Энантиорниты - Enantiornithes
| Энантиорниты | |
|---|---|
 | |
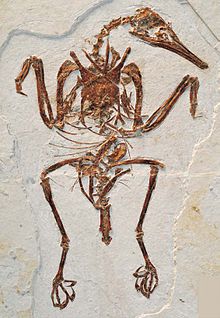
| Ископаемый образец бохайорнитид (Жуорнис хани ) | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Clade: | Динозаврия |
| Clade: | Заурисхия |
| Clade: | Тероподы |
| Clade: | Орнитоторасы |
| Clade: | †Энантиорниты Уокер, 1981 |
| Подгруппы | |
и посмотреть текст | |
Энантиорниты группа вымерших авиалайнеры («птицы» в широком смысле), наиболее многочисленная и разнообразная группа, известная по Мезозойская эра.[3][4][5] Почти у всех сохранились зубы и когтистые пальцы на каждом крыле, но в остальном они были очень похожи на современных птиц. Было названо более 80 видов энантиорнитей, но некоторые названия представляют собой только отдельные кости, поэтому вполне вероятно, что не все верны. Энантиорнитеи вымерли в Граница мела и палеогена, вместе с гесперорнитиды и все другие нептичьи динозавры.
Открытие и наименование
Первые обнаруженные энантиорнити были ошибочно отнесены к современным группам птиц. Например, первый известный энантиорнитей, Gobipteryx minuta изначально считалась палеогнат относится к страусы и тинамо.[6] Enantiornitheans были впервые признаны отдельной ветвью или «подклассом» птиц Сирил А. Уокер в 1981 году. Уокер сделал это открытие на основании некоторых частичных останков позднего Меловой период из того, что сейчас Аргентина, который он отнес к новому роду, Энантиорнис, давая название всей группе. С 1990-х годов было обнаружено гораздо больше полных энантиорнитов, и было установлено, что несколько ранее описанных «птиц» (например, Иберомесорнис, Катайорнис, и Sinornis ) тоже были энантиорнитами.
Название «Энантиорнитес» означает «противоположные птицы», от Древнегреческий энантиос (ἐνάντιος) "напротив" + орниты (ὄρνιθες) "птицы". Название было придумано Кирилл Александр Уокер в его знаменательном документе, который основал группу.[7] В своей статье Уокер объяснил, что он имел в виду под «противоположностью»:
Возможно, наиболее фундаментальное и характерное отличие энантиорнитов от всех других птиц заключается в характере сочленения между ними. лопатка [...] и коракоидный, где «нормальное» условие полностью изменено.[7]
Это относится к анатомической особенности - сочленению плечо кости - с вогнутым и выпуклым суставом, противоположным суставу современных птиц. В частности, у энантиорнитеев грань, в которой лопатка (лопатка) встречает коракоидный (первичная кость плечевого пояса у позвоночных животных, кроме млекопитающих) - это выпуклый ручка и соответствующая точка на лопатке вогнутый и тарельчатые. У современных птиц суставы суставов противоположны.[8]
Уокеру не совсем ясно, почему он дал это имя в разделе этимологии своей статьи, и эта двусмысленность привела к некоторой путанице среди более поздних исследователей. Например, Алан Федучча заявлено в 1996 году:
Птицы названы так потому, что среди многих отличительных черт есть уникальное образование трехкостного канала, а плюсневые кости срастаются проксимально и дистально, в отличие от современных птиц.[9]
Точка зрения Федуччи о предплюсна (сочетание верхней части стопы и лодыжки) является правильным, но Уокер не использовал это рассуждение в своей исходной статье. Уокер никогда не описывал слияние tarsometatarsus как противоположное, а как «только частичное». Кроме того, нет уверенности в том, что у энантиорнитей были трехкостные каналы, поскольку ни одно ископаемое не сохранило эту особенность.[3]
Группу энантиорнитов часто называют «энантиорнитинами». Однако некоторые ученые отметили, что это неверно, поскольку следуя стандартным правилам формирования названий групп животных, подразумевается ссылка только на подсемейство. Энантиорнитины. Следуя соглашениям об именах, используемым для современных птиц, а также для вымерших групп, было указано, что правильный термин - «энантиорниты».[10]
Происхождение и диапазон
Энантиорнитеи были обнаружены на всех континентах, кроме Антарктида. Ископаемые останки, относящиеся к этой группе, исключительно Меловой по возрасту, и считается, что энантиорнитеи вымерли одновременно с их нептичными динозавр родственники. Самые ранние известные энантиорниты происходят из Раннемеловой период из Испания (например. Noguerornis ) и Китай (например. Протоптерикс ) и последний из Поздний мел Северной и Южной Америки (например, Авизавр и Энантиорнис ). Широкое распространение этой группы предполагает, что, по крайней мере, некоторые энантиорнити были способны пересекать океаны своими собственными силами; они являются первой известной птичьей линией глобального распространения.
Описание

Многие окаменелости энантиорнитей очень фрагментарны, а некоторые виды известны только по куску единственной кости. Почти все образцы полные, с полным сочленением и сохранением мягких тканей известны из Лас Хойас в Куэнка, Испания и Группа Jehol в Ляонин (Китай ). Окаменелости энантиорнифеев были найдены как во внутренних, так и в морских отложениях, что позволяет предположить, что они были экологически разнообразной группой. К энантиорнитам относились кулики, пловцы, зерноядные, насекомоядные, рыболовы и хищники. Подавляющее большинство видов энантиорнитей были небольшими, размером с воробей и скворец,[11] в то время как самые крупные члены этой клады Pengornis houi,[12] Xiangornis shenmi[13] и Жуорнис хани.[11] По крайней мере, несколько гораздо более крупных видов могли существовать, в том числе виды, потенциально размером с крана, известные только по следам в Формация Эумералла (и, возможно, также представлен в Формация Вонтхагги одним фуркула ), который может принадлежать энантиорнитею.[14] Один таксон, Mirarce, описывается как похожий по размеру на современный индейки,[15] в то время как предыдущие «крупнейшие энантиорниты» описываются как черный дрозд размер.[12]
Необычные останки энантиорнифеев сохранились также в Бирманский янтарь отложения датируются 99 миллионами лет назад. Эти останки являются одними из наиболее хорошо сохранившихся среди мезозойских динозавров. Первыми обнаруженными останками энантиорнифея в янтарной оболочке были два крыла (см. Ниже), описанные в 2016 году.[16] Почти все тело вылупившегося энантиорнитея было описано в 2017 г.[17] еще один детеныш был описан в 2018 году.[18] В 2019 году была описана практически полная стопа вместе с крылом.[19] В 2020 году было описано крыло крупного таксона.[20]
Череп

Учитывая широкий диапазон сред обитания и рациона, черепа энантиорнитей значительно различались между видами. Черепа энантиорнитей сочетают в себе уникальный набор примитивных и сложных особенностей. Как и в более примитивных авиалайнах вроде Археоптерикс, они сохранили несколько отдельных черепных костей, мелких предчелюстные кости (кости кончика морды) и у большинства видов были зубастые челюсти, а не беззубые клювы. Только несколько видов, например Gobipteryx minuta, были полностью беззубыми и имели клювы. У них также были простые квадратные кости, полная полоса, разделяющая каждую орбита (глазок) от каждого анторбитальное окно, и зубные кости (основные зубчатые кости нижней челюсти) без раздвоенных задних вершин. А чешуйчатая кость сохранился у неопределенного молодняка, а заглазничный сохраняется в Шенкиорнис и Pengornis. У современных птиц эти кости ассимилируются в черепной коробке. Некоторые энантиорнитеи могли иметь височные отверстия (отверстия в боковой части головы) слились в орбиты, как у современных птиц, из-за того, что задние глазницы либо отсутствуют, либо недостаточно длинные, чтобы разделить отверстия.[21] А четырехъядерная кость, который у современных птиц срастается с скуловой грудью, сохранился в Птеригорнис.[22] Наличие этих примитивных черт черепа сделало энантиорнитей способными лишь к ограниченному черепной кинезис (способность двигать челюстью независимо от черепа).[23]
Крыло

Как очень большая группа птиц, энантиорнитеи демонстрировали большое разнообразие различных строений тела, основанных на различиях в экологии и питании, что отражалось в одинаковом разнообразии форм крыльев, во многих параллельных адаптациях к разному образу жизни, наблюдаемых у современных птиц. В целом крылья энантиорнитеев были более развитыми по сравнению с более примитивными птицами, такими как Археоптерикс, и отображал некоторые особенности, связанные с полетом, аналогичные тем, которые встречаются в линии, ведущей к современным птицам, Орнитуроморфа. Хотя у большинства энантиорнитов были когти, по крайней мере, на некоторых пальцах, у многих видов были укороченные руки, очень подвижный плечевой сустав и пропорциональные изменения в костях крыльев, как у современных птиц. Как и современные птицы, энантиорнитеи имели Alulas, или «ублюдочные крылья», небольшие направленные вперед перья на первом пальце, которые обеспечивали большую маневренность в воздухе и помогали при точном приземлении.[24]
Несколько крыльев с сохранившимися перьями были найдены сохранившимися в Бирманский янтарь. Это первые полные Мезозойский динозавр таким образом сохранились (несколько изолированных перьев известны иначе, не относящиеся ни к какому виду), и это одна из самых прекрасно сохранившихся окаменелостей динозавров.[25] Сохранившиеся крылья демонстрируют вариации пигмента пера и доказывают, что у энантиорнитей были полностью современные перья, включая зазубрины, зазубрины и крючки, а также современное расположение перьев крыльев, включая длинные маховые перья, короткие кроющие, большой алула и подшерсток из пуха.[16]
Одна окаменелость энантиорнифеев показывает похожие на крылья пучки перьев на ногах, похожие на Археоптерикс. Перья на ногах также напоминают четырехкрылых динозавров. Микрораптор однако у энантиорнитей в отличие от перьев они короче, более неорганизованы (не образуют четко крыла) и простираются только до щиколотки, а не вдоль стопы.[26]
Хвост
Кларк и другие. (2006) исследовали все известные тогда окаменелости энантиорнитов и пришли к выводу, что ни у одного из них не сохранились рулевые перья, которые образуют веер, создающий подъемную силу, как у современных птиц. Они обнаружили, что все авиалайнеры за пределами Euornithes (клада, которую они назвали Орнитура ) с сохранившимися рулевыми перьями имели только короткие кроющие или удлиненные парные хвостовые оперения. Они предположили, что разработка пигостиль у энантиорнитов, должно быть, это результат укорочения хвоста, а не развитие современной анатомии хвостовых перьев. Эти ученые предположили, что поклонник рулевых перьев и связанной с ними мускулатуры необходим для управления ими, известной как прямоугольная колба, развивались вместе с коротким треугольным пигостилем, как у современных птиц, а не с длинными пигостилями в форме стержня или кинжала у более примитивных птичьих особей, таких как энантиорнитеи. Вместо веера из перьев у большинства энантиорнитей была пара длинных специализированных перьев, похожих на перья вымерших. Конфуциусорнис и некоторые райские птицы.[27]
Однако дальнейшие открытия показали, что, по крайней мере, у примитивных энантиорнитов анатомия хвоста была более сложной, чем считалось ранее. Один энантиорнитей, Shanweiniao, изначально было интерпретировано как наличие по крайней мере четырех длинных рулевых перьев, которые перекрывают друг друга[28] и, возможно, образовал поверхность, создающую подъемную силу, подобную хвостовым веерам еворнитей,[29] хотя более позднее исследование показывает, что Shanweiniao с большей вероятностью имел рахис - преобладают рулевые перья, похожие на перья, присутствующие в Парапротоптерикс.[30] Чиаппевис, примитивный Pengornithid энантиорнитей, имел веер из хвостовых перьев, как у более примитивных птиц, таких как Сапеорнис, что позволяет предположить, что это могло быть наследственным состоянием, поскольку булавочные перья были особенностью, которая развивалась несколько раз у ранних птиц для демонстрации.[30] Другой энантиорнитей, Фейтианиус, также имел сложный веер из рулевых перьев. Что еще более важно, мягкая ткань, сохранившаяся вокруг хвоста, была интерпретирована как остатки прямоугольной луковицы, предполагая, что эта особенность на самом деле не ограничивалась видами с современными пигостилями, но могла развиться намного раньше, чем считалось ранее, и присутствовала во многих энантиорниты.[31] По крайней мере, один род энантиорнитеев, Cruralispennia, имел современный пигостиль, но не имел хвостового веера.[32]
Биология
Рацион питания
Учитывая большое разнообразие форм черепа у энантиорнитов, в группе должно было присутствовать множество различных диетических специализаций. Некоторые, как Шенкиорнис, имел большие крепкие челюсти, подходящие для поедания беспозвоночных с твердым панцирем. В лонгиптеригиды, морды были длинными и тонкими, с зубами, ограниченными кончиками челюстей, и они, вероятно, принадлежали к иловым зондам (мелкозубые виды) и рыболовам (зубастые виды). Короткие тупые зубы Pengornis вероятно, использовались для питания мягкотелых членистоногих.[21] Сильно загнутые когти бохайорнитиды Предполагают, что они были хищниками мелких и средних позвоночных, но их крепкие зубы вместо этого предполагают диету из животных с твердым панцирем.[1]
Некоторые образцы сохраняют фактическое содержимое желудка. К сожалению, ни один из них не сохраняет череп, поэтому прямая корреляция между их известной диетой и формой морды / зубов не может быть установлена. Эоалулавис обнаружили останки экзоскелеты из водных ракообразные сохраняется в пищеварительном тракте,[33] и Энантиофеникс сохранились частицы янтаря среди окаменелых костей, что позволяет предположить, что это животное питалось древесным соком, как и современные сапсосы и другие птицы. Сок бы окаменел и превратился в янтарь.[34] Однако совсем недавно было высказано предположение, что сок перемещался после смерти, следовательно, не представлял истинного содержимого желудка. В сочетании с предполагаемой рыбой пеллеты из Piscivorenantiornis оказалось рыбьими экскрементами, странное содержимое желудка некоторых видов оказалось яичники и предполагаемые гастролиты Бохайорнис будучи случайными минеральными осадками, только Эоалулавис отображает фактическое содержимое желудка.[35]
Исследование паравианской пищеварительной системы показывает, что известные энантиорниты не имели зоба и желудка, не использовали гастролиты и не выбрасывали гранулы. Это считается противоречащим большому разнообразию диет, которые подразумевают их разные зубы и формы черепа.[36] хотя некоторые современные птицы лишились желудка и полагаются исключительно на сильную желудочную кислоту.[37]
Хищничество
Ископаемое из Испании, о котором сообщает Санс. и другие. в 2001 г. включены останки четырех вылупившихся скелетов энантиорнитов трех разных видов. Они практически закончены, очень тесно связаны и показывают изъязвления костей на поверхности, что указывает на частичное пищеварение. Авторы пришли к выводу, что эта ассоциация была отрыгнутым осадком, и, исходя из деталей пищеварения и размера, вылупившиеся птенцы были проглочены целиком. птерозавр или маленький теропод динозавр. Это было первое доказательство того, что Мезозойский птичьи были хищными животными, и что некоторые мезозойские пан-птицы срыгнули гранулы, как сегодня совы.[38]
История жизни

Известные окаменелости энантиорнитей включают: яйца,[39][40] эмбрионы,[41] и детеныши.[42] Сообщалось, что эмбрион энантиорнифея, все еще свернувшийся в яйце, Формация Исянь.[43] Молодые особи могут быть идентифицированы по совокупности факторов: грубая текстура их кончиков костей, указывающая на части, которые на момент смерти все еще состояли из хряща, относительно небольшие грудные кости, большие черепа и глаза, а также кости, которые еще не срослись друг с другом. .[44] Некоторым вылупившимся особям были даны официальные имена, включая "Liaoxiornis delicatus "; тем не мение, Луис Чиаппе и его коллеги считали, что наименование новых видов на основе молоди вредно для изучения энантиорнитов, потому что практически невозможно определить, к какому взрослому виду принадлежит данный молодой особь, что делает любой вид с голотипом вылупившихся особей номен дубиум.[44]
Вместе с вылупившимися особями монгольской Гобиптерикс[45] и Gobipipus,[46][47] Эти находки демонстрируют, что у детенышей энантиорнитов было окостенение скелета, хорошо развитые перья крыльев и большой мозг, которые коррелируют с ранний или же суперпрофессиональный закономерности развития современных птиц. Другими словами, энантиорнити, вероятно, вылупились из яйца, уже хорошо развитого и готового бегать, добывать корм и, возможно, даже летать в возрасте всего нескольких дней.[44]
Для определения скорости роста этих животных был проведен анализ гистологии энантиорнитной кости. Исследование 2006 г. Конкорнис кости показали образец роста, отличный от современных птиц; хотя рост был быстрым в течение нескольких недель после вылупления, вероятно, до оперение, этот мелкий вид долгое время не достигал взрослых размеров, вероятно, несколько лет.[48] Все другие исследования подтвердили мнение о том, что рост до взрослого размера был медленным, как и у современных ранних птиц (в отличие от альтриальный птицы, которые, как известно, быстро достигают взрослых размеров).[33] Исследования скорости роста костей у различных энантиорнитов показали, что более мелкие виды имеют тенденцию расти быстрее, чем более крупные, что противоположно паттерну, наблюдаемому у более примитивных видов, таких как Jeholornis и у нептичьих динозавров.[49] Некоторые анализы интерпретировали гистологию кости, чтобы указать на то, что энантиорниты, возможно, не имели полностью птичьего эндотермия, вместо промежуточного скорость метаболизма.[50]
Доказательства колониального гнездования были обнаружены у энантиорнитей в отложениях Поздний мел (Маастрихтский ) из Румыния.[51] Данные из мест гнездования показывают, что энантиорнити закапывали свои яйца, как современные мегаподы, что согласуется с их предполагаемой суперпрофессиональной адаптацией.[52]
Исследование молодых перьев энантиорнитей, проведенное в 2020 году, еще больше подчеркивает онтологическое сходство с современными мегаподами, но предостерегает от некоторых различий, таких как древесная природа большинства энантиорнитов в отличие от наземного образа жизни мегапод.[53]
Полет
Поскольку многие энантиорнити не имели сложных хвостов и обладали совершенно иной анатомией крыльев по сравнению с современными птицами, они стали предметом нескольких исследований, проверяющих их летные способности.
Традиционно их считали летчиками невысокого качества из-за того, что анатомия плечевого пояса считается более примитивной и не может поддерживать наземный пусковой механизм.[54] а также из-за отсутствия прямоугольники у многих видов.[27][29][55]
Однако несколько исследований показали, что они были эффективными летчиками, как и современные птицы, обладая такой же сложной нервной системой и связками перьев крыльев. Кроме того, отсутствие сложного хвоста, по-видимому, не очень важно для птичий полет в целом - некоторые вымершие птицы любят литорниды также отсутствовали сложные рулевые перья, но были хорошими летчиками,[56] и они, похоже, были способны запускать с земли.[57]
Из-за разницы в анатомии грудины и плечевого пояса многие энантиорнити использовали стиль полета, отличный от любого современного вида птиц.[требуется разъяснение ], хотя присутствовали и более типичные стили полета.[58]
По меньшей мере Эльсорнис кажется, стало вторично нелетающий.[59]
Классификация
Некоторые исследователи классифицируют энантиорнитеев, наряду с настоящими птицами, в класс Авес. Другие используют более ограничительный группа короны определение Aves (которое включает только неорниты, анатомически современные птицы) и поместите энантиорнитов в более инклюзивную группу Avialae. Энантиорнити были более развитыми, чем Археоптерикс, Конфуциусорнис, и Сапеорнис, но в некоторых отношениях они были более примитивными, чем современные птицы, возможно, следуя промежуточному эволюционному пути.
Консенсус научного анализа показывает, что энантиорниты - одна из двух основных групп внутри большой группы. Орнитоторасы. Другая группа орнитоторацинов - это Euornithes или же Орнитуроморфа, который включает всех живых птиц как подмножество. Это означает, что энантиорнити были успешной ветвью эволюции птиц, но диверсифицировавшейся полностью отдельно от линии, ведущей к современным птицам.[3] Однако одно исследование показало, что общая анатомия грудины была получена независимо, и такую взаимосвязь необходимо пересмотреть.[60]
Классификация и таксономия энантиорниев исторически осложнялась рядом факторов. В 2010 году палеонтологи Джингмай О'Коннор и Гарет Дайк обрисовали ряд критических замечаний в отношении преобладающей практики ученых, неспособных описать многие образцы достаточно подробно, чтобы другие могли их тщательно оценить. Некоторые виды были описаны на основе образцов, которые хранятся в частных коллекциях, что делает невозможным дальнейшее изучение или обзор предыдущих находок. Поскольку другие ученые часто не могут лично изучить каждый образец, учитывая всемирное распространение энантиорнитов, и из-за большого количества неинформативных описаний, которые были опубликованы на возможно важных образцах, многие из этих образцов становятся «функциональными». Номина Дубия ".[61] Более того, многие виды были названы на основе чрезвычайно фрагментарных образцов, которые не были бы очень информативными с научной точки зрения, даже если бы они были описаны достаточно. Более трети всех названных видов энантиорнитей основаны только на фрагменте одной кости. О'Коннор и Дайк утверждали, что, хотя эти образцы могут помочь расширить знания о временном промежутке или географическом ареале энантиорнитов, и важно их описать, наименование таких образцов «неоправданно».[61]
Отношения
Enantiornithes - это родственная группа Euornithes, и вместе они образуют клады называется Орнитоторасы (хотя см. выше). Наиболее филогенетический исследования выявили энантиорнитов как монофилетическую группу, отличную от современных птиц и их ближайших родственников. Филогенетический анализ 2002 г., проведенный Кларком и Нореллом, тем не менее, снизил количество энантиорнитиев. аутапоморфии всего четыре.[62]
Систематика энантиорнитей весьма условна и, как известно, трудна для изучения из-за того, что энантиорниты, как правило, чрезвычайно гомопластический, или очень похожи друг на друга по большинству черт своего скелета из-за конвергентной эволюции, а не общего происхождения.[30] К настоящему времени кажется вполне определенным, что внутри энантиорнитов были подразделения, возможно, включающие некоторые второстепенные базальные линии в дополнение к более продвинутым Euenantiornithes. Детали взаимоотношений всех этих линий, а также достоверность большинства из них, оспариваются, хотя Avisauridae, например, кажется, вероятно, составляют действительную группу. Филогенетические систематики до сих пор очень неохотно предлагали разграничения энантиорнитовых клад.[63]
Одно такое разграничение назвало Euenantiornithes, был определен Chiappe (2002) как включающий все виды, близкие к Sinornis чем Иберомесорнис. Потому что Иберомесорнис часто обнаруживается, что это самый примитивный или базальный энантиорнит, Euenantiornithes может быть чрезвычайно инклюзивной группой, состоящей из всех энантиорнитов, за исключением Иберомесорнис сам. Несмотря на соответствие филогенетическая номенклатура, это определение Euenantiornithes подверглось резкой критике со стороны некоторых исследователей, таких как Пол Серено, который назвал это "неопределенным клады [...] хороший пример плохого выбора в филогенетическом определении ».[63]
В кладограмма ниже было найдено анализом Ванга и другие. в 2015 году, обновлено на основе предыдущего набора данных, созданного Джингмай О'Коннор.[22]
| Орнитоторасы |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Список родов
Incertae sedis
Таксономию энантиорнитов трудно оценить, и в результате филогенетический анализ последовательно обнаруживает несколько клад внутри группы. Большинство энантиорнитов не входят в какое-либо конкретное семейство и перечислены здесь. Многие из них считались евэнантиорнитами, хотя разногласия, стоящие за этим названием, означают, что оно не используется постоянно в исследованиях энантиорнитей.
| Имя | Год | Формирование | Место расположения | Примечания | Изображений |
|---|---|---|---|---|---|
| Абаворнис | 1998 | Биссектская свита (Поздний мел, Туронский к Коньяк ) | Один из многих фрагментарных энантиорниев Биссекты, известный только с коракоиды | ||
| Алетоалаорнис | 2007 | Формация Цзюфотан (Раннемеловой период, Аптян ) | Малоизвестный | ||
| Alexornis | 1976 | Формация Ла Бокана Роха (Поздний мел, Кампанский ) | Один из первых известных энантиорнитов. Когда-то считался древним родственником ролики и дятлы | ||
| Avimaia | 2019 | Формация Сягоу (Поздний мел, Аптян ) | Один представитель этого рода умер с невыложенным яйцом в теле. |  | |
| Bauxitornis | 2010 | Формация Чехбанья (Поздний мел, Сантон ) | Фрагментарно, но уникально по строению предплюсны. |  | |
| Катенолеймус | 1998 | Биссектская свита (Поздний мел, Туронский к Коньяк ) | Один из многих фрагментарных энантиорниев Bissekty, известный только коракоидный | ||
| Катайорнис | 1992 | Формация Цзюфотан (Раннемеловой период, Аптян ) | Один из первых описанных энантиорнитов Jehol biota. Известны по многим видам, хотя некоторые теперь относятся к своим родам. Возможно, имел внешний вид и образ жизни, похожий на питта |  | |
| Конкорнис | 1992 | Лас Хойас (Раннемеловой период, Барремский ) | Один из самых полных энантиорнитов Лас-Хояс |  | |
| Кратоавис[64] | 2015 | Формация Сантана (Раннемеловой период, Аптян ) | Очень хорошо сохранившийся южноамериканский член группы с лентами на хвосте. | ||
| Cruralispennia[32] | 2017 | Формация Хуацзин (Раннемеловой период, Готеривский ) | Обладает необычным орнитуроморфоподобным пигостилем и кистевидными перьями на бедрах. Один из старейших энантиорнитеев |  | |
| Куспирострисорнис | 1997 | Формация Цзюфотан (Раннемеловой период, Аптян ) | Первоначально ошибочно считалось, что обладал заостренным клювом. | ||
| Далингхеорнис | 2006 | Формация Исянь (Раннемеловой период, Аптян ) | Был хорошо приспособлен для лазания благодаря своему гетеродактильные ноги, как у трогон | ||
| Дуньхуанья[65] | 2015 | Формация Сягоу (Раннемеловой период, Аптян ) | Энантиорнитей из бассейна Чангма, области, в которой необычно преобладают орнитуроморфы. | ||
| Эльбреторнис | 2009 | Формация Лечо (Поздний мел, Маастрихтский ) | Известен только по костям крыльев. Может быть синонимом других энантиорнитей формации Lecho | ||
| Elektorornis | 2019 | Бирманский янтарь (Поздний мел, Сеноманский ) | Известен по стопе, сохранившейся в янтаре, с удлиненным средним пальцем. | ||
| Эльсорнис | 2007 | Джадочтинская свита (Поздний мел, Кампанский ) | Его скелет, хотя и неполный, сохраняет трехмерную сохранность. Возможно, нелетающий из-за размеров крыльев | ||
| Энантиорнис | 1981 | Формация Лечо (Поздний мел, Маастрихтский ) | Хотя этот род известен только по нескольким костям, он является тезкой Enantiornithes. Он также был одним из крупнейших и последних представителей группы до их исчезновения. | ||
| Эоалулавис | 1996 | Лас Хойас (Раннемеловой период, Барремский ) | Сохраняет перья, в том числе Alula, специальный тип пера, который контролирует поток воздуха над крылом. | ||
| Эокатайорнис | 2002 | Формация Цзюфотан (Раннемеловой период, Аптян ) | Когда-то считался базальным близким родственником Катайорнис, хотя сейчас считается более отдаленным | ||
| Эоэнантиорнис | 1999 | Формация Исянь (Раннемеловой период, Аптян ) | Хорошо сохранились, но несовместимы в филогенетическом расположении |  | |
| Евгенавис | 2014 | Илекская формация (Раннемеловой период, Барремский ) | Известен только по tarsometatarsus, который имеет некоторые общие черты с таковыми у энантиорнитей. | ||
| Explorornis | 1998 | Биссектская свита (Поздний мел, Туронский к Коньяк ) | Один из многих фрагментарных энантиорниев Биссекты, известный только с коракоиды | ||
| Falcatakely | 2020 | Формация Маэварано (Поздний мел, Маастрихтский ) | Развил массивную морду только с одним зубом, несмотря на сохранение «примитивного» черепа в отличие от современных птиц. | ||
| Фейтианиус[31] | 2015 | Формация Сягоу (Раннемеловой период, Аптян ) | Обладал сложным набором рулевых перьев, в отличие от парных лентообразных перьев большинства энантиорнитов. | ||
| Flexomornis | 2010 | Формация Вудбайн (Поздний мел, Сеноманский ) | Один из старейших обнаруженных птиц в Северной Америке, хотя известен только по фрагментарным останкам. | ||
| Fortunguavis[66] | 2014 | Формация Цзюфотан (Раннемеловой период, Аптян ) | Имел крепкие кости, в том числе лапы и когти, которые могли быть приспособлены для лазания по деревьям. | ||
| Grabauornis[67] | 2015 | Формация Исянь (Раннемеловой период, Барремский ) | Пропорции крыльев этого энантиорнифея, а также наличие alula позволяют предположить, что это был хороший летчик. | ||
| Грасилорнис | 2011 | Формация Цзюфотан (Раннемеловой период, Аптян ) | Возможный родственник Катайорнис с характерно тонкими костями | ||
| Гурилина | 1999 | Формация Немегт (Поздний мел, Маастрихтский ) | Малоизвестный энантиорнитей, но, очевидно, крупный и поздно выживший член группы. | ||
| Голландия Люцерия[68] | 2010 | Формация Барун Гойот (Поздний мел, Кампанский ) | Первоначально идентифицированный как орнитуроморф, но затем переинтерпретированный как энантиорнит, тесно связанный с Лектавис.[69] | ||
| Холботия[70] | 2015 | Андайхудагская свита (Раннемеловой период, Аптян ) | Считается небольшим птерозавр с момента его открытия в 1977 году до получения официального описания в 2015 году. Обладал уникальными шейными позвонками и примитивным небом. | ||
| Houornis | 1997 | Формация Цзюфотан (Раннемеловой период, Аптян ) | Когда-то считалось сомнительным или разновидностью Катайорнис, хотя исследование 2015 г. считало его допустимым родом[71] | ||
| Хуошанорнис | 2010 | Формация Цзюфотан (Раннемеловой период, Аптян ) | Возможно, он был очень маневренным летчиком из-за строения кисти и грудины. | ||
| Иберомесорнис | 1992 | Лас Хойас (Раннемеловой период, Барремский ) | Один из первых энантиорнитей, известных по приличным останкам. Также один из самых старых и примитивных членов группы. |  | |
| Инколорнис | 1998 | Биссектская свита (Поздний мел, Туронский к Коньяк ) | Один из многих фрагментарных энантиорниев Биссекты, известный только с коракоиды. Один вид когда-то считался принадлежащим к Энантиорнис | ||
| Юнорнис[72] | 2017 | Формация Исянь (Раннемеловой период, Аптян ) | Настолько хорошо сохранился, что рисунок его полета можно было воссоздать, используя пропорции его перьев и крыльев. |  | |
| Кызылкумавис | 1984 | Биссектская свита (Поздний мел, Туронский к Коньяк ) | Один из многих фрагментарных энантиорниев Биссекты, известный только плечевая кость фрагмент | ||
| Ларгиросторнис | 1997 | Формация Цзюфотан (Раннемеловой период, Аптян ) | Возможно связано с Куспирострисорнис или синоним Катайорнис | ||
| Лектавис | 1993 | Формация Лечо (Поздний мел, Маастрихтский ) | Крупный и длинноногий представитель группы, пропорционально похожий на современных куликов. | ||
| Lenesornis | 1996 | Биссектская свита (Поздний мел, Туронский к Коньяк ) | Один из многих фрагментарных энантиорниев Bissekty, известный только синсакрум фрагмент. Первоначально считалось принадлежащим Ихтиорнис | ||
| Ляонингорнис | 1996 | Формация Исянь (Раннемеловой период, Аптян ) | Первоначально считался орнитураном, но теперь считается родственником Эоалулавис | ||
| Лонгченгорнис | 1997 | Формация Цзюфотан (Раннемеловой период, Аптян ) | Может быть синонимом Катайорнис | ||
| Мартинавис | 2007 | Формация Grès à Reptiles, Формация Лечо (Поздний мел, Маастрихтский ) | Хотя этот род известен только по плечевым суставам, он был большим и обитал в широком диапазоне. | ||
| Микроэнантиорнис | 2017 | Формация Цзюфотан (Раннемеловой период, Аптян ) | Небольшой член группы, обладавший несколькими примитивными и производными чертами по сравнению с другими энантиорнитами. | ||
| Моноэнантиорнис[73] | 2016 | Формация Исянь (Раннемеловой период, Аптян ) | Известен по молодому экземпляру, который показывает, как у энантиорнитей развивались различные черты с возрастом. | ||
| Нанантий | 1986 | Формация Тулебук (Раннемеловой период, Альбианский ) | Фрагментарно, но, возможно, это была морская птица, потому что останки этого рода были найдены как ихтиозавр содержимое кишечника | ||
| Noguerornis | 1989 | Эль-Монсек (Раннемеловой период, Барремский ) | Сохраняет впечатления от пропатагиум, кожный лоскут на плече, который является частью крыла | ||
| Ориенантий | 2018 | Формация Хуацзин (Раннемеловой период, Готеривский ) | Многие детали мягких тканей особей этого рода были обнаружены УФ-светом. | ||
| Отогорнис | 1993 | Формация Йиджинхолуо (Раннемеловой период ) | Малоизвестный | ||
| Парапротоптерикс | 2007 | Цяотоу член Формация Хуацзин (Раннемеловой период, Аптян ?) | По-видимому, у них было четыре ленточных хвостовых пера вместо двух, как у большинства энантиорнитей. | ||
| Парвавис[74] | 2014 | Формация Цзяндихэ (Поздний мел, Туронский к Сантон ) | Маленький, но полностью созревший к моменту смерти. Единственный описанный китайский энантиорнит, датируемый поздним меловым периодом. | ||
| Piscivorenantiornis[75] | 2017 | Формация Цзюфотан (Раннемеловой период, Аптян ) | Известен по разобранному скелету, сохранившемуся над частью содержимого желудка, состоящего из рыбьих костей, который, возможно, был его последней едой. | ||
| Протоптерикс | 2000 | Формация Хуацзин (Раннемеловой период, Готеривский ) | Один из самых старых и примитивных членов группы. |  | |
| Птеригорнис[22] | 2016 | Формация Цзюфотан (Раннемеловой период, Аптян ) | Один разрозненный скелет из этого рода имеет хорошо сохранившиеся кости черепа, в том числе квадратично-скелетную кость. | ||
| Qiliania | 2011 | Формация Сягоу (Раннемеловой период, Аптян ) | Некоторые из останков этого рода включают хорошо сохранившиеся задние конечности. названия видов, Q. graffini, назван в честь Грег Граффин из группы Плохая религия | ||
| Сазавис | 1989 | Биссектская свита (Поздний мел, Туронский к Коньяк ) | Один из многих фрагментарных энантиорниев Bissekty, известный только большеберцовая кость (кость голени) | ||
| Шанъян | 2019 | Формация Цзюфотан (Раннемеловой период, Аптян ) | Необычно то, что предчелюстные кости этого рода срослись. | ||
| Sinornis | 1992 | Формация Цзюфотан (Раннемеловой период, Аптян ) | Один из первых описанных энантиорнитов Jehol biota. Похожий на Катайорнис но обычно считается отличным |  | |
| Xiangornis | 2012 | Формация Цзюфотан (Раннемеловой период, Аптян ) | Рука этого рода была похожа на руку орнитуроморфов, вероятно, через конвергентная эволюция. Большой член группы | ||
| Юаньцзяваорнис[76] | 2015 | Формация Цзюфотан (Раннемеловой период, Аптян ) | Один из крупнейших энантиорнитов, известных по приличным останкам | ||
| Юнгаволукрис | 1993 | Формация Лечо (Поздний мел, Маастрихтский ) | Имел большой и необычно широкий предплюсневой минет (лодыжка) | 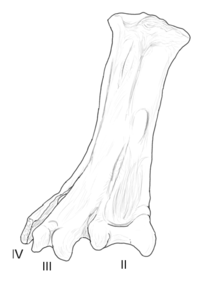 |
Longipterygidae
В Longipterygidae был семейством длиннорылых энантиорнитей раннего мелового периода, с зубами только на кончиках морды. Обычно они считаются базальными членами группы.[28]
| Имя | Год | Формирование | Место расположения | Примечания | Изображений |
|---|---|---|---|---|---|
| Болуохия | 1995 | Формация Цзюфотан (Раннемеловой период, Аптян ) | Первоначально ошибочно считалось, что у него был крючковатый клюв. | ||
| Camptodontornis | 2010 | Формация Цзюфотан (Раннемеловой период, Аптян ) | Первоначально назывался Camptodontus, хотя это название рода занято жук | ||
| Дапингфангорнис | 2006 | Формация Цзюфотан (Раннемеловой период, Аптян ) | На лбу могла быть шиповидная структура | ||
| Лонгиптерикс | 2001 | Формация Цзюфотан (Раннемеловой период, Аптян ) | Самый распространенный и известный член семьи |  | |
| Longirostravis | 2004 | Формация Исянь (Раннемеловой период, Аптян ) | Как и другие лонгиптеригиды, у него была тонкая морда, которую, возможно, использовали для поиска беспозвоночных в грязи или коре. | ||
| Rapaxavis | 2009 | Формация Цзюфотан (Раннемеловой период, Аптян ) | Специализируется на насесте благодаря строению лап. | ||
| Shanweiniao | 2009 | Формация Исянь (Раннемеловой период, Аптян ) | Приобретены несколько рулевых перьев, которые могли создавать подъемную силу, как у современных птиц. |  | |
| Shengjingornis | 2012 | Формация Цзюфотан (Раннемеловой период, Аптян ) | Большой член семьи |
Pengornithidae
В Pengornithidae была семьей больших ранних энантиорнитеев. У них были многочисленные мелкие зубы и многочисленные примитивные черты, которые утрачены у большинства других энантиорнитов.[2] Некоторые исследования утверждают, что они могут быть вовсе не энантиорнитами, а скорее орнитуроморфами, более близкими к современным птицам.
| Имя | Год | Формирование | Место расположения | Примечания | Изображений |
|---|---|---|---|---|---|
| Чиаппевис[30] | 2015 | Формация Цзюфотан (Раннемеловой период, Аптян ) | Обладал веерообразным хвостом, состоящим из множества перьев. | ||
| Eopengornis | 2014 | Формация Хуацзин (Раннемеловой период, Готеривский ) | Самый старый известный член семьи и один из старейших известных энантиорнитов. Обладал очень хорошо сохранившимися хвостовыми лентами. | ||
| Parapengornis[77] | 2015 | Формация Цзюфотан (Раннемеловой период, Аптян ) | Предлагается вести образ жизни дятла из-за особенностей лапы и хвоста. |  | |
| Pengornis | 2008 | Формация Цзюфотан (Раннемеловой период, Аптян ) | Первый обнаруженный пенгорнитид, а также один из крупнейших энантиорнитидов, известных по приличным останкам. |
"Bohaiornithidae"
"Бохайорнитиды «были большими, но геологически недолговечными ранними энантиорнитами, с длинными крючковатыми когтями и крепкими зубами с изогнутыми кончиками. Возможно, они были эквивалентами хищных птиц, хотя эта интерпретация вызывает много споров.[1] В монофилия этой группы сомнительно, и на самом деле это может быть эволюционная ступень.[78]
| Имя | Год | Формирование | Место расположения | Примечания | Изображений |
|---|---|---|---|---|---|
| Бохайорнис | 2011 | Формация Цзюфотан (Раннемеловой период, Аптян ) | Первоначально считалось, что сохранились с гастролиты, хотя позже выяснилось, что это минеральные конкреции | ||
| Гретченияо | 2019 | Формация Исянь (Раннемеловой период, Барремский ) | Приспособлен к взмаху, а не к парящему полету. Может указывать на парафилию или полифилию "Bohaiornithidae". | ||
| Линьорнис[79] | 2016 | Формация Цзюфотан (Раннемеловой период, Аптян ) | Возможный член семьи, известный по хорошо сохранившемуся скелету со структурами, предположительно развивающими яйца. | ||
| Longusunguis | 2014 | Формация Цзюфотан (Раннемеловой период, Аптян ) | Достаточно типичный член семьи |  | |
| Parabohaiornis | 2014 | Формация Цзюфотан (Раннемеловой период, Аптян ) | Близкий родственник Бохайорнис | ||
| Шенкиорнис | 2010 | Цяотоу член Формация Хуацзин (Раннемеловой период, Аптян ?) | Первый известный член семьи, хотя и не считался близким родственником Бохайорнис пока не прошло несколько лет. Сохраняет большую заглазничную кость | ||
| Сулькавис | 2013 | Формация Исянь (Раннемеловой период, Аптян ) | Близкий родственник Шенкиорнис с рифленым эмаль на зубах, уникальный среди ископаемых птиц |  | |
| Жуорнис | 2013 | Формация Цзюфотан (Раннемеловой период, Аптян ) | Крупный член семьи с хорошо сохранившейся черепной коробкой. |
Gobipterygidae
Эта семья может быть монотипный (состоящий только из одного рода или вида), поскольку некоторые члены группы неясны или плохо описаны и могут быть синонимами своего типового вида, Gobipteryx minuta.
| Имя | Год | Формирование | Место расположения | Примечания | Изображений |
|---|---|---|---|---|---|
| Гобиптерикс | 1974 | Формация Барун Гойот (Поздний мел, Кампанский ) | Беззубый продвинутый энантиорнит, обладающий крепким клювом, который эволюционировал вместе с клювом современных птиц. | ||
| Джибейния | 1997 | Цяотоу член Формация Хуацзин (Раннемеловой период, Аптян ?) | Малоизвестен и описан по скелету, который сейчас утерян. Может быть синонимом Vescornis | ||
| Vescornis | 2004 | Цяотоу член Формация Хуацзин (Раннемеловой период, Аптян ?) | Маленький энантиорнит с короткой мордой, который может быть синонимом Джибейния |
Avisauridae
Avisauridae подлежит двум различным определениям различной инклюзивности. Здесь используется более всеобъемлющее определение, которое следует за Cau & Arduini (2008). Авизавриды были давними и широко распространенными энантиорнитами, которые в основном отличались особенностями тарзометатарзалов (костей голеностопных суставов). Самые крупные и продвинутые представители этой группы выжили в Северной и Южной Америке до конца мелового периода, но они очень фрагментарны по сравнению с некоторыми более ранними таксонами.
| Имя | Год | Формирование | Место расположения | Примечания | Изображений |
|---|---|---|---|---|---|
| Авизавр | 1985 | Формация Адского ручья (Поздний мел, Маастрихтский ) | Одноименный авизаврид, а также один из самых крупных представителей семейства. Первоначально считался нептичьим динозавром |  | |
| Энантиофеникс | 2008 | Формация Уади-эль-Габур (Поздний мел, Сеноманский ) | Возможно, питался древесным соком, так как он был сохранен вместе с Янтарь бусы | ||
| Gettyia | 2018 | Формирование двух лекарств (Поздний мел, Кампанский ) | Новый род для Avisaurus gloriae | ||
| Халиморнис | 2002 | Меловая формация Мурвилля (Поздний мел, Кампанский ) | Жил бы в прибрежной среде | ||
| Intiornis | 2010 | Формация Лас Куртиембрес (Поздний мел, Кампанский ) | Хотя представители этого рода были тесно связаны с некоторыми из крупнейших авизаврид, они были очень маленькими птицами. |  | |
| Mirarce | 2018 | Кайпаровицкая свита (Поздний мел, Кампанский ) | Наиболее полный из известных североамериканских авизаврид | ||
| Мистиорнис | 2011 | Илекская формация (Раннемеловой период, Барремский к Аптян ) | Обладает множеством функций из различных групп в Паравес, хотя больше всего похож на авизаврид среди выбранных групп | ||
| Neuquenornis | 1994 | Формация Бахо-де-ла-Карпа (Поздний мел, Сантон ) | Обладает длинными крыльями и перевернутым пальцем, что указывает на хорошие способности к полету и усаживанию. | ||
| Сороавизавр | 1993 | Формация Лечо (Поздний мел, Маастрихтский ) | Очень близкий родственник Авизавр |
Сомнительные роды и известные безымянные экземпляры
- Gobipipus reshetovi: Описан в 2013 г. по образцам эмбрионов в яичной скорлупе из Формация Барун Гойот из Монголия. Эти экземпляры были очень похожи на эмбриональные Гобиптерикс экземпляров, хотя описатели Gobipipus (набор спорных палеонтологов, включая Евгений Курочкин и Санкар Чаттерджи ) считают это отличным.[80]
- Hebeiornis fengningensis: Синоним Vescornis из-за того, что были описаны с того же экземпляра. Несмотря на то, что он был описан в 1999 году, за 5 лет до описания Vescornis, описание было настолько плохим по сравнению с описанием Vescornis что последнее название считается приоритетным для большинства авторов. В результате имя Hebeiornis считается nomen nudum («голое имя»).
- ‘’Proornis ’’ - сомнительная птица из Северной Кореи.
- "Liaoxiornis delicatus ": Описан в 1999 г. по образцу энантиорнитей, найденному в Формация Исянь. Первоначально этот экземпляр считался крошечным взрослым, но позже было обнаружено, что это вылупившийся детеныш. В дальнейшем к этому роду были отнесены другие экземпляры. Из-за отсутствия отличительных признаков многие палеонтологи сочли этот род недиагностическим. номен дубиум.
- LP-4450: Молодь неопределенного энантиорнитея из Эль-Монсек Формирование Испания. Его описание 2006 г. изучило гистология скелета, тогда как более поздние исследования сообщили о наличии чешуйчатой кости в образце, но неизвестной у других энантиорнитей.
- ИВПП В 13939: Кратко описанный в 2004 году, этот иссянский энантиорнитей имел развитые перистые перья на ногах, подобные (хотя и более короткие, чем) перья других паравианцев, таких как Микрораптор и Анхиорнис.[26]
- ДИП-В-15100 и ДИП-В-15101: Два разных крыла вылупившихся особей, которые были описаны в 2015 году. Их описание привлекло значительное внимание средств массовой информации. Они сохранились в исключительных деталях, так как находились в бирманском янтаре примерно 99 миллионов лет.[16]
- HPG-15-1: Частичный труп вылупившегося энантиорнифея, также сохранившийся в бирманском янтаре. Несмотря на неопределенность, он привлек даже больше внимания СМИ, чем два крыла после его описания в 2017 году.[17]
- CUGB P1202: Индерминантный детеныш бохайорнитид от Формация Цзюфотан. Анализ оперения 2016 года показал, что меланосомы, предполагая, что большая часть его оперения была радужный.
- ДИП-В-15102: Еще один труп неопределенного детеныша, сохраненный в бирманском янтаре. Описан в начале 2018 года.[18]
- MPCM-LH-26189 а / б: Частичный скелет детеныша из Лас Хойас в Испании, включая компоненты перекрытий и контрплит. Его описание 2018 года показало, как различные черты характера развивались у энантиорнитов по мере их старения. К таким функциям относятся окостенение из грудина из различных более мелких костей, и слияние хвостовых позвонков в пигостиль.
- YLSNHM01001: Нога и хвост сохранены в бирманском янтаре.[81]
Рекомендации
- ^ а б c Ван, Мин; Чжоу, Чжун-Хэ; О'Коннор, Jingmai K .; Зеленков, Никита В. (2014). «Новое разнообразное семейство энантиорнитинов (Bohaiornithidae fam. Nov.) Из нижнего мела Китая с информацией по двум новым видам» (PDF). Позвоночные PalAsiatica. 52 (1): 31–76.
- ^ а б Ван, X .; О'Коннор, Дж. К .; Чжэн, X .; Wang, M .; Хм.; Чжоу, З. (2014). «Понимание эволюции хвостовых перьев с преобладанием рахиса из нового базального энантиорнитина (Aves: Ornithothoraces)». Биологический журнал Линнеевского общества. 113 (3): 805–819. Дои:10.1111 / bij.12313.
- ^ а б c Chiappe, Луис М .; Уокер, Кирилл А. (2002). "Морфология скелета и систематика меловых Euenantiornithes (Ornithothoraces: Enantiornithes)". In Chiappe, Luis M .; Витмер, Лоуренс М. (ред.). Птицы мезозоя: над головами динозавров. Калифорнийский университет Press. С. 240–67. ISBN 978-0-520-20094-4.
- ^ Чиаппе, Луис М. (2007). Прославленные динозавры: происхождение и ранняя эволюция птиц. Хобокен, Нью-Джерси: Джон Уайли и сыновья. ISBN 978-0-471-24723-4.[страница нужна ]
- ^ О'Коннор, Jingmai K .; Chiappe, Луис М .; Гао, Чуньлин; Чжао, Бо (2011). «Анатомия энантиорнитиновой птицы раннего мела. Rapaxavis pani" (PDF). Acta Palaeontologica Polonica. 56 (3): 463–75. Дои:10.4202 / app.2010.0047.
- ^ Эльзановский, Анджей (1974). «Предварительные сведения о палеогнатных птицах из верхнего мела Монголии» (PDF). Палеонтология Полоника. 29: 103–9.
- ^ а б Уокер, К.А. (1981). «Новый подкласс птиц мелового периода Южной Америки». Природа. 292 (5818): 51–3. Bibcode:1981 Натур 292 ... 51 Вт. Дои:10.1038 / 292051a0.
- ^ Надежда, Сильвия (2002). «Мезозойское излучение неорнитов». In Chiappe, Luis M .; Витмер, Лоуренс М. (ред.). Птицы мезозоя: над головами динозавров. Калифорнийский университет Press. С. 339–88. ISBN 978-0-520-20094-4.
- ^ Федучча, Алан (1996). Происхождение и эволюция птиц. Нью-Хейвен: издательство Йельского университета. ISBN 978-0-300-06460-5.[страница нужна ]
- ^ Ты, Хай-лу; Ламанна, Мэтью С .; Харрис, Джеральд Д.; Chiappe, Луис М .; О'Коннор, Джингмай; Цзи, Шуан; Люй, Цзюнь-чан; Юань, Чун-си; Ли, Да-цин; Чжан, Син; Lacovara, Kenneth J .; Додсон, Питер; Цзи, Цян (16 июня 2006 г.). «Почти современная птица-амфибия из раннего мела Северо-Западного Китая». Наука. 312 (5780): 1640–1643. Bibcode:2006Научный ... 312.1640Y. Дои:10.1126 / science.1126377. PMID 16778053.
- ^ а б Чжан, Цзихуэй; Chiappe, Луис М .; Хан, банда; Чинсами, Анусуя (2013). «Крупная птица раннего мела Китая: новые сведения о черепе энантиорнитинов». Журнал палеонтологии позвоночных. 33 (5): 1176–89. Дои:10.1080/02724634.2013.762708.
- ^ а б Чжоу, Чжунхэ; Кларк, Джулия; Чжан, Фучэн (май 2008 г.). «Понимание разнообразия, размеров тела и морфологической эволюции крупнейшей энантиорнитиновой птицы раннего мелового периода». Журнал анатомии. 212 (5): 565–77. Дои:10.1111 / j.1469-7580.2008.00880.x. ЧВК 2409080. PMID 18397240.
- ^ Ху, Дунъюй; Сюй, Син; Хоу, Ляньхай; Салливан, Корвин (2012). "Новая энантиорнитиновая птица из нижнего мела Западного Ляонина, Китай, и ее значение для ранней эволюции птиц". Журнал палеонтологии позвоночных. 32 (3): 639–45. Дои:10.1080/02724634.2012.652321.
- ^ Мартин, Энтони Дж .; Викерс-Рич, Патрисия; Rich, Thomas H .; Холл, Майкл; Angielczyk, Кеннет (январь 2014 г.). «Самые старые известные следы птиц из Австралии: формация Эумералла (Альбиан), бухта Динозавров, Виктория». Палеонтология. 57 (1): 7–19. Дои:10.1111 / pala.12082.
- ^ Аттерхолт, Джесси; Хатчисон, Дж. Ховард; О’Коннор, Джингмай К. (13 ноября 2018 г.). «Самый полный энантиорнитин из Северной Америки и филогенетический анализ Avisauridae». PeerJ. 6: e5910. Дои:10.7717 / peerj.5910. ЧВК 6238772. PMID 30479894.
- ^ а б c Син, Лида; Маккеллар, Райан С .; Ван, Мин; Бай, Мин; О’Коннор, Джингмай К .; Бентон, Майкл Дж .; Чжан, Цзяньпин; Ван, Ян; Ценг, Куовей; Локли, Мартин Дж .; Ли, банда; Чжан, Вэйвэй; Сюй, Син (28 июня 2016 г.). «Мумифицированные крылья скороспелых птиц в бирманском янтаре среднего мела». Nature Communications. 7 (1): 12089. Bibcode:2016НатКо ... 712089X. Дои:10.1038 / ncomms12089. ЧВК 4931330. PMID 27352215.
- ^ а б Син, Лида; О'Коннор, Jingmai K .; Маккеллар, Райан С .; Chiappe, Луис М .; Ценг, Куовей; Ли, банда; Бай, Мин (сентябрь 2017 г.). «Птенец энантиорнитина (Aves) среднего мела, сохранившийся в бирманском янтаре с необычным оперением». Исследования Гондваны. 49: 264–277. Bibcode:2017GondR..49..264X. Дои:10.1016 / j.gr.2017.06.001.
- ^ а б Син, Лида; О'Коннор, Jingmai K .; Маккеллар, Райан С .; Chiappe, Луис М .; Бай, Мин; Ценг, Куовей; Чжан, Цзе; Ян, Хайдун; Фанг, Джун; Ли, Банда (февраль 2018 г.). «Уплощенный энантиорнитин в бирманском янтаре среднего мела: морфология и сохранение». Научный бюллетень. 63 (4): 235–243. Дои:10.1016 / j.scib.2018.01.019.
- ^ Син, Лида; Маккеллар, Райан С .; О’Коннор, Джингмай К .; Бай, Мин; Ценг, Куовей; Чиаппе, Луис М. (30 января 2019 г.). «Полностью оперенный энантиорнитиновый фрагмент ступни и крыла, сохранившийся в бирманском янтаре среднего мелового периода». Научные отчеты. 9 (1): 927. Bibcode:2019НатСР ... 9..927X. Дои:10.1038 / с41598-018-37427-4. ЧВК 6353931. PMID 30700773.
- ^ Син, Лида; Маккеллар, Райан С .; О'Коннор, Джингмай К. (июнь 2020 г.). «Необычно большое крыло птицы в бирманском янтаре среднего мелового периода». Меловые исследования. 110: 104412. Дои:10.1016 / j.cretres.2020.104412.
- ^ а б О’Коннор, Джингмай К .; Чиаппе, Луис М. (28 февраля 2011 г.). «Пересмотр морфологии черепа энантиорнитином (Aves: Ornithothoraces)». Журнал систематической палеонтологии. 9 (1): 135–157. Дои:10.1080/14772019.2010.526639.
- ^ а б c Ван, Мин; Ху, Хань; Ли, Чжихэн (21 августа 2015 г.). «Новая маленькая энантиорнитиновая птица из биоты Джехол, имеющая значение для ранней эволюции морфологии птичьего черепа». Журнал систематической палеонтологии. 14 (6): 481–497. Дои:10.1080/14772019.2015.1073801.
- ^ Ван, Мин; Ху, Хан (январь 2017 г.). «Сравнительное морфологическое исследование скуловой кости и квадратично-скуловой кости у ранних птиц и их динозавров». Анатомический рекорд. 300 (1): 62–75. Дои:10.1002 / ar.23446. PMID 28000410.
- ^ Чиаппе, Луис М. (2009). «Уменьшенные динозавры: эволюционный переход к современным птицам». Эволюция: образование и пропаганда. 2 (2): 248–56. Дои:10.1007 / s12052-009-0133-4.
- ^ Беккер, Рэйчел (28 июня 2016 г.). «Крылья птиц, застрявшие в янтаре, - первые ископаемые из эпохи динозавров». Природа. Дои:10.1038 / природа.2016.20162.
- ^ а б Чжан, Фучэн; Чжоу, Чжунхэ (октябрь 2004 г.). «Палеонтология: перья на ногах у птиц раннего мелового периода». Природа. 431 (7011): 925. Bibcode:2004 Натур.431..925Z. Дои:10.1038 / 431925a. PMID 15496911.
- ^ а б Кларк, Джулия А .; Чжоу, Чжунхэ; Чжан, Фучэн (март 2006 г.). "Понимание эволюции полета птиц из новой клады раннемеловых орнитурин из Китая и морфологии Иксианорнис грабауи". Журнал анатомии. 208 (3): 287–308. Дои:10.1111 / j.1469-7580.2006.00534.x. ЧВК 2100246. PMID 16533313.
- ^ а б О'Коннор, Jingmai K .; Ван, Сюри; Chiappe, Луис М .; Гао, Чуньлин; Мэн, Цинцзинь; Чэн, Сяодун; Лю, Цзиньюань (12 марта 2009 г.). «Филогенетическая поддержка специализированной клады энантиорнитиновых птиц мелового периода с информацией о новом виде». Журнал палеонтологии позвоночных. 29 (1): 188–204. Дои:10.1080/02724634.2009.10010371.
- ^ а б Chiappe, Луис М .; Бо, Чжао; О'Коннор, Jingmai K .; Чуньлин, Гао; Сюри, Ван; Хабиб, Майкл; Маруган-Лобон, Иисус; Цинцзинь, Мэн; Сяодун, Чэн (2014). «Новый образец раннемеловой птицы. Hongshanornis longicresta: информация об аэродинамике и диете базального орнитуроморфа ". PeerJ. 2: e234. Дои:10.7717 / peerj.234. ЧВК 3898307. PMID 24482756.
- ^ а б c d О’Коннор, Джингмай К .; Ван, Сяоли; Чжэн, Сяотин; Ху, Хань; Чжан, Сяомэй; Чжоу, Чжунхэ (январь 2016 г.). «Энантиорнитин с веерообразным хвостом и эволюция Rectricial комплекса у ранних птиц». Текущая биология. 26 (1): 114–119. Дои:10.1016 / j.cub.2015.11.036. PMID 26748849.
- ^ а б О’Коннор, Джингмай К .; Ли, Да-Цин; Ламанна, Мэтью С .; Ван, Мин; Харрис, Джеральд Д.; Аттерхолт, Джесси; Ю, Хай-Лу (30 декабря 2015 г.). «Новый раннемеловой энантиорнитин (Aves, Ornithothoraces) из северо-западного Китая с изысканным орнаментом на хвосте». Журнал палеонтологии позвоночных. 36 (1): e1054035. Дои:10.1080/02724634.2015.1054035.
- ^ а б Ван, Мин; О’Коннор, Джингмай К .; Пан, Яньхун; Чжоу, Чжунхэ (31.01.2017). «Причудливая энантиорнитиновая птица раннего мела с уникальными перьями голени и орнитуроморфным пигостилем в форме плуга». Nature Communications. 8: 14141. Bibcode:2017НатКо ... 814141W. Дои:10.1038 / ncomms14141. ЧВК 5290326. PMID 28139644.
- ^ а б Sanz, José L .; Chiappe, Луис М .; Бускалиони, Анджела Д. (1995). "Остеология Concornis lacustris (Aves: Enantiornithes) из нижнего мела Испании и пересмотр его филогенетических связей ». Американский музей Novitates (3133): 1–23. HDL:2246/3667.
- ^ Dalla Vecchia, Fabio M .; Чиаппе, Луис М. (2003). «Первый скелет птицы из мезозоя северной Гондваны». Журнал палеонтологии позвоночных. 22 (4): 856–60. Дои:10.1671 / 0272-4634 (2002) 022 [0856: FASFTM] 2.0.CO; 2. JSTOR 4524284.
- ^ О'Коннор, Джингмай К. (1 января 2019 г.). «Трофические повадки ранних пташек». Палеогеография, палеоклиматология, палеоэкология. 513: 178–195. Bibcode:2019ППП ... 513..178О. Дои:10.1016 / j.palaeo.2018.03.006.
- ^ О'Коннор, Jingmai K .; Чжоу, Чжунхэ; Смит, Эндрю (6 ноября 2019 г.). «Эволюция современной пищеварительной системы птиц: выводы из окаменелостей паравиевых отложений биот Янляо и Джехол». Палеонтология. 63 (1): 13–27. Дои:10.1111 / pala.12453.
- ^ Хьюстон, Дэвид К .; Копси, Дж. А. (1994). «Костное пищеварение и морфология кишечника бородатого стервятника». Журнал исследований хищников. 28 (2): 73–78.
- ^ Sanz, José L .; Chiappe, Луис М .; Фернадес-Ялво, Иоланда; Ортега, Франсиско; Санчес-Чильон, Бегонья; Poyato-Ariza1, Francisco J .; Перес-Морено, Бернардино П. (февраль 2001 г.). «Раннемеловой окатыш». Природа. 409 (6823): 998–1000. Bibcode:2001Натура.409..998С. Дои:10.1038/35059172. PMID 11234054.
- ^ Михайлов, Константин Э. (1991). «Классификация ископаемых яичных скорлуп амниотических позвоночных» (PDF). Acta Palaeontologica Polonica. 36 (2): 193–238.
- ^ Михайлов, Константин Э. (1996). «Новые роды ископаемых яиц из верхнего мела Монголии». Палеонтологический журнал. 30 (2): 246–8.
- ^ Елжановский, Анджей (1981). «Эмбриональные скелеты птиц из позднего мела Монголии» (PDF). Палеонтология Полоника. 42: 147–79.
- ^ Sanz, José L .; Chiappe, Луис М .; Перес-Морено, Бернардино П .; Moratalla, José J .; Эрнандес-Карраскилья, Франсиско; Buscalioni, Angela D .; Ортега, Франсиско; Poyato-Ariza, Francisco J .; Расскин-Гутман, Диего; Мартинес-Делкло, Ксавьер (6 июня 1997 г.). «Птенец птицы из нижнего мела Испании: последствия для эволюции черепа и шеи птиц». Наука. 276 (5318): 1543–6. Дои:10.1126 / science.276.5318.1543.
- ^ Чжоу, Чжунхэ; Чжан, Фучэн (22 октября 2004 г.). «Ранний птичий эмбрион из нижнего мела Китая». Наука. 306 (5696): 653. Дои:10.1126 / science.1100000. PMID 15499011.
- ^ а б c Chiappe, Луис М .; Шуань, Цзи; Цян, Цзи (2007). «Молодые птицы из раннего мела Китая: последствия для энантиорнитинового онтогенеза». Американский музей Novitates. 3594: 1–46. Дои:10.1206 / 0003-0082 (2007) 3594 [1: JBFTEC] 2.0.CO; 2. HDL:2246/5890.
- ^ Елжановский, Анджей (1995). «Меловые птицы и птичья филогения». Курьер Forschungsinstitut Senckenberg. 181: 37–53.
- ^ Курочкин, Э.Н .; Chatterjee, S .; Михайлов, К. (2013). «Эмбриональная энантиорнитиновая птица и связанные яйца из мелового периода Монголии». Палеонтологический журнал. 47 (11): 1252–69. Дои:10.1134 / S0031030113110087.
- ^ Курочкин, Э. Н .; Chatterjee, S .; Михайлов, К. Э. (19 декабря 2013 г.). «Эмбриональная энантиорнитиновая птица и связанные яйца из мелового периода Монголии». Палеонтологический журнал. 47 (11): 1252–1269. Дои:10.1134 / S0031030113110087.
- ^ Камбра-Му, Оскар; Бускалиони, Анхела Дельгадо; Кубо, Хорхе; Кастане, Жак; Лот, Мари-Мадлен; де Маржери, Эммануэль; де Риклес, Арман (2006). «Гистологические наблюдения энантиорнитиновой кости (Saurischia, Aves) из нижнего мела Лас-Ойяс (Испания)». Comptes Rendus Palevol. 5 (5): 685–91. Дои:10.1016 / j.crpv.2005.12.018.
- ^ О'Коннор, Jingmai K .; Ван, Мин; Чжэн, Сяо-Тин; Ван, Сяо-Ли; Чжоу, Чжун-Хе (2014). «Гистология двух самок ранних меловых птиц» (PDF). Позвоночные PalAsiatica. 52 (1): 112–28.
- ^ Чиаппе, Л.М. (1995). "Филогенетическое положение меловых птиц Аргентины: Enantiornithes и Патагоптерикс deferrariisi". Курьер Forschungsinstitut Senckenberg. 181: 55–63.
- ^ Дайк, Гарет; Времир, Матьяс; Кайзер, Гэри; Нейш, Даррен (июнь 2012 г.). «Затонувшая мезозойская гнездовая колония птиц из позднего мела Трансильвании». Die Naturwissenschaften. 99 (6): 435–42. Bibcode:2012NW ..... 99..435D. CiteSeerX 10.1.1.394.9006. Дои:10.1007 / s00114-012-0917-1. PMID 22575918.
- ^ Fernández, Mariela S .; Гарсия, Родольфо А .; Фиорелли, Лукас; Сколаро, Алехандро; Сальвадор, Родриго Б.; Котаро, Карлос Н .; Кайзер, Гэри У .; Дайк, Гарет Дж .; Фарке, Эндрю А. (17 апреля 2013 г.). «Большое скопление птичьих яиц из позднего мела в Патагонии (Аргентина) раскрывает новую стратегию гнездования мезозойских птиц». PLOS ONE. 8 (4): e61030. Bibcode:2013PLoSO ... 861030F. Дои:10.1371 / journal.pone.0061030. ЧВК 3629076. PMID 23613776.
- ^ О'Коннор, Jingmai K .; Фальк, Аманда; Ван, Мин; Чжэн, Сяо-Тин (2020). «Первое сообщение о незрелых перьях у молодых энантиорнитинов из раннемеловой орнитофауны Jehol». Позвоночные PalAsiatica. 58: 24–44. Дои:10.19615 / j.cnki.1000-3118.190823.
- ^ Падиан, Кевин; Чиаппе, Луис М. (11 января 2007 г.). «Происхождение и ранняя эволюция птиц». Биологические обзоры. 73 (1): 1–42. Дои:10.1111 / j.1469-185x.1997.tb00024.x.
- ^ Чжоу, Шуанг; Чжоу, Чжун-Хэ; О'Коннор, Джингмай К. (2012). «Новая орнитовая птица с базальным клювом из нижнего мела Западного Ляонина, Китай» (PDF). Позвоночные PalAsiatica. 50 (1): 9–24. Сложить резюме – Phys.org (13 февраля 2012 г.).
- ^ Худ, Питер В. (1988). «Палеогнатные птицы из раннего третичного периода Северного полушария». Публикации Орнитологического клуба Nuttall (Кембридж, Массачусетс, США: Nuttall Ornithological Club) 22
- ^ Навалон, Гильермо; Маруган-Лобон, Хесус; Chiappe, Луис М .; Луис Санс, Хосе; Бускалиони, Анхела Д. (6 октября 2015 г.). «Расположение мягких тканей и дермы в крыле раннемеловой птицы: последствия для эволюции полета птиц». Научные отчеты. 5 (1): 14864. Bibcode:2015НатСР ... 514864Н. Дои:10.1038 / srep14864. ЧВК 4594305. PMID 26440221.
- ^ Ван, Ся; Макгоуэн, Алистер Дж .; Дайк, Гарет Дж .; Терви, Сэмюэл Т. (7 декабря 2011 г.). "Пропорции крыльев и стили полета птиц: первый шаг к прогнозированию режимов полета мезозойских птиц". PLOS ONE. 6 (12): e28672. Bibcode:2011PLoSO ... 628672W. Дои:10.1371 / journal.pone.0028672. ЧВК 3233598. PMID 22163324.
- ^ Chiappe, Луис М .; Сузуки, Сигэру; Дайк, Гарет Дж .; Ватабе, Махито; Цогтбаатар, К .; Барсболд, Ринчен (январь 2007 г.). «Новая энантиорнитиновая птица из позднего мела пустыни Гоби». Журнал систематической палеонтологии. 5 (2): 193–208. Дои:10.1017 / S1477201906001969.
- ^ Чжэн, Сяотин; Ван, Сяоли; О'Коннор, Джингмай; Чжоу, Чжунхэ (9 октября 2012 г.). «Понимание ранней эволюции грудины птиц из молодых энантиорнитинов». Nature Communications. 3 (1): 1116. Bibcode:2012 НатКо ... 3.1116Z. Дои:10.1038 / ncomms2104. PMID 23047674.
- ^ а б О'Коннор, Джингмай; Дайк, Гарет (2010). "Переоценка Sinornis santensis и Cathayornis yandica (Aves: Enantiornithes) ". Записи Австралийского музея. 62: 7–20. Дои:10.3853 / j.0067-1975.62.2010.1540.
- ^ Кларк, Джулия А .; Норелл, Марк А. (2002). «Морфология и филогенетическое положение Апсаравис ухаана из позднего мела Монголии ». Американский музей Novitates. 3387: 1–46. CiteSeerX 10.1.1.693.8475. Дои:10.1206 / 0003-0082 (2002) 387 <0001: TMAPPO> 2.0.CO; 2.
- ^ а б Серено, П. (2005) Таксон Поиск: Стеблевая архозаврия В архиве 2007-02-19 в Wayback Machine. Версия 1.0, 2005-НОЯ-7. Источник 2006-ОКТ-02.
- ^ Карвалью; Новас; Аньолин; Исаси; Фрейтас; Андраде (2015). «Новый род и вид энантиорнитиновой птицы из раннего мела Бразилии». Бразильский журнал геологии. 45 (2): 161–171. Дои:10.1590/23174889201500020001.
- ^ Ван, Ли; О'Коннор, Чжоу; Ты (2015). «Второй вид энантиорнитиновых птиц из нижнего мела бассейна Чангма, северо-западный Китай с последствиями для таксономического разнообразия орнитофауны Чангмы». Меловые исследования. 55: 56–65. Дои:10.1016 / j.cretres.2015.01.008.
- ^ Wang, M .; О'Коннор, Дж. К .; Чжоу, З. (2014). «Новая здоровая птица-энантиорнитин из нижнего мела Китая с приспособлениями для сканирования». Журнал палеонтологии позвоночных. 34 (3): 657–671. Дои:10.1080/02724634.2013.812101.
- ^ Dalsätt, J .; Ericson, P.G .; Чжоу, З. (2015). «Новый энантиорнитес (авес) из раннего мела Китая». Acta Geologica Sinica. 86 (2): 801–807. Дои:10.1111/1755-6724.12270.
- ^ Белл, Алисса К .; Chiappe, Луис М .; Эриксон, Грегори М .; Сузуки, Сигэру; Ватабе, Махито; Барсболд, Ринчен; Цогтбаатар, К. (февраль 2010 г.). «Описание и экологический анализ Hollanda luceria, позднемеловой птицы из пустыни Гоби (Монголия)». Меловые исследования. 31 (1): 16–26. Дои:10.1016 / j.cretres.2009.09.001.
- ^ Хартман, Скотт; Мортимер, Микки; Wahl, William R .; Lomax, Dean R .; Липпинкотт, Джессика; Лавлейс, Дэвид М. (10 июля 2019 г.). «Новый паравианский динозавр из поздней юры Северной Америки поддерживает позднее приобретение птичьего полета». PeerJ. 7: e7247. Дои:10.7717 / peerj.7247. ЧВК 6626525. PMID 31333906.
- ^ Зеленков, Никита В .; Аверьянов, Александр Олегович (13 июня 2015 г.). «Исторический образец энантиорнитиновой птицы из раннего мела Монголии, представляющий новый таксон со специальной морфологией шеи». Журнал систематической палеонтологии. 14 (4): 319–338. Дои:10.1080/14772019.2015.1051146.
- ^ Wang, M .; Лю Д. (2015). «Таксономическая переоценка Cathayornithidae (Aves: Enantiornithes)». Журнал систематической палеонтологии. 14: 1–19. Дои:10.1080/14772019.2014.994087.
- ^ Лю, Ди; Chiappe, Луис М .; Серрано, Франсиско; Хабиб, Майкл; Чжан, Югуан; Мэн, Циньцзин; Шоуки, Мэтью (11 октября 2017 г.). «Аэродинамика полета в энантиорнитинах: информация от новой китайской раннемеловой птицы». PLOS One. 12 (10): e0184637. Bibcode:2017PLoSO..1284637L. Дои:10.1371 / journal.pone.0184637. ЧВК 5636078. PMID 29020077.
- ^ Ху, Хань; О'Коннор, Джингмай К. (14 ноября 2016 г.). «Первый вид энантиорнитов из Сихеданга объясняет развитие скелета в раннемеловых энантиорнитинах». Журнал систематической палеонтологии. 15 (11): 909–926. Дои:10.1080/14772019.2016.1246111.
- ^ Ван, Мин; Чжоу, Чжунхэ; Сюй, Гуанхуй (7 января 2014 г.). «Первая энантиорнитиновая птица из верхнего мела Китая». Журнал палеонтологии позвоночных. 34 (1): 135–145. Дои:10.1080/02724634.2013.794814.
- ^ Ван, Мин; Чжоу, Чжунхэ (12 апреля 2017 г.). «Морфологическое исследование первой известной рыбоядной энантиорнитиновой птицы из раннего мела Китая». Журнал палеонтологии позвоночных. 37 (2): e1278702. Дои:10.1080/02724634.2017.1278702.
- ^ Ху, Дунъюй; Лю, Инь; Ли, Цзиньхуа; Сюй, Син; Хоу, Ляньхай (июль 2015 г.). «Yuanjiawaornis viriosus, gen. Et sp. Nov., Большая энантиорнитиновая птица из нижнего мела западного Ляонина, Китай». Меловые исследования. 55: 210–219. Дои:10.1016 / j.cretres.2015.02.013.
- ^ Ху, Хань; О’Коннор, Джингмай К .; Чжоу, Чжунхэ; Фарке, Эндрю А. (3 июня 2015 г.). «Новый вид Pengornithidae (Aves: Enantiornithes) из нижнего мела Китая предлагает специализированную среду обитания, ранее неизвестную у ранних птиц». PLOS One. 10 (6): e0126791. Bibcode:2015PLoSO..1026791H. Дои:10.1371 / journal.pone.0126791. ЧВК 4454694. PMID 26039693.
- ^ Chiappe, Луис М .; Цинцзинь, Мэн; Серрано, Франсиско; Сигурдсен, Тронд; Мин, Ван; Белл, Алисса; Ди, Лю (25 октября 2019 г.). "Новый Бохайорнис-подобная птица из раннего мела Китая: энантиорнитиновые взаимоотношения и летные качества ». PeerJ. 7: e7846. Дои:10.7717 / peerj.7846. ЧВК 6816414. PMID 31667014.
- ^ Ван, Ян; Ван, Мин; О'Коннор, Jingmai K .; Ван, Сяоли; Чжэн, Сяотин; Чжан Сяомэй (11 января 2016 г.). «Новая птица Jehol enantiornithine с трехмерным сохранением и фолликулами яичников». Журнал палеонтологии позвоночных. 36 (2): e1054496. Дои:10.1080/02724634.2015.1054496.
- ^ Курочкин, Э. Н .; Chatterjee, S .; Михайлов, К. Э. (19 декабря 2013 г.). «Эмбриональная энантиорнитиновая птица и связанные яйца из мелового периода Монголии». Палеонтологический журнал. 47 (11): 1252–1269. Дои:10.1134 / s0031030113110087.
- ^ Син, Лида; Маккеллар, Райан С .; О’Коннор, Джингмай К .; Ниу, Кеченг; Май, Хуэйцзюань (29 октября 2019 г.). "Энантиорнитиновая ступня и перо хвоста среднего мелового периода, сохраненные в бирманском янтаре". Научные отчеты. 9 (1): 15513. Bibcode:2019НатСР ... 915513X. Дои:10.1038 / s41598-019-51929-9. ЧВК 6820775. PMID 31664115.
внешняя ссылка
![]() СМИ, связанные с Энантиорниты в Wikimedia Commons
СМИ, связанные с Энантиорниты в Wikimedia Commons

















Птицы (учебный класс: Авес) | |||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Анатомия | |||||||||||||||||||||||||||||||||||||||||||||||||||||
| Поведение | |||||||||||||||||||||||||||||||||||||||||||||||||||||
| Эволюция | |||||||||||||||||||||||||||||||||||||||||||||||||||||
| Ископаемые птицы | |||||||||||||||||||||||||||||||||||||||||||||||||||||
| Человек взаимодействие | |||||||||||||||||||||||||||||||||||||||||||||||||||||
| Списки | |||||||||||||||||||||||||||||||||||||||||||||||||||||
| Neornithes |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||