по сей день вокруг этого дофаминовая гипотеза шизофрении. Несмотря на разногласия, антагонисты дофамина остаются стандартным и успешным средством лечения шизофрении. Эти антагонисты включают нейролептики первого поколения (типичные) Такие как бутирофеноны, фенотиазины, и тиоксантены. Эти препараты в значительной степени были заменены антипсихотические препараты второго поколения (атипичные) Такие как клозапин и палиперидон. Как правило, эти препараты действуют не на сами нейроны, продуцирующие дофамин, а на рецепторы постсинаптического нейрона.
Другие, нефармакологические доказательства в поддержку гипотезы дофамина, относящейся к черной субстанции, включают структурные изменения компактной части, такие как уменьшение размера синаптического терминала.[32] Другие изменения в черной субстанции включают повышенную экспрессию Рецепторы NMDA в черной субстанции и уменьшенной дисбиндин выражение. Увеличение рецепторов NMDA может указывать на участие глутамат -дофамин взаимодействия при шизофрении. Дисбиндин, который был (спорно), связанных с шизофренией, может регулировать высвобождение допамина, и низкое выражение Дисбиндин в черной субстанции могут играть важную роль в этиологии шизофрении.[33] Из-за изменений в черной субстанции в головном мозге шизофреника со временем может появиться возможность использовать определенные методы визуализации (такие как нейромеланин-специфическая визуализация) для обнаружения физиологических признаков шизофрении в черной субстанции.[34]
Синдром деревянного сундука
Деревянный сундук, также называемый синдромом жесткости грудной стенки фентанила, является редким побочным эффектом синтетического опиоиды Такие как Фентанил, Сульфентанил, Альфентанил, Ремифентанил. Это приводит к общему увеличению скелетной мышечный тонус. Считается, что этот механизм связан с повышенным высвобождением дофамина и пониженным высвобождением ГАМК в нервах черной субстанции / полосатого тела. Эффект наиболее выражен на мышцах грудной стенки и может привести к нарушению вентиляции. Это состояние чаще всего наблюдается при анестезии, когда быстрые и высокие дозы этих препаратов вводятся внутривенно.[нужна цитата ]
Множественная системная атрофия
Множественная системная атрофия характеризующийся дегенерацией нейронов в полосатом теле и черной субстанции, ранее назывался стриатонигральная дегенерация.
Химическая модификация черной субстанции
Химическая обработка и модификация черной субстанции важны в областях нейрофармакология и токсикология. Различные соединения, такие как леводопа и МФТП, используются в лечении и изучении болезни Паркинсона, а многие другие препараты действуют на черную субстанцию.
Амфетамин и следовые амины
Исследования показали, что в определенных областях мозга амфетамин и следовые амины повышают концентрацию дофамина в организме. синаптическая щель, тем самым усиливая реакцию постсинаптического нейрона.[35] Различные механизмы, с помощью которых амфетамин и следовые амины влияют на концентрацию дофамина, были тщательно изучены и, как известно, включают оба DAT и VMAT2.[35][36][37] Амфетамин похож по структуре на дофамин и следовые амины; как следствие, он может попасть в пресинаптический нейрон через DAT а также путем прямой диффузии через нервную мембрану.[35] При попадании в пресинаптический нейрон амфетамин и следовые амины активируют TAAR1, который через протеинкиназа сигнализация, вызывает отток дофамина, зависимый от фосфорилирования DAT интернализация и неконкурентное ингибирование обратного захвата.[35][38] Из-за сходства между амфетамином и следовыми аминами он также является субстратом для переносчиков моноаминов; как следствие, он (конкурентно) ингибирует обратный захват дофамина и других моноаминов, также конкурируя с ними за захват.[35]
Кроме того, амфетамин и следовые амины являются субстратами для нейронального переносчика везикулярных моноаминов, везикулярный переносчик моноаминов 2 (VMAT2).[37] Когда амфетамин принимает VMAT2везикула высвобождает (отталкивает) молекулы дофамина в цитозоль взамен.[37]
Кокаин
Кокаин Механизм действия в мозге человека включает ингибирование обратного захвата дофамина,[39] что объясняет вызывающие привыкание свойства кокаина, поскольку дофамин является критическим нейромедиатором вознаграждения. Однако кокаин более активен в дофаминергических нейронах вентральной тегментальной области, чем в черной субстанции. Прием кокаина увеличивает метаболизм в черной субстанции, что может объяснить изменение двигательной функции, наблюдаемое у субъектов, употребляющих кокаин.[40] Подавление обратного захвата дофамина кокаином также подавляет запуск спонтанных потенциалов действия компактной частью.[41] Механизм, с помощью которого кокаин ингибирует обратный захват дофамина, включает его связывание с переносчик дофамина белок. Однако исследования показывают, что кокаин также может вызывать снижение DAT. мРНК уровни[42] скорее всего, из-за того, что кокаин блокирует дофаминовые рецепторы, а не из-за прямого вмешательства в пути транскрипции или трансляции.[42]
Инактивация черной субстанции может оказаться возможным лечением кокаиновой зависимости. В исследовании на кокаинозависимых крысах инактивация черной субстанции путем имплантации канюли значительно снизился рецидив кокаиновой зависимости.[43]
Леводопа
Черная субстанция является мишенью химических терапевтических средств для лечения болезни Паркинсона. Леводопа (обычно называемый L-DOPA), предшественник дофамина, является наиболее часто назначаемым лекарством от болезни Паркинсона, несмотря на разногласия относительно нейротоксичности дофамина и L-DOPA.[44] Препарат особенно эффективен при лечении пациентов на ранних стадиях болезни Паркинсона, хотя со временем он теряет свою эффективность.[45] Леводопа может пересечь гематоэнцефалический барьер и увеличивает уровень дофамина в черной субстанции, тем самым облегчая симптомы болезни Паркинсона. Недостатком лечения леводопой является то, что оно лечит симптомы болезни Паркинсона (низкий уровень дофамина), а не причину (гибель дофаминергических нейронов в черной субстанции).
MPTP
MPTP, представляет собой нейротоксин, специфичный для дофаминергических клеток головного мозга, особенно в черной субстанции. МФТП оказался в центре внимания в 1982 году, когда у потребителей героина в Калифорнии появились симптомы болезни Паркинсона после употребления. MPPP загрязнены МФТП. Пациенты, которые были жесткими и почти полностью неподвижными, ответили на лечение леводопой. О ремиссии симптомов, подобных болезни Паркинсона, не сообщалось, что свидетельствует о необратимой гибели дофаминергических нейронов.[46] Предлагаемый механизм МФТП предполагает нарушение митохондриальный функции, в том числе нарушение метаболизм и создание свободные радикалы.[47]
Вскоре после этого МФТП был протестирован на животных моделях на предмет его эффективности в индукции болезни Паркинсона (с успехом). MPTP вызывал акинезию, ригидность и тремор у приматов, и было обнаружено, что его нейротоксичность очень специфична для компактной части черной субстанции.[48] У других животных, таких как грызуны, индукция болезни Паркинсона с помощью МРТР является неполной или требует гораздо более высоких и частых доз, чем у приматов. Сегодня MPTP остается наиболее популярным методом индукции болезни Паркинсона у животные модели.[47][49]
История
Черная субстанция была открыта в 1784 г. Феликс Вик-д'Азир,[50] и Самуэль Томас фон Зёммерринг упомянул эту структуру в 1791 году.[51] Различение между pars reticulata и компактной черной субстанцией было впервые предложено Сано в 1910 году.[52] В 1963 г. Олег Горныкевич на основании своих наблюдений пришел к выводу, что «потеря клеток в черной субстанции (пациентов с болезнью Паркинсона) вполне может быть причиной дефицита дофамина в полосатом теле».[53]
Дополнительные изображения

Дофамин и серотонин

Деградация черной субстанции, связанная с болезнью Паркинсона.

Горизонтальный срез МРТ (взвешенный по T1) с выделением, указывающим на расположение черной субстанции.

Улучшенная МРТ нейромеланина с цветными изображениями (RGB), показывающими компактную часть черной субстанции

Микрофильмирование





Рекомендации
- ^ Рабей Дж. М., Хефти Ф (1990). «Синтез нейромеланина в черной субстанции крысы и человека». Журнал нейронной передачи. Отделение болезни Паркинсона и деменции. 2 (1): 1–14. Дои:10.1007 / BF02251241. PMID 2357268.
- ^ Kim SJ, Sung JY, Um JW, Hattori N, Mizuno Y, Tanaka K, Paik SR, Kim J, Chung KC (октябрь 2003 г.). «Паркин расщепляет внутриклеточные включения альфа-синуклеина посредством активации кальпаина». Журнал биологической химии. 278 (43): 41890–9. Дои:10.1074 / jbc.M306017200. PMID 12917442.
- ^ Bolam, J. P .; Браун, М. Т. С .; Moss, J .; Мэджилл, П. Дж. (1 января 2009 г.), Сквайр, Ларри Р. (ред.), «Базальные ганглии: внутренняя организация», Энциклопедия неврологии, Oxford: Academic Press, стр. 97–104, Дои:10.1016 / b978-008045046-9.01294-8, ISBN 978-0-08-045046-9, получено 7 сентября 2020
- ^ Кита Х, Джегер Д (2016). «Организация Глобус Паллидус». Справочник по структуре и функциям базальных ганглиев, второе издание. Справочник по поведенческой неврологии. 24. С. 259–276. Дои:10.1016 / B978-0-12-802206-1.00013-1. ISBN 9780128022061.
- ^ Наута Х.Дж., Коул М. (июль 1978 г.). «Эфферентные проекции субталамического ядра: авторадиографическое исследование на обезьянах и кошках». Журнал сравнительной неврологии. 180 (1): 1–16. Дои:10.1002 / cne.901800102. PMID 418083.[нужен лучший источник ]
- ^ Карпентер МБ, Накано К., Ким Р. (февраль 1976 г.). «Нигроталамические проекции у обезьяны, продемонстрированные авторадиографической техникой». Журнал сравнительной неврологии. 165 (4): 401–15. Дои:10.1002 / cne.901650402. PMID 57125.
- ^ Дениау Дж. М., Китай СТ, Донохью Дж. П., Грофова И. (1982). «Нейрональные взаимодействия в сетчатой части черной субстанции через коллатерали аксонов проекционных нейронов. Электрофизиологическое и морфологическое исследование». Экспериментальное исследование мозга. 47 (1): 105–13. Дои:10.1007 / BF00235891. PMID 6288427.
- ^ Никола СМ, Surmeier J, Маленка RC (2000). «Дофаминергическая модуляция возбудимости нейронов в полосатом теле и прилежащем ядре». Ежегодный обзор нейробиологии. 23: 185–215. Дои:10.1146 / annurev.neuro.23.1.185. PMID 10845063.
- ^ Гернерт М., Федровиц М., Влаз П., Лёшер В. (ноябрь 2004 г.). «Субрегиональные изменения скорости разряда, паттерна и лекарственной чувствительности предполагаемых ГАМКергических нигральных нейронов в киндлинг-модели эпилепсии». Европейский журнал нейробиологии. 20 (9): 2377–86. Дои:10.1111 / j.1460-9568.2004.03699.x. PMID 15525279.
- ^ Сато М., Хикосака О. (март 2002 г.). "Роль приматов черной субстанции pars reticulata в ориентированном на вознаграждение саккадическом движении глаз". Журнал неврологии. 22 (6): 2363–73. Дои:10.1523 / JNEUROSCI.22-06-02363.2002. ЧВК 6758246. PMID 11896175.
- ^ Зар Н.М., Мартин Л.П., Ващак Б.Л. (ноябрь 2004 г.). «Повреждения субталамического ядра изменяют базальный уровень, а агонист дофамина стимулирует электрофизиологический выход базальных ганглиев крысы». Синапс. 54 (2): 119–28. Дои:10.1002 / syn.20064. PMID 15352137.
- ^ Хикосака О., Вюрц Р.Х. (май 1983 г.). «Зрительные и глазодвигательные функции обезьяны nigra pars reticulata. III. Зрительные и саккадные реакции, обусловленные памятью». Журнал нейрофизиологии. 49 (5): 1268–84. Дои:10.1152 / jn.1983.49.5.1268. PMID 6864250.
- ^ Tseng KY, Riquelme LA, Belforte JE, Pazo JH, Murer MG (январь 2000 г.). «Черная субстанция pars reticulata единиц у крыс с поражением 6-гидроксидофамином: ответы на стимуляцию дофаминового рецептора D2 в полосатом теле и субталамические поражения». Европейский журнал нейробиологии. 12 (1): 247–56. Дои:10.1046 / j.1460-9568.2000.00910.x. PMID 10651879.
- ^ Deransart C, Hellwig B, Heupel-Reuter M, Léger JF, Heck D, Lücking CH (декабрь 2003 г.). "Одноэлементный анализ нейронов pars reticulata черной субстанции у свободно ведущих крыс с генетической абсансной эпилепсией". Эпилепсия. 44 (12): 1513–20. Дои:10.1111 / j.0013-9580.2003.26603.x. PMID 14636321.
- ^ Ходж Г.К., Мясник Л.Л. (август 1980 г.). «Компактная часть черной субстанции модулирует двигательную активность, но не играет важной роли в регулировании потребления пищи и воды». Архив фармакологии Наунин-Шмидеберг. 313 (1): 51–67. Дои:10.1007 / BF00505805. PMID 7207636.
- ^ Pioli EY, Meissner W, Sohr R, Gross CE, Bezard E, Bioulac BH (июнь 2008 г.). «Дифференциальные поведенческие эффекты частичных двусторонних поражений вентральной тегментальной области или компактной части черной субстанции у крыс». Неврология. 153 (4): 1213–24. Дои:10.1016 / j.neuroscience.2008.01.084. PMID 18455318.
- ^ а б Юнгберг Т., Апичелла П., Шульц В. (январь 1992 г.). «Ответы дофаминовых нейронов обезьяны во время обучения поведенческим реакциям». Журнал нейрофизиологии. 67 (1): 145–63. Дои:10.1152 / ян.1992.67.1.145. PMID 1552316.
- ^ Да Кунья С., Сильва М.Х., Витцикоски С., Витцикоски Е.С., Ферро М.М., Кузмин И., Кантерас Н.С. (декабрь 2006 г.). «Стратегия обучения места у крыс с поражением компактной части черной субстанции». Поведенческая неврология. 120 (6): 1279–84. Дои:10.1037/0735-7044.120.6.1279. PMID 17201473.
- ^ Да Кунья С., Витцикоски С., Витцикоски Е.С., Миёши Е., Ферро М.М., Ансельмо-Франси Д.А., Кантерас Н.С. (май 2003 г.). «Доказательства того, что компактная часть черной субстанции является важным компонентом системы памяти, независимой от системы памяти гиппокампа». Нейробиология обучения и памяти. 79 (3): 236–42. Дои:10.1016 / S1074-7427 (03) 00008-X. PMID 12676522.
- ^ Matell MS, Meck WH (январь 2000 г.). «Нейропсихологические механизмы интервального временного поведения». BioEssays. 22 (1): 94–103. Дои:10.1002 / (SICI) 1521-1878 (200001) 22: 1 <94 :: AID-BIES14> 3.0.CO; 2-E. PMID 10649295.
- ^ Лима М.М., Андерсен М.Л., Рексидлер А.Б., Витал М.А., Туфик С (июнь 2007 г.). Броснан С (ред.). «Роль компактной части черной субстанции в регуляции режима сна у крыс». PLOS One. 2 (6): e513. Bibcode:2007PLoSO ... 2..513L. Дои:10.1371 / journal.pone.0000513. ЧВК 1876809. PMID 17551593.

- ^ Дзираса К., Рибейро С., Коста Р, Сантос Л. М., Лин С. К., Гросмарк А., Сотникова Т. Д., Гайнетдинов Р. Р., Карон М. Г., Николелис М. А. (октябрь 2006 г.). «Дофаминергический контроль состояний сна и бодрствования». Журнал неврологии. 26 (41): 10577–89. Дои:10.1523 / JNEUROSCI.1767-06.2006. ЧВК 6674686. PMID 17035544.
- ^ Янкович Дж. (Апрель 2008 г.). «Болезнь Паркинсона: клиника и диагностика». Журнал неврологии, нейрохирургии и психиатрии. 79 (4): 368–76. Дои:10.1136 / jnnp.2007.131045. PMID 18344392.
- ^ Адлер СН (2005). «Немоторные осложнения при болезни Паркинсона». Двигательные расстройства. 20 Дополнение 11: S23-9. Дои:10.1002 / mds.20460. PMID 15822106.
- ^ Доусон TM, Доусон В.Л. (октябрь 2003 г.). «Молекулярные пути нейродегенерации при болезни Паркинсона». Наука. 302 (5646): 819–22. Bibcode:2003Наука ... 302..819D. Дои:10.1126 / science.1087753. PMID 14593166.
- ^ Лян С.Л., Синтон К.М., Сонсалла П.К., округ Колумбия, Германия (декабрь 1996 г.). «Дофаминергические нейроны среднего мозга у мышей, которые содержат кальбиндин-D28k, демонстрируют пониженную уязвимость к нейродегенерации, вызванной МРТР». Нейродегенерация. 5 (4): 313–8. Дои:10.1006 / нейр.1996.0042. PMID 9117542.
- ^ а б Опрос. Йоланд Смит, доктор философии[требуется проверка ]
- ^ Менке Р.А., Джбабди С., Миллер К.Л., Мэтьюз П.М., Зарей М. (октябрь 2010 г.). «Сегментация на основе подключения черной субстанции у человека и ее последствия при болезни Паркинсона». NeuroImage. 52 (4): 1175–80. Дои:10.1016 / j.neuroimage.2010.05.086. PMID 20677376.
- ^ Трист Б.Г., Заяц DJ, Дабл КЛ. Окислительный стресс при старении черной субстанции и этиология болезни Паркинсона. Ячейка старения. 2019 Декабрь; 18 (6): e13031. DOI: 10.1111 / acel.13031. Epub 2019 20 августа. Обзор. PMID 31432604
- ^ а б Schaser AJ, Osterberg VR, Dent SE, Stackhouse TL, Wakeham CM, Boutros SW, Weston LJ, Owen N, Weissman TA, Luna E, Raber J, Luk KC, McCullough AK, Woltjer RL, Unni VK. Альфа-синуклеин - это ДНК-связывающий белок, который модулирует восстановление ДНК с последствиями для заболеваний с тельцами Леви. Sci Rep.29 июля 2019; 9 (1): 10919. DOI: 10.1038 / s41598-019-47227-z. PMID 31358782
- ^ ван Россум Дж (1967). «Значение блокады дофаминовых рецепторов для действия нейролептических препаратов». В Brill H, Cole J, Deniker P, Hippius H, Bradley PB (ред.). Нейропсихофармакология, Труды Пятой международной коллегии Neuropsychopharmacologicum. С. 321–9. OCLC 458719.
- ^ Коломеец Н.С., Уранова Н.А. (1999). «Синаптические контакты при шизофрении: исследования с использованием иммуноцитохимической идентификации дофаминергических нейронов». Неврология и поведенческая физиология. 29 (2): 217–21. Дои:10.1007 / BF02465329. PMID 10432512.
- ^ Кумамото Н., Мацузаки С., Иноуэ К., Хаттори Т., Симидзу С., Хашимото Р., Яматодани А., Катаяма Т., Тохьяма М. (июнь 2006 г.). «Гиперактивация дофаминергической системы среднего мозга при шизофрении может быть связана с подавлением дисбиндина». Сообщения о биохимических и биофизических исследованиях. 345 (2): 904–9. Дои:10.1016 / j.bbrc.2006.04.163. PMID 16701550.
- ^ Сибата Е., Сасаки М., Тохьяма К., Оцука К., Эндох Дж., Тераяма Ю., Сакаи А. (сентябрь 2008 г.). «Использование нейромеланин-чувствительной МРТ для различения пациентов с шизофренией и депрессией и здоровых людей на основе изменений сигнала в черной субстанции и голубом пятне». Биологическая психиатрия. 64 (5): 401–6. Дои:10.1016 / j.biopsych.2008.03.021. PMID 18452894.
- ^ а б c d е Миллер GM (январь 2011 г.). «Возникающая роль следового аминосвязанного рецептора 1 в функциональной регуляции переносчиков моноаминов и дофаминергической активности». Журнал нейрохимии. 116 (2): 164–76. Дои:10.1111 / j.1471-4159.2010.07109.x. ЧВК 3005101. PMID 21073468.
- ^ «Амфетамин». DrugBank. Университет Альберты. 8 февраля 2013 г.. Получено 13 октября 2013.
- ^ а б c Эйден Л. Е., Вэйхэ Э (январь 2011 г.). «VMAT2: динамический регулятор моноаминергической нейрональной функции мозга, взаимодействующий с наркотиками». Летопись Нью-Йоркской академии наук. 1216 (1): 86–98. Bibcode:2011НЯСА1216 ... 86Е. Дои:10.1111 / j.1749-6632.2010.05906.x. ЧВК 4183197. PMID 21272013.
- ^ Магуайр Дж. Дж., Паркер В. А., Форд С. М., Боннер Т. И., Нойбиг Р. Р., Давенпорт А. П. (март 2009 г.). "Международный союз фармакологии. LXXII. Рекомендации по номенклатуре следовых аминовых рецепторов". Фармакологические обзоры. 61 (1): 1–8. Дои:10.1124 / пр.109.001107. ЧВК 2830119. PMID 19325074.
- ^ Heikkila RE, Cabbat FS, Duvoisin RC (1979). «Двигательная активность и вращательное поведение после аналогов кокаина: корреляция с блокадой захвата дофамина». Коммуникации в психофармакологии. 3 (5): 285–90. PMID 575770.
- ^ Лакоски Дж. М., Галлоуэй депутат, Белый FJ (1991). Кокаин. Телфорд Пресс. ISBN 978-0-8493-8813-2.
- ^ Лейси М.Г., Меркури Н.Б., Северное РА (апрель 1990 г.). «Действие кокаина на дофаминергические нейроны крысы in vitro». Британский журнал фармакологии. 99 (4): 731–5. Дои:10.1111 / j.1476-5381.1990.tb12998.x. ЧВК 1917549. PMID 2361170.
- ^ а б Ся Й, Гебель Д. Д., Капатос Г., Бэннон М. Дж. (Сентябрь 1992 г.). «Количественное определение мРНК переносчика дофамина крысы: эффекты лечения кокаином и отмены». Журнал нейрохимии. 59 (3): 1179–82. Дои:10.1111 / j.1471-4159.1992.tb08365.x. PMID 1494906.
- ^ См. RE, Elliott JC, Feltenstein MW (октябрь 2007 г.). «Роль дорсальных и вентральных стриатальных путей в поиске кокаина после длительного воздержания у крыс». Психофармакология. 194 (3): 321–31. Дои:10.1007 / s00213-007-0850-8. PMID 17589830.
- ^ Ченг Н., Маэда Т., Куме Т., Канеко С., Кочияма Х., Акаике А., Гошима Ю., Мису Ю. (декабрь 1996 г.). «Дифференциальная нейротоксичность, вызванная L-DOPA и дофамином в культивируемых нейронах полосатого тела». Исследование мозга. 743 (1–2): 278–83. Дои:10.1016 / S0006-8993 (96) 01056-6. PMID 9017256.
- ^ Rascol O, Payoux P, Ory F, Ferreira JJ, Brefel-Courbon C, Montastruc JL (2003). «Ограничения современной терапии болезни Паркинсона». Анналы неврологии. 53 Дополнение 3: S3–12, обсуждение S12–5. Дои:10.1002 / ana.10513. PMID 12666094.
- ^ Лэнгстон Дж. В., Баллард П., Тетруд Дж. В., Ирвин I (февраль 1983 г.). «Хронический паркинсонизм у людей, вызванный продуктом синтеза аналога меперидина». Наука. 219 (4587): 979–80. Bibcode:1983Sci ... 219..979L. Дои:10.1126 / science.6823561. PMID 6823561.
- ^ а б Шмидт Н, Фергер Б (2001). «Нейрохимические данные в модели болезни Паркинсона MPTP». Журнал нейронной передачи. 108 (11): 1263–82. Дои:10.1007 / s007020100004. PMID 11768626.
- ^ Лэнгстон Дж. В., Форно Л. С., Реберт С. С., Ирвин I (февраль 1984 г.). «Избирательная нигральная токсичность после системного введения 1-метил-4-фенил-1,2,5,6-тетрагидропирина (МРТР) у беличьей обезьяны». Исследование мозга. 292 (2): 390–4. Дои:10.1016/0006-8993(84)90777-7. PMID 6607092.
- ^ Бланше П.Дж., Калон Ф., Мориссетт М., Хадж Тахар А., Беланже Н., Самади П., Грондин Р., Грегуар Л., Мельцер Л., Ди Паоло Т., Бедар П.Дж. (июль 2004 г.). «Актуальность модели приматов MPTP в изучении механизмов прайминга дискинезии». Паркинсонизм и связанные с ним расстройства. 10 (5): 297–304. Дои:10.1016 / j.parkreldis.2004.02.011. PMID 15196509.
- ^ Таббс Р.С., Лукас М., Шоджа М.М., Мортазави М.М., Коэн-Гадол А.А. (июль 2011 г.). «Феликс Вик д'Азир (1746–1794): один из первых основателей нейроанатомии и королевский французский врач». Нервная система ребенка. 27 (7): 1031–4. Дои:10.1007 / s00381-011-1424-y. PMID 21445631.
- ^ Свонсон Л.В. (2014). Нейроанатомическая терминология: лексикон классических истоков и исторических основ. Англия: Издательство Оксфордского университета. ISBN 9780195340624.
- ^ Сано Т. (1910). "Beitrag zur vergleichenden Anatomie der Substantia nigra, des Corpus Luysii und der Zona incerta". MSCHR Psychiat Neurol. 28 (1): 26–34. Дои:10.1159/000209678.
- ^ Хорныкевич, О. (2006). «Открытие дофаминовой недостаточности в паркинсоническом мозге». В Riederer, P .; Reichmann, H .; Youdim, M. B. H .; Герлах, М. (ред.). Болезнь Паркинсона и связанные с ней заболевания. Журнал нейронной передачи. Дополнение. Springer Вена. С. 9–15. Дои:10.1007/978-3-211-45295-0_3. ISBN 978-3-211-28927-3. PMID 17017502.
внешняя ссылка
- Изображения окрашенных срезов головного мозга, которые включают "черную субстанцию" на Проект BrainMaps
- Схема и объяснение кортико-базальных путей
| Авторитетный контроль |
|---|