Галлимим - Gallimimus
| Галлимим | |
|---|---|
 | |
| Реконструированный скелет (по образцу взрослого голотип и ювенильная особь), Музей естественной истории, Лондон | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Clade: | Динозаврия |
| Clade: | Заурисхия |
| Clade: | Тероподы |
| Clade: | †Орнитомимозаврия |
| Семья: | †Орнитомимиды |
| Род: | †Галлимим Осмольска и другие., 1972 |
| Разновидность: | †G. bullatus |
| Биномиальное имя | |
| †Галлимим буллатус Осмольска и другие., 1972 | |
| Синонимы | |
Галлимим (/ˌɡæлɪˈмаɪмəs/ GAL-я-МОЙ-məs ) это род из теропод динозавр что жил в том, что сейчас Монголия вовремя Поздний мел период, около семидесяти миллион лет назад (мя). Несколько окаменелостей на разных стадиях роста были обнаружены польско-монгольскими экспедициями в пустыня Гоби Монголии в 1960-е годы; большой скелет, обнаруженный в этом районе, был сделан голотип экземпляр нового рода и вида Галлимим буллатус в 1972 году. Общее название означает "курица имитировать ", имея в виду сходство между его шейными позвонками и позвонками Galliformes. Конкретное название происходит от булла, золотая капсула, которую носил Римский молодежи, имея в виду выпуклую структуру у основания черепа Галлимим. В то время, когда это было названо, окаменелости Галлимим представлены наиболее полными и лучше всего сохранившимися орнитомимид ("страус динозавр ") материал еще не обнаружен, и этот род остается одним из самых известных представителей группы.
Галлимим самый крупный из известных орнитомимид; взрослые были около 6 метров (20 футов) в длину, 1,9 метра (6 футов 3 дюйма) в бедрах и весили около 440 килограммов (970 фунтов). Как свидетельствует его родственник Орнитомим, это было бы были перья. Голова была маленькая и светлая, с большими глазами, обращенными в стороны. Морда была длинной по сравнению с другими орнитомимидами, но шире и закруглялась на конце, чем у других видов. Галлимим был беззубым с ороговевший (роговой) клюв и нежная нижняя челюсть. Многие из позвонки имел отверстия, указывающие на то, что они были пневматический (наполненный воздухом). Шея была пропорционально длинной по отношению к туловищу. Руки были пропорционально самыми короткими среди всех орнитомимозавров, и каждая имела по три пальца с изогнутыми когтями. Передние конечности были слабыми, а задние - пропорционально длинными. Семейство Ornithomimidae является частью группы Орнитомимозаврия. Ансеримим, также из Монголии, считается ближайшим родственником Галлимим.
Как орнитомимид, Галлимим был бы флот (или беглый ) животное, использующее свою скорость, чтобы убежать от хищников; его скорость была оценена в 42–56 км / ч (29–34 миль в час). У него могло быть хорошее зрение и интеллект, сопоставимый с ратит птицы. Галлимим возможно, жили группами, на основании открытия нескольких образцов, сохранившихся в костное ложе. Были предложены различные теории относительно диеты Галлимим и другие орнитомимиды. Очень подвижная шея, возможно, помогала обнаруживать мелкую добычу на земле, но также могла быть оппортунистической. всеядный. Также было высказано предположение, что он использовал небольшие столбчатые структуры в клюве для фильтрующий в воде, хотя эти структуры могли вместо этого быть гребнями, используемыми для питания твердым растительным материалом, что указывает на травоядный рацион питания. Галлимим является наиболее часто встречающимся орнитомимозавром в Формация Немегт, где он жил вместе со своими родственниками Ансеримим и Дейнохейрус. Галлимим был показан в фильме парк Юрского периода, в сцене, которая имела важное значение для истории спецэффекты, и в формировании общего представления о динозаврах как птицеподобных животных.
История открытия

Между 1963 и 1965 гг. Польская Академия Наук и Монгольская Академия Наук организовал польско-монгольские палеонтологические экспедиции в пустыня Гоби Монголии. Среди останков динозавров, обнаруженных в песчаных пластах Бассейн Немегт были многочисленные орнитомимиды на разных стадиях роста из местонахождений Немегт, Цааган Хушуу, Алтан Ула IV и Наран Булак. Собрано три частично полных скелета, два с черепами, а также множество фрагментарных останков. Самый крупный скелет (впоследствии ставший голотипом Галлимим буллатус) был обнаружен палеонтологом Зофья Киелан-Яворовская в Цааган-Хушуу в 1964 г .; он сохранился лежащим на спине, а череп был найден под его тазом. Один небольшой экземпляр был найден в том же году в Цааган Хушуу, а другой небольшой экземпляр был найден в местности Немегт. Небольшой скелет без передних конечностей был обнаружен в 1967 году монгольской палеонтологической экспедицией в Бугин Цав за пределами бассейна Немегт. Окаменелости были размещены в монгольских, польских и Академия Наук СССР.[1][2][3] Польско-монгольские экспедиции были известны тем, что возглавлялись женщинами, некоторые из которых были среди первых женщин, давших имена новым динозаврам. Окаменелости, обнаруженные в ходе этих экспедиций, проливают новый свет на обмен фауной между Азией и Северной Америкой во времена Меловой период.[4][5][6] Некоторые из скелетов были выставлены в Варшаве в 1968 году, они сидели в высоких полустоячих позах, что было принято в то время, хотя сегодня предпочитают более горизонтальные позы.[6]

В 1972 г. палеонтологи Halszka Osmólska, Ева Роневич и Ринчен Барсболд назвал новый род и виды Галлимим буллатус, используя самый крупный собранный скелет, образец IGM 100/11 (из Цааган Хушуу, ранее называвшийся G.I. No.DPS 100/11 и MPD 100/11), как голотип. Общее название происходит от латинский галл, "курица" и греческое мимос, «мимический», по отношению к передней части шейных позвонков, которые напоминают позвонки Galliformes. В конкретное имя происходит от латинского булла, золотая капсула, которую носил Римский молодость на шее, по отношению к выпуклой капсуле на парасфеноиде у основания черепа динозавра. Такая особенность не была описана у других рептилий в то время и считалась необычной. Голотип состоит из почти полного скелета с искаженной мордой, неполной нижней челюсти, позвоночного ряда, таза, а также некоторых недостающих костей кисти и стопы.[1][7] Остальные частично завершенные скелеты принадлежали молодым особям; ZPAL MgD-I / 1 (из Цааган Хушуу) имеет раздробленный череп с отсутствующим верхушкой, поврежденными позвонками, фрагментированными ребрами, грудным поясом и передними конечностями, а также неполной левой задней конечностью, ZPAL MgD-I / 94 (из местности Немегт) отсутствует череп, атлас, кончик хвоста, грудной пояс и передние конечности, а у самого маленького экземпляра, IGM 100/10 (из Бугин Цав), отсутствует грудной пояс, передние конечности и несколько позвонков и ребер. Осмольска и его коллеги перечислили всего двадцать пять известных образцов, девять из которых были представлены только отдельными костями.[1][8]

В то время, когда это было названо, окаменелости Галлимим представляли собой наиболее полный и лучше всего сохранившийся материал орнитомимид, из когда-либо обнаруженных, и этот род остается одним из самых известных представителей группы. Орнитомимиды ранее были известны в основном из Северной Америки, Археорнитомим единственный известный до этого член из Азии (хотя и без черепа). После первых открытий новые образцы были найдены в ходе международных экспедиций под руководством Монголии.[1][8][5][6] Три из Галлимим скелеты (включая голотип) позже стали частью передвижной выставки окаменелостей монгольских динозавров, которая объехала различные страны.[9][10] Браконьерство на ископаемые стала серьезной проблемой в Монголии в 21 веке, и несколько Галлимим образцы были разграблены. В 2017 году Ханг-Дже Ли и его коллеги сообщили об обнаруженном в 2009 году следе окаменелостей, связанном со сжатым Галлимим стопа (экз. MPC-D100F / 17). Остальная часть скелета, похоже, была удалена браконьерами вместе с несколькими другими. Галлимим образцы (на что указывают пустые котлованы, мусор и разбросанные сломанные кости в карьере). Необычно найти следы, тесно связанные с окаменелостями тела; некоторые следы соответствуют ногам орнитоимимидов, а другие принадлежат другим динозаврам.[11][12] В 2014 году плита с двумя Галлимим образцы были репатриированы в Монголию вместе со скелетами других динозавров после контрабанды в США.[13]
В 1988 году палеонтолог Грегори С. Пол пришли к выводу, что черепа орнитомимид были более похожи друг на друга, чем считалось ранее, и переместили большинство видов в один и тот же род, Орнитомим, в результате чего новая комбинация О. bullatus.[14] В 2010 году он вместо этого перечислил это как "Галлимим (или же Струтиомим) буллатус", но вернулся к использованию только названия рода Галлимим в 2016 году.[15][16] Участвующие виды обычно содержались в отдельных родах другими авторами.[17] Орнитомимидный позвонок из Японии неофициально названный "Sanchusaurus" был опубликован в журнале 1988 г., но был назначен Галлимим sp. (неопределенных видов) палеонтологом Дун Чжимин и коллеги в 1990 году.[7] Барсболд неофициально назвал почти полный скелет (IGM 100/14) «Gallimimus mongoliensis», но поскольку он отличается от Галлимим в некоторых деталях Ёсицугу Кобаяси и Барсболд в 2006 году предположили, что он, вероятно, принадлежит к другому роду. В 2000 году палеонтолог Филип Дж. Карри предложил, чтобы Ансеримим, который известен только по одному скелету из Монголии, был младший синоним из Галлимим, но это было отклонено Кобаяши и Барсболд, которые указали на несколько различий между ними. Барсболд отметил некоторые морфологический вариация среди новых Галлимим образцы, хотя это никогда не публиковалось.[8]
Описание
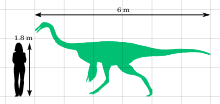
Галлимим является крупнейшим известным членом семья Ornithomimidae. Взрослый голотип (экземпляр IGM 100/11) был около 6 метров (20 футов) в длину и 1,9 метра (6,2 фута) в бедре; его череп был 330 миллиметров (1,08 фута) в длину, а бедренная кость (бедренная кость) составляла 660 миллиметров (2,17 фута). Он весил бы около 440 килограммов (970 фунтов). Для сравнения, один молодой особь (ZPAL MgD-I / 94) был около 2,15 метра (7,1 фута) в длину, 0,79 метра (2,6 фута) в высоту в бедре и весил примерно 26 килограммов (57 фунтов).[14] На основе окаменелостей родственных Орнитомим, известно, что орнитомимозавры мы пернатый, и что взрослые особи несли структуры, похожие на крылья, о чем свидетельствует наличие игольчатых выступов на локтевая кость кость нижней части руки, неровности, указывающие на то, где могли бы прикрепиться перья.[18]
Череп

Глава Галлимим был очень маленьким и легким по сравнению с позвоночником. Из-за длины морды череп был длиннее по сравнению с другими орнитомимидами, а морда имела слегка выпуклый покатый верхний профиль. Боковой профиль рыла отличался от других орнитомимид тем, что не сужался к передней половине, а нижний передний край рыла предчелюстная кость передняя часть верхней челюсти поднимается вверх, а не горизонтально. При взгляде сверху морда была почти лопатообразной (ложкообразной), широкой и закругленной на кончике (или U-образный ), тогда как у североамериканских орнитомимид он был острым (или V-образным). В орбиты (глазницы) были большими и обращенными вбок, как у других орнитомимид. В височная область сбоку черепа за глазами был глубокий, а подвисочное отверстие (нижнее отверстие за орбитой) было почти треугольным и меньше, чем у соответствующего Струтиомим. У него были глубокие мышечные рубцы в задней части крыши черепа, вдоль теменная кость. В парасфеноид (кость черепной коробки на нижней стороне основания черепа) была тонкостенной, полой и образовывала грушевидную луковичную структуру. Структура имела неглубокую борозду, которая открывалась вперед. В внутренние ноздри (внутренние отверстия для носового хода) были большими и располагались далеко позади нёбо, благодаря наличию обширного вторичное небо, что было обычным для орнитомимид.[1][17][19][20]
Нежная нижняя челюсть, состоящая из тонких костей, была тонкой и неглубокой спереди, углубляясь к задней части. Передняя часть нижней челюсти была похожа на лопату, что давало зазор между кончиками челюстей в закрытом состоянии. Форма лопаты была похожа на форму обыкновенная чайка, а нижний клюв мог иметь форму, похожую на форму этой птицы. Ретроартикулярный отросток в задней части челюсти (где прикрепляются мышцы челюсти, открывающие клюв) был хорошо развит и состоял в основном из угловая кость. В надугловой была самой большой костью нижней челюсти, которая обычна у теропод. В нижнечелюстное отверстие отверстие в нижней челюсти, обращенное в сторону, было удлиненным и сравнительно небольшим. Нижняя челюсть не имела венечный отросток или надзубная кость, отсутствие которой характерно для клювовидных теропод (орнитомимозавров, овирапторозавры, теризинозавры и птицы), но необычный среди теропод в целом.[1][21] Челюсти Галлимим мы беззубый (беззубый), а передняя часть была бы покрыта ороговевший rhamphotheca (роговой клюв) в жизни. Клюв мог покрывать меньшую площадь, чем у североамериканских родственников, из-за отсутствия питающих отверстий на верхней челюсти. Внутренняя сторона клюва имела небольшие, плотно расположенные и равномерно расположенные столбчатые структуры (их точная природа обсуждается), которые были самыми длинными спереди и укорачивались к спине.[19][21][22]
Посткраниальный скелет

Галлимим было 64–66 позвонки в его позвоночник, меньше, чем у других орнитомимид. Центры (или тела) позвонков пластинчатые, с плоской передней поверхностью и вогнутой задней поверхностью, за исключением первых шести. каудальный (хвостовой) позвонок, где задняя поверхность также была плоской, и позвонки на конце хвоста, которые были амфиплатическими с плоскими обеими поверхностями. Многие из центров имели отверстие (отверстия, которые также назывались "плевроцели "), и поэтому, вероятно, пневматический (с их полыми камерами, захваченными воздушные мешочки ). Шея состояла из 10 шейные позвонки, которые все были длинными и широкими, за исключением атласная кость (первый позвонок, который соединяется с задней частью черепа). Атлас отличался от других орнитомимид тем, что передняя поверхность его промежуточного центра была наклонена вниз к спине, вместо того, чтобы быть вогнутой и обращенной вверх для поддержки затылочный мыщелок. Шея была пропорционально длиннее туловища, чем у других орнитомимид. Шея была разделена на две отдельные части: шейные позвонки спереди имели центры, которые были почти треугольными на виде сбоку и сужались к спине, а также низкие нервные дуги и короткие, широкие зигапофизы (в процессы сочлененный между позвонками); шейные позвонки на спине имели катушкообразные центры, которые постепенно становились выше, и длинные тонкие зигапофизы. Пневматические отверстия здесь были маленькими и овальными, а нервные отростки, выступающие наружу из центров, образовывали длинные, низкие и острые гребни, за исключением самых задних шейных позвонков.[1][23]

Задняя часть Галлимим было 13 спинные позвонки, с катушкообразными центрами, которые были короткими, но имели тенденцию становиться глубже и длиннее к спине. Их поперечные отростки (сочленяющиеся с ребрами) немного увеличиваются в длину к спине. Два первых дорсальных центра имели глубокие пневматические отверстия, в то время как остальные имели только неглубокие отверстия. ямки (впадины), и были видны нервные отростки, имеющие несколько треугольную или прямоугольную форму. В крестец (сросшиеся позвонки между тазовыми костями) состояли из пяти крестцовые позвонки которые были примерно равной длины. Центры здесь были катушковидными, уплощенными сбоку и имели ямки, которые, по-видимому, продолжались как глубокие отверстия у некоторых экземпляров. Нервные отростки здесь были прямоугольными, широкими и выше, чем спинные позвонки. Они были выше или равны по высоте верхнему краю подвздошный лезвие и были отдельными, тогда как у других орнитомимидов они были сплавлены вместе. Хвост имел 36–39 хвостовых позвонков, при этом центры из передних имели форму катушки, а задние были почти треугольными и удлиненными в поперечнике. Нервные отростки здесь были высокими и плоскими, но уменьшились назад, где они стали похожи на гребни. Единственным признаком пневматичности хвоста были глубокие ямки между нервными отростками и поперечный отросток двух первых хвостовых позвонков. Все позвонки перед крестцом растоплены ребра кроме атласа и последнего спинного позвонка.[1][23]
В лопатка (лопатка) была короткой и изогнутой, тонкой на переднем конце и толстой сзади. Это было относительно слабо связано с коракоидный, который был большим и глубоким сверху донизу. В целом передние конечности не сильно отличались от таковых у других орнитомимид, все они были сравнительно слабыми. В плечевая кость (кость плеча), которая имела почти круглое поперечное сечение, была длинной и искривленной. Дельто-грудной гребень в верхней передней части плечевой кости был сравнительно небольшим и, следовательно, имел небольшую поверхность для прикрепления мышц плеча. Локтевая кость была тонкой, длинной и слабо изогнутой, с почти треугольным стержнем. В олекранон (выступ от локтя) был заметен у взрослых, но недостаточно развит у молодых. В радиус (другая кость в нижней части руки) была длинной и тонкой с более расширенным верхним концом по сравнению с нижним. В манус (рука) была пропорционально короткой по сравнению с таковыми у других орнитомимозавров, имела наименьшее соотношение длины кисти к длине плечевой кости среди всех членов группы, но в остальном была схожей по структуре. У него было три пальца, которые были одинаково развиты; первый («большой палец») был самым сильным, третий - самым слабым, а второй - самым длинным. В ногтевые (копытные кости) были сильными, несколько изогнутыми (наиболее изогнутыми был первый палец) и сдавлены вбок с глубокими бороздками с каждой стороны. Когтевые кости были развиты аналогично, хотя третья была немного меньше.[1][8][17]
В лобок (лобковая кость) была длинной и тонкой, заканчивалась лобковым ботинком, расширяющимся к передней и задней части, что является общей чертой орнитомимозавров. Задние конечности мало отличались от таковых других орнитомимид и были пропорционально длиннее, чем у других теропод. Бедро было почти прямым, длинным и тонким, с уплощенным вбок стержнем. В большеберцовая кость был прямым, длинным, с двумя хорошо развитыми мыщелки (закругленный конец кости) на верхнем конце и уплощенный нижний конец. В малоберцовая кость голени была плоской, тонкой и широкой на верхнем конце, сужающейся к нижнему. Нижняя половина третьего плюсневой при осмотре был широким, частично закрывая две смежные плюсневые кости с каждой стороны, но резко сужался в середине длины, заклинивая между этими костями и исчезая ( арктометатарсалий структура стопы). Третий палец по отношению к конечности был пропорционально короче, чем у других орнитомимид. Как и у других орнитомимид, стопа не имела Hallux (или dewclaw, первый палец большинства других теропод). Когти пальцев ног с нижней стороны плоские; два внешних слегка отклонились от своих цифр.[1][17]
Классификация


Осмольска и его коллеги назначили Галлимим к семейству Ornithomimidae в 1972 г. с североамериканскими Струтиомим как ближайший родственник, сокрушаясь о том, что сравнение между таксоны было сложно, потому что другие известные в то время орнитомимиды либо плохо сохранились, либо неадекватно описаны.[1] В 1975 году Киелан-Яворовска заявила, что, хотя многие динозавры из Азии были помещены в те же семьи, что и североамериканские родственники, эта категория классификации имела тенденцию быть более всеобъемлющей, чем та, которая используется для современных птиц. Она подчеркнула, что пока Галлимим имели закругленный клюв (похожий на клюв гуся или утки), у североамериканских орнитомимид - заостренный клюв - разница, которая в противном случае заставила бы систематиков относить современных птиц к разным семьям.[5] В 1976 году Барсболд поместил орнитомимид в новую группу орнитомимозавров («страусиные динозавры»). В 2003 году Кобаяши и Джун-Чанг Лю обнаружили, что Ансеримим был сестринский таксон к Галлимим, оба образуют полученный (или "продвинутый") клады с североамериканскими родами, что подтвердили Кобаяши и Барсболд в 2006 г.[8][19]
Следующее кладограмма показывает размещение Галлимим среди орнитомимид, по данным Ли Сюй и его коллег, 2011 г .:[24]
| Орнитомимиды |
| ||||||||||||||||||||||||||||||||||||
Орнитомимозавры принадлежали к кладе Maniraptoriformes из целурозавр теропод, в том числе современных птиц. У ранних орнитомимозавров были зубы, которые были потеряны у более производных членов группы.[25] В 2004 году Маковицкий, Кобаяши и Карри предположили, что большая часть ранней эволюционной истории орнитомимозавров имела место в Азии, где было обнаружено большинство родов, включая наиболее базальный (или «примитивных») таксонов, хотя они признали, что наличие базальных Пелеканимимус в Европе представляет собой сложность классификации. Группа, должно быть, раз или два разошлась из Азии в Северную Америку через Берингия для учета найденных здесь позднемеловых родов. Как видно из некоторых других групп динозавров, орнитомимозавры были в основном ограничены Азией и Северной Америкой после того, как Европа была отделена от Азии Тургайский пролив.[17]
В 1994 году палеонтолог Томас Р. Хольц сгруппированные орнитомимозавры и троодонтиды в кладе, основанной на общих чертах, таких как наличие луковичной капсулы на парасфеноиде. Он назвал кладу Bullatosauria, исходя из конкретного названия Галлимим буллатус, что также относилось к капсуле.[26] В 1998 году Хольц вместо этого обнаружил, что троодонтиды были базальными манирапторами, а это означало, что все представители этой клады попали в состав Bullatosauria, который, следовательно, стал младшим синонимом Maniraptoriformes, и с тех пор эта клада вышла из употребления.[27][28]
Палеобиология

Шейные позвонки Галлимим указывают на то, что он держал шею наискосок, наклоняясь вверх под углом 35 градусов. Осмольска и его коллеги обнаружили, что руки Галлимим не были цепкий (или способный схватить), и что большой палец не противопоставлен. Они также предположили, что руки были слабыми по сравнению, например, с руками орнитомимозавра. Дейнохейрус. Они согласились с интерпретациями биологии орнитомимид палеонтологами. Дейл Рассел с начала 1972 года, включая то, что они были бы очень флотом (или беглый ) животные, хотя и менее подвижные, чем крупные современные наземные птицы, и использовали бы свою скорость, чтобы убежать от хищников. Рассел также предположил, что у них хорошее зрение и интеллект, сопоставимый с современными. ратит птицы. Поскольку их хищники могли иметь цветовое зрение, он предположил, что это повлияло бы на их окраску, что, возможно, привело бы к камуфляж.[1][29] В 1982 году палеонтолог Ричард А. Талборн подсчитал, что Галлимим мог двигаться со скоростью 42–56 км / ч (29–34 миль в час). Он обнаружил, что орнитимимиды не были бы такими быстрыми, как страусы, которые могут развивать скорость до 70–80 км / ч (43–49 миль в час), отчасти из-за того, что их руки и хвосты увеличивают их вес.[30]
В 1988 году Пол предположил, что глазные яблоки орнитомимидов сплюснуты и обладают минимальной подвижностью внутри глазниц, что требует движения головы для просмотра объектов. Поскольку их глаза смотрели вбок, чем у некоторых других птицеподобных теропод, их бинокулярное зрение был бы более ограниченным, что является адаптацией некоторых животных, улучшающей их способность видеть хищников позади себя. Пол считал относительно короткие хвосты, уменьшающие вес, и отсутствие больших пальцев у орнитомимидов приспособлением к скорости. Он предположил, что они могли защитить себя клеванием и ногами, но в основном полагались на свою скорость для побега.[14] В 2015 году Акинобу Ватанабэ и его коллеги обнаружили, что вместе с Дейнохейрус и Археорнитомим, Галлимим имел самый пневматизированный скелет среди орнитомимозавров. Пневматизация считается полезной для полета у современных птиц, но ее функция у нептичьих динозавров точно неизвестна. Было высказано предположение, что пневматизация использовалась для уменьшения массы крупных костей, что было связано с высоким метаболизм, балансировать во время передвижения или использоваться для терморегуляция.[23]

В 2017 году Ли и его коллеги предложили различные возможные тафономический обстоятельства (изменения во время распада и фоссилизации), чтобы объяснить, как Галлимим нога, обнаруженная в 2009 году, была связана с путепроводом. Дорога сохранилась в песчаник в то время как ступня сохраняется в аргиллит, простираясь на 20 см (7,9 дюйма) ниже слоя с дорожками. Возможно, окаменелость представляет собой животное, которое умерло на своем пути, но глубина ступни в грязи может быть слишком мелкой, чтобы она могла увязнуть. Возможно, он был убит наводнением, после чего был похоронен в пруду. Однако слои грязи и песка указывают не на наводнение, а, вероятно, на сухую среду, и отложения вокруг окаменелости указывают на то, что животное было живым, когда пришло в этот район. Таким образом, авторы предположили, что следы были созданы в течение длительного периода времени и периода сушки, и что, вероятно, ни один из них не был создан человеком, которому принадлежала стопа. Животное могло пройти по дну пруда, пробив слой наносов со следами, пока он был намокшим от дождя или содержал воду. Животное могло умереть в этом положении от жажды, голода или по другой причине, а затем грязь осела на песке, тем самым покрывая и сохраняя следы и тушу. При разложении ступня могла быть сжатой и расчлененной, что привело к сухожилия flex, а позже на него наступили тяжелые динозавры. Область могла быть единственной костное ложе (исходя из возможного количества браконьеров), представляющих собой Галлимим массовая смертность, возможно, из-за засухи или голода. Тот факт, что животные, по всей видимости, умерли одновременно (пустые котлованы были стратиграфически идентичны), может указывать на то, что Галлимим был общительный (жили группами), что также предполагалось для других орнитомимид.[12]
Кормление и диета

Осмольска и его коллеги отметили, что передняя часть шеи Галлимим были бы очень подвижными (задняя часть была более жесткой), нервные дуги в позвонках в этой области были похожи на курицу и других Galliformes, что указывает на схожие пищевые привычки. Они нашли клюв Галлимим похож на утку или гуся, и что он питался бы маленькой живой добычей, которую глотал целиком. Подвижность шеи могла быть полезна при обнаружении добычи на земле, поскольку глаза располагались по бокам черепа. Они предположили, что у всех орнитомимид были одинаковые пищевые привычки, и указали, что Рассел сравнивал клювы орнитомимид с клювами насекомоядный птицы. Осмольска и его коллеги предположили, что Галлимим был способен черепной кинезис (из-за кажущейся неплотной связи между некоторыми костями в задней части черепа), особенность, которая позволяет отдельным костям черепа перемещаться относительно друг друга. Они также предположили, что он использовал свои короткие передние конечности не для того, чтобы подносить пищу ко рту, а для того, чтобы копать или копать землю для доступа к пище.[1] Руки Галлимим могли быть слабее, чем, например, Струтиомим, который мог вместо этого использовать свои руки для захвата и захвата, согласно статье 1985 года палеонтологов Элизабет Л. Николлс и Энтони П. Рассела.[31]
В 1988 году Пол не согласился с тем, что орнитомимиды были всеядные которые ели мелких животных и яйца, а также растения, как предполагалось ранее. Он отметил, что страусы и эму в основном травоядные и браузеры, и что черепа орнитомимидов были наиболее похожи на черепа вымерших моа, которые были достаточно сильными, чтобы откусить веточки, о чем свидетельствует их внутреннее содержимое. Он также предположил, что орнитомимиды были хорошо приспособлены для просмотра твердых растений и использовали свои руки, чтобы подносить ветви к своим челюстям.[14] Палеонтолог Йорн Хурум предположил в 2001 году, что из-за схожего строения челюстей, Галлимим возможно, у них была оппортунистическая, всеядная диета, как у чаек. Он также заметил, что плотный внутринижнечелюстной сустав предотвратит любое движение между передней и задней частями нижней челюсти.[21]

В 2001 году палеонтологи Марк А. Норелл, Маковицки и Карри сообщили о Галлимим череп (IGM 100/1133) и Орнитомим череп, сохранивший структуры мягких тканей на клюве. Внутренняя сторона Галлимим клюв имел столбчатые структуры, которые авторы нашли похожими на ламели в клюве гусеобразный птицы, которые используют их для манипулирования пищей, процеживания отложений, фильтрующий путем отделения пищевых продуктов от других материалов и для срезания растений во время выпаса. Они нашли Северная лопата, который питается растениями, моллюсками, остракоды, и фораминиферы, чтобы быть современной гусеобразной формы со структурами, наиболее похожими по анатомии на анатомические структуры Галлимим. Авторы отметили, что орнитомимиды, вероятно, не использовали клюв для охоты на крупных животных и были многочисленны в мезический окружающей среде, хотя и реже в более засушливых средах, что позволяет предположить, что они могли зависеть от водных источников пищи.[22] Если эта интерпретация верна, Галлимим был бы одним из крупнейших известных наземных фильтров-фидеров.[17]
В 2005 году палеонтолог Пол Барретт указал, что ламеллоподобные структуры Галлимим не представляли собой гибкие щетинки, как у птиц, питающихся фильтром (поскольку нет никаких указаний на то, что эти структуры перекрываются или разрушаются), а были скорее похожи на тонкие, регулярно расположенные вертикальные гребни в клювах черепах и гадрозаврид динозавры. Считается, что у этих животных такие гребни связаны с диетами травоядных, используемых для выращивания жесткой растительности. Барретт предположил, что гребни в клюве Галлимим представляет собой естественный слепок внутренней поверхности клюва, что указывает на то, что животное было травоядным, питающимся материалом с высоким содержанием клетчатки. Открытие многих гастролиты (желудок камни) у некоторых орнитомимидов указывают на наличие мельницы желудка и, следовательно, указывают на диету для травоядных, так как они используются для измельчения пищи животных, у которых отсутствует необходимый жевательный аппарат. Барретт также подсчитал, что 440 кг (970 фунтов) Галлимим потребовалось бы от 0,07 до 3,34 кг (0,15-7,36 фунта) еды в день, в зависимости от того, есть ли у него эндотермический или экзотермический («теплый» или «холодный» - кровяной) метаболизм, потребление которого, как он обнаружил, было невозможным, если бы это был фильтр-питатель. Он также обнаружил, что орнитомимиды были в изобилии не только в формациях, которые представляли мезическую среду, но и в засушливых средах, где было недостаточно воды для поддержания диеты, основанной на питании через фильтр.[20] В 2007 году палеонтолог Эспен М. Кнутсен написал, что форма клюва орнитомимид, по сравнению с таковыми у современных птиц, соответствует всеядности или травоядности с высоким содержанием клетчатки.[32]
Разработка

Форма и пропорции черепа значительно менялись в процессе роста. Задняя часть черепа и орбиты уменьшились в размерах, а морда стала относительно длиннее; аналогичные изменения происходят в современных крокодилы. Череп также был пропорционально больше у более молодых особей, а наклон верхнего профиля рыла был менее заметным. Ребра на шее срослись с позвонками только у взрослых. Похоже, что передние конечности пропорционально удлинились в процессе роста, тогда как пропорциональная длина костей задних конечностей изменилась очень мало.[1][21] В 2012 году палеонтолог Дарла К. Зеленицкий и его коллеги пришли к выводу, что, поскольку взрослые орнитомимозавры имели на руках структуры, похожие на крылья, а молодые - нет (о чем свидетельствуют образцы Орнитомим) эти конструкции изначально были вторичные половые признаки, которые могли быть использованы для репродуктивного поведения, например ухаживание, отображать, и задумчивый.[18]
Исследование, проведенное в 1987 году биологами Романом Павлицким и П. Болехала, показало возрастные различия в содержании кальций и фосфор (важные компоненты в формировании кости) Галлимим образцы. Они обнаружили, что это соотношение было самым высоким у животных молодого и среднего возраста, уменьшаясь с возрастом.[33] В 1991 году они сообщили, что в костях старых людей содержится больше всего свинца и железа, а в костях молодых животных - меньше.[34] Исследование кости гистология различных динозавров в 2000 году биологами Джоном М. Ренсбергером и Махито Ватабе показали, что Canaliculi (каналы, соединяющие костные клетки) и коллаген пучки волокон Галлимим и другие орнитомимиды были больше похожи на птиц, чем на млекопитающих, в отличие от орнитомимидов орнитисхий динозавры, которые были больше похожи на млекопитающих. Эти различия могли быть связаны с процессом и скоростью формирования кости.[35]
Палеоокружающая среда

Галлимим известно из Формация Немегт в пустыне Гоби на юге Монголии. Этот геологическая формация никогда не был датировано радиометрически, но фауна присутствующие в летописи окаменелостей указывают на то, что они, вероятно, были отложены во время Маастрихтский этап, в конце Поздний мел около 70 миллионов лет назад.[36][37][38] Осадки Галлимим тип местности Цааган Хушуу состоит из илы, алевролиты, аргиллиты, пески, а также реже маломощные пласты песчаников.[3] Рок фации формации Немегт предполагают наличие русел рек, илистых отмелей, мелководных озер и поймы в среде, подобной Дельта Окаванго современного Ботсвана.[39] Крупные русла рек и отложения почвы свидетельствуют о значительно более влажном климате, чем в более старых Барун Гойот и Джадохта образования, хотя Caliche отложения указывают на периодические засухи.[40] Ископаемые кости из бассейна Немегт, в том числе Галлимим, более радиоактивный чем окаменелости из окрестностей, возможно, потому что уран накапливается в костях, переносится туда просачиванием грунтовых вод.[6][41]
Реки Немегт, где Галлимим жили, были домом для самых разных организмов. Случайный моллюск окаменелости, а также множество других водных животных, таких как рыбы, черепахи,[36] и крокодиломорфы, включая Шамозух, были обнаружены в этом регионе.[42] Млекопитающее окаменелости редки в формации Немегт, но многие птицы, в том числе энантиорнитин Гурилина, то гесперорнитиформный Юдинорнис, а также Тевиорнис, возможная гусеобразная форма. Среди травоядных динозавров, обнаруженных в формации Немегт, есть анкилозавриды Такие как Тархия, то пахицефалозавр Prenocephale, крупные гадрозавриды, такие как Зауролоф и Барсболдия, и зауроподы, такие как Немегтозавр и Опистоцеликаудия.[36][43] Хищные тероподы, жившие рядом Галлимим включают тираннозавроиды Такие как Тарбозавр, Алиорамус и Багараатан, и троодонтиды, такие как Бороговия, Tochisaurus и Занабазар. Травоядные или всеядные тероподы включают теризинозавров, таких как Теризинозавр, а также овирапторозавры, такие как Эльмизавр, Немегтомайя, и Ринчения.[44] Другие орнитомимозавры, включая Ансеримим и Дейнохейрус, тоже встречаются, но Галлимим - самый распространенный член группы в Немегте.[8]
Культурное значение

Галлимим был показан в фильме 1993 года парк Юрского периода режиссером Стивен Спилберг; похожая сцена в оригинале Роман 1990 года вместо этого фигурировали гадрозавры. Спилберг хотел давка эпизод со стадами животных в фильме, но не знал, как этого добиться, и изначально планировалось визуализировать через покадровая анимация. В то время мало кто верил в создание животных с помощью компьютерная анимация, но визуальный эффект Компания Промышленный свет и магия продюсеры фильма дали добро на изучение возможностей. ILM создал Галлимим скелета в компьютере и анимировал тест, показывающий стадо бегущих скелетов, а затем Тиранозавр погоня за полностью обработанным Галлимим стадо. Производственная группа была полна энтузиазма, поскольку ничего подобного ранее не было достигнуто, и Спилберга убедили записать сцену в сценарий, а также использовать компьютерную графику для других кадров динозавров в фильме вместо покадровой анимации.[45][46] В Галлимим были оживлены путем отслеживания кадров из видеозаписей страусов и видеозаписи выпаса газели также упоминалось.[47] Келан-Яворовска, открывшая образец голотипа, назвала это «красивой сценой».[6] Динозавры в фильме были одним из самых широко разрекламированных приложений компьютерные изображения в кино, и считались более реалистичными, чем то, что было ранее достигнуто с спецэффекты.[48]
Подчеркивая птичье стайное поведение Галлимим стадо было точкой в Парк Юрского периода's истории, поскольку они должны были представлять предшественников птиц. Было показано, что стадо движется как единое целое, а не отдельные животные, бегающие вокруг. Галлимим были показаны в центре группы, как если бы они находились под защитой.[45] Во время сцены палеонтолог Алан Грант говорит, что стадо движется «равномерно, меняет направление, как стая птиц, уклоняющихся от хищника» и «держу пари, что вы больше никогда не будете смотреть на птиц так же», наблюдая за движениями быстрых, изящных Галлимим. Это контрастировало с тем, как динозавры традиционно изображались в средствах массовой информации как неуклюжие, волочащиеся за хвосты животные, а фильм помог изменить общее восприятие динозавров. Эта и другие сцены отражали недавние теории эволюция птиц поощряется научным руководителем фильма, палеонтологом Джон Р. Хорнер, идеи, которые в то время все еще вызывали споры.[49][50][51] Несмотря на такие теории, Галлимим и другие динозавры из фильма были изображены без перьев, отчасти потому, что в то время было неизвестно, насколько широко они были распространены среди группы.[49][52]
Утверждалось, что Следы Lark Quarry (одна из крупнейших в мире концентраций следов динозавров) в Квинсленд, Австралия, послужила источником вдохновения и "научной поддержки" для Галлимим сцена давки в парк Юрского периода; Эти следы первоначально были интерпретированы как представление о давке динозавров, вызванной прибытием хищника-теропода. Идея о том, что следы представляют собой давку, с тех пор оспаривалась («теропод», возможно, был травоядным, похожим на Муттабурразавр ) и консультантом парк Юрского периода отрицает, что треки послужили вдохновением для фильма.[53][54][55]
Смотрите также
Рекомендации
- ^ а б c d е ж грамм час я j k л м п Osmolska, H .; Roniewicz, E .; Барсболд Р. (1972). "Новый динозавр, Галлимим буллатус п. gen., n. sp. (Ornithomimidae) из верхнего мела Монголии » (PDF). Палеонтология Полоника. 27: 103–143.
- ^ Kielan-Jaworowska, Z .; Довчин, Н. (1968). «Рассказ о польско-монгольских палеонтологических экспедициях 1963–1965 гг.» (PDF). Palaeontologica Polonica: 7–30.
- ^ а б Gradzihski, R .; Kazmierczak, J .; Лефельд, Дж. (1969). «Географические и геологические данные польско-монгольских палеонтологических экспедиций» (PDF). Palaeontologica Polonica. 19: 33–82.
- ^ Додсон, П. (1998). Рогатые динозавры: естественная история. Принстон: Издательство Принстонского университета. п. 9. ISBN 978-0-691-05900-6.
- ^ а б c Келан-Яворовска, З. (1975). «Позднемеловые млекопитающие и динозавры из пустыни Гоби: окаменелости, раскопанные польско-монгольскими палеонтологическими экспедициями 1963–1971 годов, проливают новый свет на примитивных млекопитающих и динозавров, а также на обмен фауной между Азией и Северной Америкой». Американский ученый. 63 (2): 150–159. Bibcode:1975AmSci..63..150K. JSTOR 27845359.
- ^ а б c d е Келан-Яворовска, З. (2013). В погоне за ранними млекопитающими. Жизнь прошлого. Блумингтон: Издательство Индианского университета. С. 47–48, 50–51. ISBN 978-0-253-00824-4.
- ^ а б Глут, Д. Ф. (1997). Динозавры: Энциклопедия. Джефферсон: McFarland & Co., стр.429 –432. ISBN 978-0-89950-917-4.
- ^ а б c d е ж Кобаяши, Ю .; Барсболд Р. (2006). "Орнитомимиды из немегтской свиты Монголии" (PDF). Журнал Палеонтологического общества Кореи. 22 (1): 195–207.
- ^ «Монгольские динозавры мелового периода». DinoCasts.com. Архивировано из оригинал 2 февраля 2015 года.
- ^ Oest, S. G .; Hansen, S .; Ост, Т. (1997). "Динозавр, æg og unger - en særudstilling om en forsvunden verden" (PDF). ГеологискНыт (на датском). 6: 24–25. Архивировано из оригинал (PDF) на 2018-05-14. Получено 2018-05-13.
- ^ Fanti, F .; Bell, P.R .; Tighe, M .; Милан, Л. А .; Динелли, Э. (2018). «Геохимический дактилоскопический анализ как инструмент репатриации останков выловленных браконьерами динозавров в Монголии: пример для местности Немегт, пустыня Гоби». Палеогеография, палеоклиматология, палеоэкология. 494: 51–64. Bibcode:2018ППП ... 494 ... 51F. Дои:10.1016 / j.palaeo.2017.10.032.
- ^ а б Ли, Х.-Дж .; Lee, Y.-N .; Adams, T. L .; Карри, П. Дж .; Кобаяши, Ю .; Jacobs, L. L .; Коппельхус, Э. Б. (2018). "Следы теропод, связанные с Галлимим скелет стопы из свиты Немегт, Монголия ". Палеогеография, палеоклиматология, палеоэкология. 494: 160–167. Bibcode:2018ППП ... 494..160л. Дои:10.1016 / j.palaeo.2017.10.020.
- ^ Ганнон, М. (2014). «Украденное« гнездо динозавров »возвращено в Монголию». uk.news.yahoo.com. LiveScience.com. Получено 21 мая 2019.
- ^ а б c d Пол, Г.С. (1988). Хищные динозавры мира. Нью-Йорк: Саймон и Шустер. стр.384–386, 393–394. ISBN 978-0-671-61946-6.
- ^ Пол, Г.С. (2016). Принстонский полевой путеводитель по динозаврам (2-е изд.). Принстон: Издательство Принстонского университета. п. 131. ISBN 978-0-691-16766-4.
- ^ Пол, Г.С. (2010). Принстонский полевой путеводитель по динозаврам (1-е изд.). Принстон: Издательство Принстонского университета. стр.112–114. ISBN 978-0-691-13720-9.
- ^ а б c d е ж Makovicky, P.J .; Кобаяши, Ю .; Карри, П. Дж. (2004). «Орнитомимозаврия». В Weishampel, D. B .; Dodson, P .; Осмольска, Х. (ред.). Динозаврия (2-е изд.). Беркли: Калифорнийский университет Press. стр.137 –150. ISBN 978-0-520-24209-8.
- ^ а б Зеленицкий, Д.К .; Therrien, F .; Эриксон, Г. М .; DeBuhr, C.L .; Кобаяши, Ю .; Eberth, D. A .; Хэдфилд, Ф. (2012). «Пернатые динозавры из Северной Америки дают представление о происхождении крыльев». Наука. 338 (6106): 510–514. Bibcode:2012Научный ... 338..510Z. Дои:10.1126 / наука.1225376. PMID 23112330. S2CID 2057698.
- ^ а б c Кобаяши, Ю .; Лю, Ж.-К. (2003). «Новый динозавр-орнитомимид со стайными привычками из позднего мела Китая». Acta Palaeontologica Polonica. 48 (2): 235–259.
- ^ а б Барретт П. М. (2005). «Рацион страусиных динозавров (Theropoda: Ornihomimosauria)». Палеонтология. 48 (2): 347–358. Дои:10.1111 / j.1475-4983.2005.00448.x.
- ^ а б c d Хурум, Дж. (2001). "Нижняя челюсть Галлимим буллатус". В Танке, Д. Х .; Карпентер, К .; Скрепник, М. В. (ред.). Мезозойская жизнь позвоночных. Блумингтон: Издательство Индианского университета. стр.34–41. ISBN 978-0253339072.
- ^ а б Норелл, М. А .; Makovicky, P.J .; Карри, П. Дж. (2001). «Клювы страусиных динозавров». Природа. 412 (6850): 873–874. Bibcode:2001Натура.412..873Н. Дои:10.1038/35091139. PMID 11528466. S2CID 4313779.
- ^ а б c Watanabe, A .; Eugenia Leone Gold, M .; Brusatte, S.L .; Benson, R. B.J .; Choiniere, J .; Дэвидсон, А .; Норелл, М. А .; Классенс, Л. (2015). «Вертебральная пневматичность у орнитомимозавра. Археорнитомим (Dinosauria: Theropoda), выявленные с помощью компьютерной томографии и переоценки осевой пневматичности орнитомимозаврии ». PLOS ONE. 10 (12): e0145168. Bibcode:2015PLoSO..1045168W. Дои:10.1371 / journal.pone.0145168. ЧВК 4684312. PMID 26682888.
- ^ Xu, L .; Кобаяши, Ю .; Lü, J .; Lee, Y.N .; Liu, Y .; Tanaka, K .; Чжан, X .; Jia, S .; Чжан, Дж. (2011). «Новый динозавр-орнитомимид с североамериканским родством из позднемеловой формации Цюпа в провинции Хэнань в Китае». Меловые исследования. 32 (2): 213–222. Дои:10.1016 / j.cretres.2010.12.004.
- ^ Hendrickx, C .; Hartman, S.A .; Матеус, О. (2015). «Обзор открытий и классификации нептичьих теропод». Журнал PalArch по палеонтологии позвоночных. 12 (1): 1–73.
- ^ Хольц, Т. Р. (1994). "Филогенетическое положение тираннозавров: значение для систематики теропод". Журнал палеонтологии. 68 (5): 1100–1117. Дои:10.1017 / S0022336000026706. JSTOR 1306180.
- ^ Хольц, Т. Р. (1998). «Новая филогения хищных динозавров». GAIA. 15: 5–61. CiteSeerX 10.1.1.211.3044. ISSN 0871-5424.
- ^ Naish, D .; Martill, D.M .; Фрей, Э. (2004). «Экология, систематика и биогеографические отношения динозавров, включая нового теропод, из формации Сантана (? Альб, ранний мел) Бразилии». Историческая биология. 16 (2–4): 57–70. CiteSeerX 10.1.1.394.9219. Дои:10.1080/08912960410001674200. S2CID 18592288.
- ^ Рассел, Д. А. (1972). «Страусиные динозавры из позднего мела Западной Канады». Канадский журнал наук о Земле. 9 (4): 375–402. Bibcode:1972CaJES ... 9..375R. Дои:10.1139 / e72-031.
- ^ Талборн, Р. А. (1982). «Скорости и походки динозавров». Палеогеография, палеоклиматология, палеоэкология. 38 (3–4): 227–256. Bibcode:1982ППП .... 38..227Т. Дои:10.1016/0031-0182(82)90005-0.
- ^ Nicholls, E.L .; Рассел, А. П. (1985). "Строение и функции грудного пояса и передних конечностей Struthiomimus altus (Theropoda: Ornithomimidae) ". Палеонтология. 28 (4): 64–677.
- ^ Мэдсен, Э. К. (2007). «Морфология клюва у современных птиц с последствиями для морфологии клюва у орнитомимид». Det Matematisk-Naturvitenskapelige Fakultet - Диссертация: 1–21.
- ^ Pawlicki, R .; Болехала, П. (1987). «Рентгеновский микроанализ ископаемых костей динозавров: возрастные различия в содержании кальция и фосфора в Галлимим буллатус кости ». Folia Histochemica et Cytobiologica. 25 (3–4): 241–244. PMID 3450541.
- ^ Pawlicki, R .; Болехала, П. (1991). «Рентгеновский микроанализ ископаемых костей динозавров: возрастные различия в содержании свинца, железа и магния». Folia Histochemica et Cytobiologica. 29 (2): 81–83. PMID 1804726.
- ^ Rensberger, J.M .; Ватабе, М. (2000). «Тонкая структура костей динозавров, птиц и млекопитающих». Природа. 406 (6796): 619–622. Bibcode:2000Натура.406..619р. Дои:10.1038/35020550. PMID 10949300. S2CID 4384549.
- ^ а б c Jerzykiewicz, T .; Рассел, Д. А. (1991). «Позднемезозойская стратиграфия и позвоночные животные Гобийской впадины». Меловые исследования. 12 (4): 345–377. Дои:10.1016/0195-6671(91)90015-5.
- ^ Салливан, Р. М. (2006). "Таксономический обзор Pachycephalosauridae (Dinosauria: Ornithischia)" (PDF). В Lucas, Spencer G .; Салливан, Роберт М. (ред.). Позднемеловые позвоночные из внутренних районов Запада. 35. Бюллетень Музея естественной истории и науки Нью-Мексико. С. 347–366.
- ^ Градштейн, Ф. М .; Ogg, J. G .; Смит, А. Г. (2005). Шкала геологического времени 2004. Кембридж: Издательство Кембриджского университета. стр.344 –371. Дои:10.1017 / CBO9780511536045. ISBN 978-0-521-78142-8.
- ^ Хольц, Т. Р. (2014). «Палеонтология: разгадана тайна ужасных рук». Природа. 515 (7526): 203–205. Bibcode:2014Натура.515..203H. Дои:10.1038 / природа13930. PMID 25337885. S2CID 205241353.
- ^ Новачек, М. (1996). Динозавры пылающих скал. Нью-Йорк: Якорь. п.133. ISBN 978-0-385-47775-8.
- ^ Jaworowski, Z .; Пенско, Дж. (1967). «Необычно радиоактивные ископаемые кости из Монголии». Природа. 214 (5084): 161–163. Bibcode:1967Натура.214..161J. Дои:10.1038 / 214161a0. S2CID 4148855.
- ^ Ефимов, М. Б. (1983). «Пересмотр ископаемых крокодилов Монголии». Совместная Советско-Монгольская Палеонтологическая Экспедиция Труды (на русском). 24: 76–96.
- ^ Hurum, J. H .; Сабат, К. (2003). "Гигантские динозавры-тероподы из Азии и Северной Америки: черепа Тарбозавр bataar и тиранозавр Рекс в сравнении" (PDF). Acta Palaeontologica Polonica. 48 (2): 188.
- ^ Хольц, Т. Р. (2004). «Тиранносавроидеа». В Weishampel, David B .; Додсон, Питер; Osmólska, Halszka (ред.). Динозаврия (2-е изд.). Беркли: Калифорнийский университет Press. п.124. ISBN 978-0-520-24209-8.
- ^ а б Shay, D .; Дункан, Дж. (1993). Создание Парка Юрского периода. Нью-Йорк: Boxtree Ltd., стр. 48–52, 135–136. ISBN 978-1-85283-774-7.
- ^ Крайтон, М. (1990). парк Юрского периода. Лондон: Random Century Group. стр.137–138. ISBN 978-0-394-58816-2.
- ^ Бэрд, Р. (1998). "Анимализация Парк Юрского периода динозавры: схемы блокбастеров и кросс-культурное познание в сцене угрозы ". Кино Журнал. 37 (4): 82–103. Дои:10.2307/1225728. JSTOR 1225728.
- ^ Принц, С. (1996). «Правдивая ложь: перцептивный реализм, цифровые изображения и теория кино». Film Quarterly (Представлена рукопись). 49 (3): 27–37. Дои:10.2307/1213468. HDL:10919/25811. JSTOR 1213468.
- ^ а б Монтанари, С. (2015). "Что мы узнали о динозаврах с тех пор, как парк Юрского периода вышел из". Forbes. Получено 7 мая 2018.
- ^ Кирби, Д. А. (2011). Лабораторные халаты в Голливуде: наука, ученые и кино. Массачусетс: MIT Press. стр.127 –129. ISBN 978-0262294867.
- ^ Кирби, Д. А. (2003). «Научные консультанты, игровые фильмы и научная практика». Социальные исследования науки. 33 (2): 231–268. Дои:10.1177/03063127030332015. JSTOR 3183078. S2CID 111227921.
- ^ Филлипс, И. (2015). "Как Мир юрского периода динозавры выглядели в реальной жизни ». Business Insider. Получено 3 декабря 2016.
- ^ Мартин, А. Дж. (2014). Динозавры без костей: судьбы динозавров, обнаруженные по ископаемым останкам. Лондон: Книги Пегаса. С. 66–67. ISBN 978-1605984995.
- ^ Ромилио, А. (2015). «Паническое бегство динозавров остановилось». Австралазийская наука. 36 (2): 24–27. ISSN 1442-679X.
- ^ Шапиро, А. Д. (2013). "Великая давка динозавров, которой никогда не было?". Новости BBC. Получено 7 мая 2018.
внешняя ссылка
 СМИ, связанные с Галлимим в Wikimedia Commons
СМИ, связанные с Галлимим в Wikimedia Commons
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||














| Авторитетный контроль |
|---|