Переносчик дофамина - Dopamine transporter




В переносчик дофамина (также активный переносчик дофамина, DAT, SLC6A3) представляет собой трансмембранный белок, который перекачивает нейротрансмиттер дофамин вне синаптическая щель назад в цитозоль. В цитозоле другие переносчики секвестрируют дофамин в пузырьки для хранения и последующего выпуска. Обратный захват дофамина через DAT обеспечивает основной механизм, с помощью которого дофамин очищается от синапсы, хотя может быть исключение в префронтальной коре, где данные указывают на возможно большую роль переносчик норэпинефрина.[5]
DAT участвует в ряде заболеваний, связанных с дофамином, включая Синдром дефицита внимания и гиперактивности, биполярное расстройство, клиническая депрессия, алкоголизм, и расстройство, связанное с употреблением психоактивных веществ. В ген который кодирует белок DAT, расположен на хромосоме 5 человека, состоит из 15 кодирующих экзоны, и составляет примерно 64 kbp длинный. Доказательства связи между DAT и расстройствами, связанными с дофамином, получены из генетический полиморфизм, известный как ВНТР, в гене DAT (DAT1 ), который влияет на количество экспрессируемого белка.[6]
Функция
DAT - это интегральный мембранный белок который удаляет дофамин из синаптическая щель и откладывает его в окружающие клетки, тем самым прекращая сигнал нейротрансмиттера. Дофамин лежит в основе нескольких аспектов познания, включая вознаграждение, а DAT способствует регулированию этого сигнала.[7]
Механизм
DAT - это сторонник который перемещает дофамин через клеточную мембрану, связывая движение с энергетически выгодным движением ионов натрия, перемещающихся от высокой к низкой концентрации в клетку. Функция DAT требует последовательной привязки и совместный транспорт из двух Na+ ионы и один Cl− ион с дофаминовым субстратом. Движущей силой DAT-опосредованного обратного захвата дофамина является градиент концентрации ионов, создаваемый плазматической мембраной. Na+/ К+ АТФаза.[8]
В наиболее распространенной модели для транспортер моноаминов функции, ионы натрия должны связываться с внеклеточным доменом переносчика, прежде чем дофамин сможет связываться. Как только дофамин связывается, белок претерпевает конформационные изменения, которые позволяют как натрию, так и дофамину расщепляться на внутриклеточной стороне мембраны.[9]
Исследования с использованием электрофизиология и радиоактивно меченый дофамин подтвердили, что переносчик дофамина подобен другим переносчикам моноаминов в том, что одна молекула нейротрансмиттера может переноситься через мембрану с одним или двумя ионами натрия. Ионы хлорида также необходимы для предотвращения накопления положительного заряда. Эти исследования также показали, что скорость и направление переноса полностью зависят от градиента натрия.[10]
Из-за плотного соединения мембранный потенциал и градиент натрия, вызванные активностью изменения полярности мембраны могут резко влиять на скорость переноса. Кроме того, переносчик может способствовать высвобождению дофамина при деполяризации нейрона.[10]
DAT – Cav связь
Предварительные данные свидетельствуют о том, что переносчик дофамина соединяется с L-типом потенциалзависимые кальциевые каналы (особенно Cav1.2 и Cav1.3 ), которые экспрессируются практически во всех дофаминовых нейронах.[11] В результате DAT – Cav соединение, подложки DAT, которые производят деполяризующий токи через транспортер способны открывать кальциевые каналы, которые связаны с транспортером, что приводит к притоку кальция в дофаминовые нейроны.[11] Считается, что приток кальция вызывает CAMKII -опосредованный фосфорилирование переносчика дофамина как последующий эффект;[11] поскольку фосфорилирование DAT с помощью CAMKII приводит к оттоку дофамина in vivo активация кальциевых каналов, связанных с транспортером, является потенциальным механизмом, с помощью которого определенные лекарства (например, амфетамин) запускают высвобождение нейромедиатора.[11]
Белковая структура
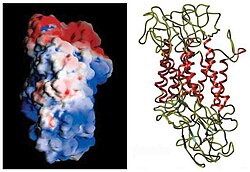
Первоначальное определение топологии мембраны DAT было основано на анализ гидрофобной последовательности и сходство последовательностей с транспортером ГАМК. Эти методы предсказали двенадцать трансмембранные домены (TMD) с большой внеклеточной петлей между третьей и четвертой TMD.[12] Используется дополнительная характеристика этого белка протеазы, которые переваривают белки на более мелкие фрагменты, и гликозилирование, которое происходит только во внеклеточных петлях, и в значительной степени подтвердило первоначальные предсказания топологии мембраны.[13] Точная структура Drosophila melanogaster переносчик дофамина (dDAT) был выяснен в 2013 г. Рентгеновская кристаллография.[14]
Расположение и распространение
Фармакодинамика амфетамина в дофаминовом нейроне |

Региональное распределение DAT было обнаружено в областях мозга с установленной дофаминергической схемой, включая: нигростриальный, мезолимбический, и мезокортикальный пути.[22] В ядра , составляющие эти пути, имеют различные паттерны выражения. Паттерны экспрессии генов у взрослых мышей показывают высокую экспрессию в черная субстанция pars compacta.[23]
DAT в мезокортикальный путь, меченный радиоактивными антителами, было обнаружено, что он обогащен дендритами и клеточными телами нейронов в черная субстанция pars compacta и вентральная тегментальная область. Этот паттерн имеет смысл для белка, который регулирует уровень дофамина в синапсе.
Окрашивание в полосатое тело и прилежащее ядро мезолимбического пути был плотным и неоднородным. В полосатом теле DAT локализован в плазматической мембране окончаний аксонов. Двойной иммуноцитохимия продемонстрировали совместную локализацию DAT с двумя другими маркерами нигростриальный терминалы, тирозингидроксилаза и Дофаминовые рецепторы D2. Таким образом было продемонстрировано, что последний ауторецептор на клетки, которые выделяют дофамин. TAAR1 представляет собой пресинаптический внутриклеточный рецептор, который также колокализованный с DAT и который при активации имеет эффект, противоположный авторецептору D2;[15][24] то есть он интернализует переносчики дофамина и вызывает отток через обратную функцию переносчика через PKA и PKC сигнализация.
Удивительно, но DAT не был идентифицирован ни в одной синаптической активной зоне. Эти результаты предполагают, что обратный захват дофамина полосатым телом может происходить вне синаптических специализаций, когда дофамин диффундирует из синаптической щели.
в черная субстанция, DAT локализован в аксональных и дендритных (то есть пре- и постсинаптических) плазматические мембраны.[25]
В рамках перикария из pars compacta нейроны, DAT был локализован в основном в шероховатой и гладкой эндоплазматической сети, аппарат Гольджи и мультивезикулярные тела, идентифицирующие вероятные места синтеза, модификации, транспорта и деградации.[26]
Генетика и регуляция
В ген для DAT, известный как DAT1, расположен на хромосоме 5p15.[6] Белковая кодирующая область гена имеет длину более 64 т.п.н. и включает 15 кодирующих сегментов или экзоны.[27] Этот ген имеет переменный номер тандемный повтор (VNTR) на 3 ’конце (rs28363170 ) и еще один в интрон 8 регион.[28] Было показано, что различия в VNTR влияют на базальный уровень экспрессии переносчика; следовательно, исследователи искали ассоциации с расстройствами, связанными с дофамином.[29]
Nurr1, а ядерный рецептор который регулирует многие гены, связанные с дофамином, может связывать промоутер области этого гена и вызывают экспрессию.[30] Этот промотор также может быть целью фактор транскрипции Сп-1.
В то время как факторы транскрипции контролируют, какие клетки экспрессируют DAT, функциональная регуляция этого белка в значительной степени осуществляется за счет киназы. MAPK,[31] CAMKII,[20][21] PKA,[15] и PKC[21][32] может модулировать скорость, с которой переносчик перемещает дофамин, или вызывать интернализацию DAT. Совместная локализация TAAR1 является важным регулятором переносчика дофамина, который при активации фосфорилирует DAT через протеинкиназа А (PKA) и протеинкиназа C (PKC) сигнализация.[15][33] Фосфорилирование любой протеинкиназой может приводить к DAT интернализация (неконкурентоспособный ингибирование обратного захвата), но PKC-опосредованный только фосфорилирование индуцирует функцию обратного переносчика (дофамин излияние ).[15][34] Ауторецепторы дофамина также регулируют DAT, прямо противодействуя эффекту активации TAAR1.[15]
Переносчик дофамина человека (hDAT) содержит высокая близость внеклеточный цинк сайт привязки который при связывании цинка ингибирует дофамин обратный захват и усиливает амфетамин -индуцированный отток дофамина in vitro.[35][36][37] В отличие от человека переносчик серотонина (hSERT) и человек переносчик норэпинефрина (hNET) не содержат сайтов связывания цинка.[37] Добавка цинка может уменьшить минимум эффективная доза амфетамина, когда он используется для лечения Синдром дефицита внимания и гиперактивности.[38]
Биологическая роль и нарушения
Скорость, с которой DAT удаляет дофамин из синапса, может иметь огромное влияние на количество дофамина в клетке. Об этом лучше всего свидетельствуют тяжелые когнитивные нарушения, двигательные аномалии и гиперактивность мышей без переносчиков дофамина.[39] Эти характеристики имеют поразительное сходство с симптомами СДВГ.
Отличия в функционале ВНТР были определены как факторы риска биполярного расстройства[40] и СДВГ.[41] Появились данные, свидетельствующие о том, что существует также связь с более сильным снятие симптомы от алкоголизм, хотя это вызывает споры.[42][43] An аллель гена DAT с нормальным уровнем белка связан с поведением некурящих и легкостью отказа от курения.[44] Кроме того, подростки мужского пола, особенно из семей с высоким уровнем риска (в тех семьях, где мать не вовлечена в процесс и отсутствует материнская привязанность), которые несут 10-аллельный повтор VNTR, демонстрируют статистически значимое сродство с антисоциальными сверстниками.[45][46]
Повышенная активность DAT связана с несколькими различными расстройствами, включая: клиническая депрессия.[47]
Было показано, что мутации в DAT вызывают синдром дефицита переносчика дофамина, аутосомно-рецессивный расстройство движения характеризуется прогрессирующим ухудшением дистония и паркинсонизм.[48]
Фармакология
Переносчик дофамина является целью субстраты, высвобождающие дофамин, ингибиторы транспорта и аллостерические модуляторы.[49][50]
Кокаин блокирует DAT, связываясь непосредственно с транспортером и снижая скорость транспорта.[12] Напротив, амфетамин попадает в пресинаптический нейрон непосредственно через нейрональную мембрану или через DAT, конкурируя за обратный захват с дофамином. Оказавшись внутри, он связывается с TAAR1 или попадает в синаптические везикулы через VMAT2. Когда амфетамин связывается с TAAR1, он снижает скорость возбуждения постсинаптического нейрона и запускает протеинкиназа А и протеинкиназа C передача сигналов, что приводит к фосфорилированию DAT. Затем фосфорилированный DAT либо действует в обратном порядке, либо уходит в пресинаптический нейрон и прекращает транспорт. Когда амфетамин попадает в синаптические пузырьки через VMAT2, дофамин высвобождается в цитозоль.[15][16] Амфетамин также вызывает отток дофамина через второй TAAR1-независимый механизм, включающий CAMKIIα -опосредованное фосфорилирование переносчика, которое предположительно возникает в результате активации DAT-связанные кальциевые каналы L-типа амфетамином.[11]
Считается, что дофаминергические механизмы каждого препарата лежат в основе приятных ощущений, вызываемых этими веществами.[7]
Взаимодействия
Переносчик дофамина может взаимодействовать с:
Помимо этих врожденных белок-белковых взаимодействий, недавние исследования показали, что вирусные белки, такие как ВИЧ-1 Tat белок взаимодействует с DAT[55][56] и это связывание может изменить гомеостаз дофамина у ВИЧ-инфицированных, что является фактором, способствующим развитию ВИЧ-ассоциированные нейрокогнитивные расстройства.[57]
Смотрите также
Рекомендации
- ^ а б c ENSG00000276996 GRCh38: Общий выпуск 89: ENSG00000142319, ENSG00000276996 - Ансамбль, Май 2017
- ^ а б c GRCm38: выпуск Ensembl 89: ENSMUSG00000021609 - Ансамбль, Май 2017
- ^ "Справочник человека по PubMed:". Национальный центр биотехнологической информации, Национальная медицинская библиотека США.
- ^ "Ссылка на Mouse PubMed:". Национальный центр биотехнологической информации, Национальная медицинская библиотека США.
- ^ Карбони Э., Танда Г.Л., фрау Р., Ди Кьяра Дж. (Сентябрь 1990 г.). «Блокада носителя норадреналина увеличивает внеклеточную концентрацию дофамина в префронтальной коре: доказательства того, что дофамин поглощается in vivo норадренергическими окончаниями». Журнал нейрохимии. 55 (3): 1067–70. Дои:10.1111 / j.1471-4159.1990.tb04599.x. PMID 2117046. S2CID 23682303.
- ^ а б Ванденберг Д. Д., Персико А. М., Хокинс А. Л., Гриффин К. А., Ли Икс, Джабс Е. В., Уль Г. Р. (декабрь 1992 г.). «Ген транспортера дофамина человека (DAT1) отображается на хромосоме 5p15.3 и отображает VNTR». Геномика. 14 (4): 1104–6. Дои:10.1016 / S0888-7543 (05) 80138-7. PMID 1478653.
- ^ а б Шульц В. (июль 1998 г.). «Прогнозирующий сигнал вознаграждения дофаминовых нейронов». Журнал нейрофизиологии. 80 (1): 1–27. Дои:10.1152 / jn.1998.80.1.1. PMID 9658025.
- ^ Торрес Г.Э., Гайнетдинов Р.Р., Карон М.Г. (январь 2003 г.). «Плазматические мембранные переносчики моноаминов: структура, регуляция и функции». Обзоры природы. Неврология. 4 (1): 13–25. Дои:10.1038 / nrn1008. PMID 12511858. S2CID 21545649.
- ^ Сондерс М.С., Чжу С.Дж., Захнисер Н.Р., Кавано М.П., Амара С.Г. (февраль 1997 г.). «Множественные ионные проводимости переносчика дофамина человека: действие дофамина и психостимуляторов». Журнал неврологии. 17 (3): 960–74. Дои:10.1523 / JNEUROSCI.17-03-00960.1997. ЧВК 6573182. PMID 8994051.
- ^ а б Уиллер Д.Д., Эдвардс А.М., Чепмен Б.М., Ондо Дж. Г. (август 1993 г.). «Модель натриевой зависимости поглощения дофамина в синаптосомах полосатого тела крысы». Нейрохимические исследования. 18 (8): 927–36. Дои:10.1007 / BF00998279. PMID 8371835. S2CID 42196576.
- ^ а б c d е Кэмерон К.Н., Солис Э., Ручала И., Де Феличе Л.Дж., Элтит Дж.М. (ноябрь 2015 г.). «Амфетамин активирует кальциевые каналы посредством деполяризации, опосредованной переносчиком дофамина». Клеточный кальций. 58 (5): 457–66. Дои:10.1016 / j.ceca.2015.06.013. ЧВК 4631700. PMID 26162812.
Одним из интересных примеров является CaMKII, который был хорошо охарактеризован как эффектор потоков Ca2 + ниже по течению от каналов Ca2 + L-типа [21,22]. Интересно, что DAT является субстратом CaMKII, а фосфорилированный DAT способствует обратному транспорту дофамина [48,49], составляя возможный механизм, с помощью которого электрическая активность и каналы Ca2 + L-типа могут модулировать состояния DAT и высвобождение дофамина. ... Таким образом, наши результаты предполагают, что с фармакологической точки зрения S (+) AMPH более эффективен, чем DA, в отношении активации hDAT-опосредованных деполяризующих токов, что приводит к активации Ca2 + -каналов L-типа, а индуцированный S (+) AMPH ток составляет более тесно связаны, чем DA, чтобы открывать каналы Ca2 + L-типа.
- ^ а б Килти Дж. Э., Лоранг Д., Амара С. Г. (октябрь 1991 г.). «Клонирование и экспрессия крысиного переносчика дофамина, чувствительного к кокаину». Наука. 254 (5031): 578–9. Bibcode:1991Научный ... 254..578K. Дои:10.1126 / science.1948035. PMID 1948035.
- ^ Воан Р.А., Кухар М.Дж. (август 1996 г.). «Домены, связывающие лиганд переносчика дофамина. Структурные и функциональные свойства, выявленные ограниченным протеолизом». Журнал биологической химии. 271 (35): 21672–80. Дои:10.1074 / jbc.271.35.21672. PMID 8702957.
- ^ Penmatsa A, Wang KH, Gouaux E (ноябрь 2013 г.). «Рентгеновская структура переносчика дофамина проясняет антидепрессивный механизм». Природа. 503 (7474): 85–90. Bibcode:2013Натура.503 ... 85С. Дои:10.1038 / природа12533. ЧВК 3904663. PMID 24037379.
- ^ а б c d е ж грамм час я j k Миллер GM (январь 2011 г.). «Возникающая роль следового аминосвязанного рецептора 1 в функциональной регуляции переносчиков моноаминов и дофаминергической активности». Журнал нейрохимии. 116 (2): 164–76. Дои:10.1111 / j.1471-4159.2010.07109.x. ЧВК 3005101. PMID 21073468.
- ^ а б c Эйден Л. Е., Вэйхэ Э (январь 2011 г.). «VMAT2: динамический регулятор моноаминергической нейрональной функции мозга, взаимодействующий с наркотиками». Летопись Нью-Йоркской академии наук. 1216 (1): 86–98. Bibcode:2011НЯСА1216 ... 86Е. Дои:10.1111 / j.1749-6632.2010.05906.x. ЧВК 4183197. PMID 21272013.
- ^ Sulzer D, Cragg SJ, Rice ME (август 2016 г.). «Стриатальная нейротрансмиссия дофамина: регуляция высвобождения и захвата». Базальный ганглий. 6 (3): 123–148. Дои:10.1016 / j.baga.2016.02.001. ЧВК 4850498. PMID 27141430.
Несмотря на проблемы с определением pH синаптических везикул, градиент протонов через мембрану везикул имеет фундаментальное значение для его функции. Воздействие протонофоров на изолированные катехоламиновые везикулы снижает градиент pH и быстро перераспределяет медиатор изнутри наружу везикулы. ... Амфетамин и его производные, такие как метамфетамин, являются слабыми основными соединениями, которые являются единственным широко используемым классом лекарств, которые, как известно, вызывают высвобождение медиатора по неэкзоцитарному механизму. В качестве субстратов как для DAT, так и для VMAT амфетамины могут попадать в цитозоль, а затем секвестрироваться в везикулах, где они действуют, разрушая везикулярный градиент pH.
- ^ Ледонн А., Берретта Н., Даволи А., Риццо Г.Р., Бернарди Дж., Меркури Н.Б. (июль 2011 г.). «Электрофизиологические эффекты следовых аминов на дофаминергические нейроны мезэнцефала». Передний. Syst. Неврологи. 5: 56. Дои:10.3389 / fnsys.2011.00056. ЧВК 3131148. PMID 21772817.
Недавно появились три новых важных аспекта действия ТА: (а) ингибирование возбуждения из-за повышенного высвобождения дофамина; (b) снижение ингибирующих ответов, опосредованных рецепторами D2 и GABAB (возбуждающие эффекты из-за растормаживания); и (c) прямая опосредованная рецептором TA1 активация каналов GIRK, которая вызывает гиперполяризацию клеточной мембраны.
- ^ «ТААР1». GenAtlas. Парижский университет. 28 января 2012 г.. Получено 29 мая 2014.
• тонически активирует внутренне выпрямляющие K (+) каналы, что снижает базальную частоту возбуждения дофаминовых (DA) нейронов вентральной тегментальной области (VTA)
- ^ а б Андерхилл С.М., Уиллер Д.С., Ли М., Уоттс С.Д., Инграм С.Л., Амара С.Г. (июль 2014 г.). «Амфетамин модулирует возбуждающую нейротрансмиссию посредством эндоцитоза транспортера глутамата EAAT3 в дофаминовых нейронах». Нейрон. 83 (2): 404–416. Дои:10.1016 / j.neuron.2014.05.043. ЧВК 4159050. PMID 25033183.
AMPH также увеличивает внутриклеточный кальций (Gnegy et al., 2004), который связан с активацией кальмодулина / CamKII (Wei et al., 2007), а также модуляцией и перемещением DAT (Fog et al., 2006; Sakrikar et al., 2012). ).
- ^ а б c Воан Р.А., Фостер Д.Д. (сентябрь 2013 г.). «Механизмы регуляции переносчика дофамина в нормальных и болезненных состояниях». Тенденции в фармакологических науках. 34 (9): 489–96. Дои:10.1016 / j.tips.2013.07.005. ЧВК 3831354. PMID 23968642.
AMPH и METH также стимулируют отток DA, который считается ключевым элементом их аддиктивных свойств [80], хотя механизмы, по-видимому, не идентичны для каждого препарата [81]. Эти процессы являются PKCβ– и CaMK-зависимыми [72, 82], и мыши с нокаутом PKCβ обнаруживают снижение индуцированного AMPH оттока, что коррелирует со снижением индуцированной AMPH локомоции [72].
- ^ Цилиакс Б.Дж., Драш Г.В., Стейли Дж.К., Хабер С., Мобли С.Дж., Миллер Г.В., Муфсон Е.Дж., Маш, округ Колумбия, Леви А.И. (июнь 1999 г.). «Иммуноцитохимическая локализация переносчика дофамина в головном мозге человека». Журнал сравнительной неврологии. 409 (1): 38–56. Дои:10.1002 / (SICI) 1096-9861 (19990621) 409: 1 <38 :: AID-CNE4> 3.0.CO; 2-1. PMID 10363710.
- ^ Лю З., Янь С.Ф., Уокер Дж. Р., Цвингман Т. А., Цзян Т., Ли Дж., Чжоу Ю. (апрель 2007 г.). «Изучение функции генов на основе пространственной коэкспрессии в атласе мозга мышей с высоким разрешением». BMC Systems Biology. 1: 19. Дои:10.1186/1752-0509-1-19. ЧВК 1863433. PMID 17437647.
- ^ Магуайр Дж. Дж., Давенпорт А. П. (19 июля 2016 г.). «Рецептор следовых аминов: TA1 рецептор ". IUPHAR / BPS Руководство по ФАРМАКОЛОГИИ. Международный союз фундаментальной и клинической фармакологии. Получено 22 сентября 2016.
- ^ Ниренберг М.Дж., Воан Р.А., Уль Г.Р., Кухар М.Дж., Пикель В.М. (январь 1996 г.). «Переносчик дофамина локализован на дендритных и аксональных плазматических мембранах нигростриатных дофаминергических нейронов». Журнал неврологии. 16 (2): 436–47. Дои:10.1523 / JNEUROSCI.16-02-00436.1996. ЧВК 6578661. PMID 8551328.
- ^ Hersch SM, Yi H, Heilman CJ, Edwards RH, Levey AI (ноябрь 1997 г.). «Субклеточная локализация и молекулярная топология переносчика дофамина в полосатом теле и черной субстанции». Журнал сравнительной неврологии. 388 (2): 211–27. Дои:10.1002 / (SICI) 1096-9861 (19971117) 388: 2 <211 :: AID-CNE3> 3.0.CO; 2-4. PMID 9368838.
- ^ Кавараи Т., Каваками Х., Ямамура Й., Накамура С. (август 1997 г.). «Структура и организация гена, кодирующего переносчик дофамина человека». Ген. 195 (1): 11–8. Дои:10.1016 / S0378-1119 (97) 00131-5. PMID 9300814.
- ^ Сано А., Кондо К., Какимото Ю., Кондо И. (май 1993 г.). «Полиморфизм 40-нуклеотидных повторов в гене переносчика дофамина человека». Генетика человека. 91 (4): 405–6. Дои:10.1007 / BF00217369. PMID 8500798. S2CID 39416578.
- ^ Миллер GM, Мадрас BK (2002). «Полиморфизмы в 3'-нетранслируемой области генов переносчиков дофамина человека и обезьяны влияют на экспрессию репортерного гена». Молекулярная психиатрия. 7 (1): 44–55. Дои:10,1038 / sj / mp / 4000921. PMID 11803445.
- ^ Саккетти П., Митчелл Т.Р., Граннеман Дж. Г., Бэннон М.Дж. (март 2001 г.). «Nurr1 усиливает транскрипцию гена переносчика дофамина человека с помощью нового механизма». Журнал нейрохимии. 76 (5): 1565–72. Дои:10.1046 / j.1471-4159.2001.00181.x. PMID 11238740. S2CID 19410051.
- ^ Морон Дж. А., Захарова И., Феррер Дж. В., Меррилл Г. А., Хоуп Б., Лафер Э. М., Лин З. К., Ван Дж. Б., Явич Дж. А., Галли А., Шиппенберг Т. С. (сентябрь 2003 г.). «Активированная митогеном протеинкиназа регулирует поверхностную экспрессию переносчика дофамина и его транспортную способность». Журнал неврологии. 23 (24): 8480–8. Дои:10.1523 / JNEUROSCI.23-24-08480.2003. ЧВК 6740378. PMID 13679416.
- ^ Приступа З.Б., МакКонки Ф., Лю Ф., Ман Х.Й., Ли Ф.Дж., Ван Ю.Т., Низник HB (сентябрь 1998 г.). «Опосредованное протеинкиназой двунаправленное движение и функциональная регуляция переносчика дофамина человека». Синапс. 30 (1): 79–87. Дои:10.1002 / (SICI) 1098-2396 (199809) 30: 1 <79 :: AID-SYN10> 3.0.CO; 2-K. PMID 9704884.
- ^ Lindemann L, Ebeling M, Kratochwil NA, Bunzow JR, Grandy DK, Hoener MC (март 2005 г.). «Рецепторы, связанные со следовыми аминогруппами, образуют структурно и функционально отдельные подсемейства новых рецепторов, связанных с G-белком». Геномика. 85 (3): 372–85. Дои:10.1016 / j.ygeno.2004.11.010. PMID 15718104.
- ^ Магуайр Дж. Дж., Паркер В. А., Форд С. М., Боннер Т. И., Нойбиг Р. Р., Давенпорт А. П. (март 2009 г.). "Международный союз фармакологии. LXXII. Рекомендации по номенклатуре следовых аминовых рецепторов". Фармакологические обзоры. 61 (1): 1–8. Дои:10.1124 / пр.109.001107. ЧВК 2830119. PMID 19325074.
- ^ Краузе Дж (апрель 2008 г.). «ОФЭКТ и ПЭТ транспортера дофамина при синдроме дефицита внимания / гиперактивности». Экспертный обзор нейротерапии. 8 (4): 611–25. Дои:10.1586/14737175.8.4.611. PMID 18416663. S2CID 24589993.
Цинк связывается во внеклеточных сайтах DAT [103], выступая в качестве ингибитора DAT. В этом контексте представляют интерес контролируемые двойные слепые исследования на детях, которые показали положительное влияние цинка [добавок] на симптомы СДВГ [105,106]. Следует отметить, что в настоящее время [добавки] цинка не интегрированы ни в один алгоритм лечения СДВГ.
- ^ Зульцер Д. (февраль 2011 г.). «Как наркотики, вызывающие привыкание, нарушают пресинаптическую нейротрансмиссию дофамина». Нейрон. 69 (4): 628–49. Дои:10.1016 / j.neuron.2011.02.010. ЧВК 3065181. PMID 21338876.
Они не подтвердили предсказанную прямую взаимосвязь между поглощением и высвобождением, а скорее то, что некоторые соединения, включая AMPH, были лучше высвобождающими, чем субстратами для поглощения. Цинк, кроме того, стимулирует отток внутриклеточного [3H] DA, несмотря на его сопутствующее ингибирование поглощения (Scholze et al., 2002).
- ^ а б Scholze P, Nørregaard L, Singer EA, Freissmuth M, Gether U, Sitte HH (июнь 2002 г.). «Роль ионов цинка в обратном транспорте, опосредованном переносчиками моноаминов». Журнал биологической химии. 277 (24): 21505–13. Дои:10.1074 / jbc.M112265200. PMID 11940571.
Переносчик дофамина человека (hDAT) содержит эндогенный высокоаффинный Zn2+ сайт связывания с тремя координирующими остатками на внеклеточной поверхности (His193, His375 и Glu396). ... Хотя Zn2+ ингибирование захвата, Zn2+ облегчает высвобождение [3H] MPP +, индуцированное амфетамином, MPP + или K + -индуцированной деполяризацией, специфически на hDAT, но не на серотонине человека и переносчике норэпинефрина (hNET).
- ^ Scassellati C, Bonvicini C, Faraone SV, Gennarelli M (октябрь 2012 г.). «Биомаркеры и синдром дефицита внимания / гиперактивности: систематический обзор и метаанализы». Журнал Американской академии детской и подростковой психиатрии. 51 (10): 1003–1019.e20. Дои:10.1016 / j.jaac.2012.08.015. PMID 23021477.
Что касается добавок цинка, то в плацебо-контролируемом исследовании сообщалось, что дозы цинка до 30 мг / день были безопасными в течение как минимум 8 недель, но клинический эффект был неоднозначным, за исключением обнаружения снижения оптимальной дозы амфетамина на 37% с 30%. мг в день цинка.110
- ^ Гайнетдинов Р.Р., Ветсел В.К., Джонс С.Р., Левин Э.Д., Джабер М., Кэрон М.Г. (январь 1999 г.). «Роль серотонина в парадоксальном успокаивающем действии психостимуляторов на гиперактивность». Наука. 283 (5400): 397–401. Bibcode:1999Наука ... 283..397Г. Дои:10.1126 / science.283.5400.397. PMID 9888856. S2CID 9629915.
- ^ Гринвуд Т.А., Александр М., Кек П.Е., МакЭлрой С., Садовник А.Д., Ремик Р.А., Келсо-младший (март 2001 г.). «Доказательства неравновесия по сцеплению между переносчиком дофамина и биполярным расстройством». Американский журнал медицинской генетики. 105 (2): 145–51. Дои:10.1002 / 1096-8628 (2001) 9999: 9999 <:: AID-AJMG1161> 3.0.CO; 2-8. PMID 11304827.
- ^ Ян Б., Чан Р.С., Цзин Дж., Ли Т., Шам П., Чен Р.Й. (июнь 2007 г.). «Метаанализ исследований ассоциации между 10-повторным аллелем полиморфизма VNTR в 3'-UTR гена транспортера дофамина и синдромом дефицита внимания с гиперактивностью». Американский журнал медицинской генетики. Часть B, Психоневрологическая генетика. 144B (4): 541–50. Дои:10.1002 / ajmg.b.30453. PMID 17440978. S2CID 22881996.
- ^ Сандер Т., Хармс Х., Подшус Дж., Финк У., Никель Б., Рольфс А., Роммельспахер Х., Шмидт Л.Г. (февраль 1997 г.). «Аллельная ассоциация полиморфизма гена переносчика дофамина при алкогольной зависимости с припадками отмены или делирием». Биологическая психиатрия. 41 (3): 299–304. Дои:10.1016 / S0006-3223 (96) 00044-3. PMID 9024952. S2CID 42947314.
- ^ Уэно С., Накамура М., Миками М., Кондо К., Исигуро Х., Аринами Т., Комияма Т., Мицусио Х., Сано А., Танабэ Х. (ноябрь 1999 г.). «Идентификация нового полиморфизма гена переносчика дофамина человека (DAT1) и значимая связь с алкоголизмом». Молекулярная психиатрия. 4 (6): 552–7. Дои:10.1038 / sj.mp.4000562. PMID 10578237.
- ^ Уэно С (февраль 2003 г.). «Генетический полиморфизм переносчиков серотонина и дофамина при психических расстройствах». Журнал медицинских исследований. 50 (1–2): 25–31. PMID 12630565.
- ^ Бивер К.М., Райт Дж. П., Делиси М. (сентябрь 2008 г.). «Формирование преступной группы сверстников: свидетельство корреляции гена x окружающей среды». Журнал генетической психологии. 169 (3): 227–44. Дои:10.3200 / GNTP.169.3.227-244. PMID 18788325. S2CID 46592146.
- ^ Университет штата Флорида (2 октября 2008 г.). «Специфический ген, обнаруженный у мужчин-подростков с правонарушителями». ScienceDaily. Получено 8 октября 2008.
- ^ Лаасонен-Балк Т., Куикка Дж., Виинамяки Х., Хуссо-Саастамойнен М., Лехтонен Дж., Тиихонен Дж. (Июнь 1999 г.). «Плотность переносчиков дофамина в полосатом теле при большой депрессии». Психофармакология. 144 (3): 282–5. Дои:10.1007 / s002130051005. PMID 10435396. S2CID 32882588.
- ^ Ng J, Zhen J, Meyer E, Erreger K, Li Y, Kakar N, Ahmad J, Thiele H, Kubisch C, Rider NL, Morton DH, Strauss KA, Puffenberger EG, D'Agnano D, Anikster Y, Carducci C, Хайланд К., Ротштейн М., Леуцци В., Борк Г., Рейт М.Э., Куриан М.А. (апрель 2014 г.). «Синдром дефицита переносчика дофамина: фенотипический спектр от младенчества до взрослого возраста». Мозг. 137 (Pt 4): 1107–19. Дои:10.1093 / мозг / awu022. ЧВК 3959557. PMID 24613933.
- ^ Ротман РБ, Анантан С., Partilla JS, Сайни С.К., Муха-Чафик О, Патак В., Бауманн М.Х. (июнь 2015 г.). «Исследования биогенных переносчиков амина 15. Идентификация новых аллостерических лигандов переносчиков дофамина с наномолярной эффективностью». Журнал фармакологии и экспериментальной терапии. 353 (3): 529–38. Дои:10.1124 / jpet.114.222299. ЧВК 4429677. PMID 25788711.
- ^ Аггарвал С., Лю X, Райс С., Менелл П., Кларк П.Дж., Папароидамис Н., Сяо Ю.С., Сальвино Дж. М., Фонтана А.С., Испания, Р.А. Кортагере, С., Мортенсен О.В. (2019). "Идентификация нового аллостерического модулятора переносчика дофамина человека". ACS Chem Neurosci. 10 (8): 3718–3730. Дои:10.1021 / acschemneuro.9b00262. ЧВК 6703927. PMID 31184115.
- ^ Верзингер К., Сидху А. (апрель 2003 г.). «Ослабление активности переносчика дофамина альфа-синуклеином». Письма о неврологии. 340 (3): 189–92. Дои:10.1016 / S0304-3940 (03) 00097-1. PMID 12672538. S2CID 54381509.
- ^ Ли Ф.Дж., Лю Ф., Приступа З. Б., Низник Х. Б. (апрель 2001 г.). «Прямое связывание и функциональное связывание альфа-синуклеина с переносчиками дофамина ускоряют апоптоз, индуцированный допамином». Журнал FASEB. 15 (6): 916–26. Дои:10.1096 / fj.00-0334com. PMID 11292651.
- ^ Торрес Г.Э., Яо В.Д., Мон А.Р., Куан Х., Ким К.М., Леви А.И., Штаудингер Дж., Карон М.Г. (апрель 2001 г.). «Функциональное взаимодействие между моноаминовыми переносчиками плазматической мембраны и синаптическим белком, содержащим домен PDZ, PICK1». Нейрон. 30 (1): 121–34. Дои:10.1016 / S0896-6273 (01) 00267-7. PMID 11343649. S2CID 17318937.
- ^ Карнейро А.М., Инграм С.Л., Болье Дж.М., Суини А., Амара С.Г., Томас С.М., Карон М.Г., Торрес Г.Е. (август 2002 г.). «Адаптерный белок Hic-5, содержащий множественные LIM-домены, синаптически колокализуется и взаимодействует с транспортером дофамина». Журнал неврологии. 22 (16): 7045–54. Дои:10.1523 / JNEUROSCI.22-16-07045.2002. ЧВК 6757888. PMID 12177201.
- ^ Midde NM, Yuan Y, Quizon PM, Sun WL, Huang X, Zhan CG, Zhu J (март 2015 г.). «Мутации тирозина 88, лизина 92 и тирозина 470 человеческого переносчика дофамина приводят к ослаблению индуцированного Tat ВИЧ-1 ингибирования транспорта дофамина». Журнал нейроиммунной фармакологии. 10 (1): 122–35. Дои:10.1007 / s11481-015-9583-3. ЧВК 4388869. PMID 25604666.
- ^ Midde NM, Хуанг X, Гомес AM, Booze RM, Zhan CG, Zhu J (сентябрь 2013 г.). «Мутация тирозина 470 переносчика дофамина человека имеет решающее значение для индуцированного Tat ВИЧ-1 ингибирования транспорта дофамина и конформационных переходов переносчика». Журнал нейроиммунной фармакологии. 8 (4): 975–87. Дои:10.1007 / s11481-013-9464-6. ЧВК 3740080. PMID 23645138.
- ^ Пурохит В., Рапака Р., Шуртлефф Д. (август 2011 г.). «Наркотики, вызывающие злоупотребление, допамин и нейрокогнитивные расстройства, связанные с ВИЧ / деменция, связанная с ВИЧ». Молекулярная нейробиология. 44 (1): 102–10. Дои:10.1007 / s12035-011-8195-z. PMID 21717292. S2CID 13319355.
внешняя ссылка
- Ассоциации, эксперименты, публикации и клинические испытания переносчиков дофамина
- Дофамин + транспортер в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)
- Обзор всей структурной информации, доступной в PDB за UniProt: Q7K4Y6 (Drosophila melanogaster Натрийзависимый переносчик дофамина) в PDBe-KB.
| Основные статьи и фармацевтические препараты |
| ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Нейрофармакология |
| ||||||||
| Активные метаболиты | |||||||||
| Статьи по Теме |
| ||||||||
| |||||||||