Хроматофор - Chromatophore - Wikipedia

Хроматофоры находятся пигмент -содержащие клетки или группы клеток, обнаруженные у широкого круга животных, включая амфибии, рыбы, рептилии, ракообразные и головоногие моллюски. Млекопитающие и птицы, напротив, имеют класс ячеек, называемый меланоциты за окраска.
Хроматофоры в значительной степени ответственны за образование кожи и цвет глаз в экзотермический животных и генерируются в нервный гребень в течение эмбриональное развитие. Зрелые хроматофоры сгруппированы в подклассы в зависимости от их цвета (точнее, "оттенок ") при белом свете: ксантофоры (желтый), эритрофоры (красный), иридофоры (отражающий / радужный ), лейкофоры (белый), меланофоры (черный / коричневый) и цианофоры (синий).

Некоторые виды могут быстро менять цвет с помощью механизмов, которые перемещают пигмент и переориентируют отражающие пластинки внутри хроматофоров. Этот процесс, часто используемый в качестве камуфляж, называется физиологическое изменение цвета или же метахроз.[1] Головоногие, такие как осьминог имеют сложные хроматофорные органы, контролируемые мышцами, в то время как позвоночные, такие как хамелеоны произвести аналогичный эффект клеточная сигнализация. Такие сигналы могут быть гормоны или же нейротрансмиттеры и может быть вызвано изменениями настроения, температуры, стресса или видимыми изменениями в окружающей среде.[нужна цитата ] Хроматофоры изучаются учеными для понимания болезней человека и как инструмент в открытие лекарств.
Человеческое открытие
Аристотель упомянул способность осьминог изменить цвет для обоих камуфляж и сигнализация в его Historia animalium (около 400 г. до н.э.):[2]
Осьминог ... ищет свою жертву, меняя цвет так, чтобы он походил на цвет соседних с ним камней; он делает то же самое при тревоге.
Giosuè Sangiovanni был первым, кто описал беспозвоночный пигментные клетки как хромофоро в итальянском научном журнале в 1819 году.[3]
Чарльз Дарвин описал способность менять цвет каракатица в Путешествие Бигля (1860):[4]
Эти животные также избегают обнаружения благодаря очень необычной, подобной хамелеону способности изменять свой цвет. Похоже, что они меняют свой оттенок в зависимости от характера земли, по которой они проходят: на глубокой воде их общий оттенок был коричневато-пурпурным, но при размещении на суше или на мелководье этот темный оттенок менялся на один из желтовато-зеленый. Цвет, при более внимательном рассмотрении, был французско-серым с многочисленными мельчайшими ярко-желтыми пятнами: первые из них различались по интенсивности; последние полностью исчезли и по очереди появлялись снова. Эти изменения были произведены таким образом, что облака, варьирующиеся по оттенку от красного гиацинта до каштаново-коричневого, непрерывно проходили по телу. Любая часть, подвергаясь легкому гальваническому удару, становилась почти черной: аналогичный эффект, но в меньшей степени, производился при царапании кожи иглой. Считается, что эти облака, или, как их еще называют, румянец, производятся в результате попеременного расширения и сжатия мельчайших пузырьков, содержащих жидкости разного цвета.
Классификация

Период, термин хроматофор был принят (после Сангиованни хромофор) как название пигментных клеток, происходящих от нервного гребня хладнокровных позвоночные и головоногие моллюски. Само слово происходит от Греческий слова Chrōma (χρῶμα), что означает «цвет», и Форос (φόρος), что означает "подшипник". Напротив, слово хроматоцит (Китос (κύτος) означает «клетка») был принят для клеток, отвечающих за цвет, обнаруженных у птиц и млекопитающих. Только один такой тип ячеек, меланоцит, был идентифицирован у этих животных.
Хроматофоры стали достаточно хорошо изучены только в 1960-х годах, чтобы их можно было классифицировать по внешнему виду. Эта система классификации существует и по сей день, хотя биохимия пигментов может быть более полезным для научного понимания того, как функционируют клетки.[5]
Молекулы, производящие цвет, делятся на два разных класса: биохромы и структурные цвета или «схемохромы».[6] Биохромы включают настоящие пигменты, такие как каротиноиды и птеридины. Эти пигменты избирательно поглощают части видимый световой спектр что составляет белый свет, позволяя другим длины волн дотянуться до глаза наблюдателя. Структурные цвета получаются за счет различных комбинаций дифракции, отражения или рассеяния света от структур с масштабом около четверти длины волны света. Многие такие структуры мешают некоторым длинам волн (цветов) света и пропускают другие просто из-за своего масштаба, поэтому они часто производят переливчатость, создавая разные цвета при взгляде с разных сторон.[нужна цитата ]
Поскольку все хроматофоры содержат пигменты или отражающие структуры (за исключением случаев, когда мутация, как в альбинизм ), не все клетки, содержащие пигмент, являются хроматофорами. Haem, например, это биохром, ответственный за красный цвет крови. Встречается в основном в красные кровяные тельца (эритроциты), которые образуются в костном мозге на протяжении всей жизни организма, а не образуются во время эмбриологического развития. Поэтому эритроциты не относятся к хроматофорам.[нужна цитата ]
Ксантофоры и эритрофоры
Хроматофоры, содержащие большое количество желтый птеридиновые пигменты называются ксантофорами; те, у кого в основном красный /апельсин каротиноиды называются эритрофорами.[5] Тем не мение, пузырьки содержащие птеридин и каротиноиды иногда обнаруживаются в одной и той же клетке, и в этом случае общий цвет зависит от соотношения красного и желтого пигментов.[7] Следовательно, различие между этими типами хроматофоров не всегда ясно.
Большинство хроматофоров могут генерировать птеридины из гуанозинтрифосфат, но ксантофоры, по-видимому, имеют дополнительные биохимические пути, позволяющие им накапливать желтый пигмент. В отличие от каротиноидов метаболизированный и переносится к эритрофорам. Впервые это было продемонстрировано путем выращивания обычно зеленых лягушек на диете из каротин -ограниченный сверчки. Отсутствие каротина в рационе лягушек означало, что красный / оранжевый каротиноидный цветной «фильтр» не присутствовал в их эритрофорах. Из-за этого лягушки казались синими, а не зелеными.[8]
Иридофоры и лейкофоры
Иридофоры, иногда также называемые гуанофорами, представляют собой пигментные клетки, которые отражают свет с помощью пластинок кристаллических хемохромов, сделанных из гуанин.[9] При освещении они создают переливающиеся цвета из-за дифракции света внутри уложенных друг на друга пластин. Ориентация схемы определяет характер наблюдаемого цвета.[10] Используя биохромы в качестве цветных фильтров, иридофоры создают оптический эффект, известный как Tyndall или же Рэлеевское рассеяние, производя яркие-синий или же -зеленый цвета.[11]
Родственный тип хроматофоров, лейкофор, встречается у некоторых рыб, в частности у Tapetum lucidum. Как иридофоры, они используют кристаллический пурины (часто гуанин) для отражения света. Однако, в отличие от иридофоров, лейкофоры имеют более организованные кристаллы, которые уменьшают дифракцию. При наличии источника белого света они производят белый блеск. Как и в случае с ксантофорами и эритрофорами, у рыб различие между иридофорами и лейкофорами не всегда очевидно, но в целом считается, что иридофоры вызывают радужные или цвета металлик, тогда как лейкофоры дают светоотражающие белые оттенки.[11]
Меланофоры
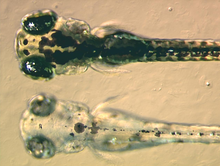
Меланофоры содержат эумеланин, тип меланин, что появляется чернить или темно-коричневый из-за его светопоглощающих свойств. Он упакован в пузырьки, называемые меланосомами, и распределяется по клетке. Эумеланин образуется из тирозин в серии катализированных химических реакций. Это сложное химическое вещество, содержащее единицы дигидроксииндол и дигидроксииндол-2-карбоновая кислота с некоторыми пиррол кольца.[12] Ключевым ферментом в синтезе меланина является тирозиназа. Когда этот белок является дефектным, не может образовываться меланин, что приводит к определенным типам альбинизма. У некоторых видов амфибий наряду с эумеланином содержатся и другие пигменты. Например, новый пигмент глубокого (винного) красного цвета был обнаружен в меланофорах филломедузиновые лягушки.[13] Впоследствии это было идентифицировано как птерорходин, птеридин димер который накапливается вокруг ядра эумеланина, а также присутствует в различных древесная лягушка виды из Австралия и Папуа - Новая Гвинея. Хотя вполне вероятно, что другие менее изученные виды имеют сложные пигменты меланофоров, тем не менее верно, что большинство изученных на сегодняшний день меланофоров действительно содержат исключительно эумеланин.[14]
У людей есть только один класс пигментных клеток, эквивалент меланофоров у млекопитающих, для создания цвета кожи, волос и глаз. По этой причине, а также из-за того, что большое количество и контрастный цвет клеток обычно упрощает их визуализацию, меланофоры, безусловно, являются наиболее изученными хроматофорами. Однако есть различия между биологией меланофоров и биологией меланоциты. В дополнение к эумеланину меланоциты могут генерировать желтый / красный пигмент, называемый феомеланин.[нужна цитата ]

Цианофоры
Почти весь яркий блюз у животных и растений создан структурная окраска а не пигментами. Однако некоторые виды Synchiropus splendidus обладают везикулами голубой биохром неизвестной химической структуры в клетках, названный цианофорами.[11] Хотя они кажутся необычными в своем ограниченном таксономическом диапазоне, могут быть цианофоры (а также другие необычные типы хроматофоров) у других рыб и земноводных. Например, ярко окрашенные хроматофоры с неопределенными пигментами встречаются в обоих Ядовитые лягушки-дротики и стеклянные лягушки,[15] и нетипичный двухцветный хроматофоры, названные эритро-иридофоры были описаны в Псевдохромная диадема.[16]
Транслокация пигмента

Многие виды способны перемещать пигмент внутри своих хроматофоров, что приводит к заметному изменению цвета тела. Этот процесс, известный как физиологический изменение цвета, наиболее широко изучен у меланофоров, поскольку меланин - самый темный и наиболее заметный пигмент. У большинства видов с относительно тонким дерма дермальные меланофоры имеют тенденцию быть плоскими и покрывать большую площадь поверхности. Однако у животных с толстым дермальным слоем, таких как взрослые рептилии, дермальные меланофоры часто образуют трехмерные единицы с другими хроматофорами. Эти дермальные хроматофорные единицы (DCU) состоят из самого верхнего слоя ксантофора или эритрофора, затем слоя иридофора и, наконец, корзинообразного слоя меланофоров с отростками, покрывающими иридофоры.[17]
Оба типа меланофоров важны для физиологического изменения цвета. Плоские кожные меланофоры часто перекрывают другие хроматофоры, поэтому, когда пигмент рассредоточен по клетке, кожа выглядит темной. Когда пигмент собирается к центру клетки, пигменты других хроматофоров подвергаются воздействию света, и кожа приобретает свой оттенок. Аналогичным образом, после агрегации меланина в DCU кожа становится зеленой из-за ксантофорной (желтой) фильтрации рассеянного света от слоя иридофора. При рассеивании меланина свет больше не рассеивается, и кожа становится темной. Поскольку другие биохроматические хроматофоры также способны к перемещению пигмента, животные с несколькими типами хроматофора могут генерировать впечатляющее множество цветов кожи, хорошо используя эффект разделения.[18][19]

Контроль и механика быстрой транслокации пигмента хорошо изучены у ряда различных видов, в частности у амфибий и костистость рыбы.[11][20] Было продемонстрировано, что процесс может быть гормональный или же нейронный контроль или и то, и другое, и для многих видов костистых рыб известно, что хроматофоры могут напрямую реагировать на внешние раздражители, такие как видимый свет, УФ-излучение, температура, pH, химические вещества и т. д.[21] Нейрохимические вещества, которые, как известно, перемещают пигмент, включают: норадреналин через его рецептор на поверхности на меланофорах.[22] Первичные гормоны, участвующие в регуляции транслокации, по-видимому, являются меланокортины, мелатонин, и меланин-концентрирующий гормон (MCH), которые вырабатываются в основном гипофизом, шишковидной железой и гипоталамусом соответственно. Эти гормоны также могут вырабатываться в паракринный мода клетками кожи. Было показано, что на поверхности меланофора гормоны активируют определенные Рецепторы, связанные с G-белком которые, в свою очередь, передают сигнал в клетку. Меланокортины вызывают диспергирование пигмента, а мелатонин и MCH вызывают агрегацию.[23]
У рыб были обнаружены многочисленные рецепторы меланокортина, МСН и мелатонина.[24] и лягушки,[25] включая гомолог из MC1R,[26] рецептор меланокортина, регулирующий кожа и цвет волос в людях.[27] Было продемонстрировано, что MC1R требуется рыбкам данио для диспергирования меланина.[28] Внутри клетки циклический аденозинмонофосфат (цАМФ) оказался важным второй посланник транслокации пигмента. Через механизм, еще не полностью изученный, цАМФ влияет на другие белки, такие как протеинкиназа А водить молекулярные моторы переносящий пигмент, содержащий пузырьки, вдоль обоих микротрубочки и микрофиламенты.[29][30][31]
Фоновая адаптация

Большинство рыб, рептилий и земноводных претерпевают ограниченное физиологическое изменение цвета в ответ на изменение окружающей среды. Этот вид камуфляжа, известный как фоновая адаптация, чаще всего проявляется в виде легкого потемнения или осветления тона кожи примерно до имитировать оттенок ближайшего окружения. Было продемонстрировано, что процесс фоновой адаптации зависит от зрения (похоже, животное должно видеть окружающую среду, чтобы адаптироваться к ней),[32] и что транслокация меланина в меланофоры является основным фактором изменения цвета.[23] Некоторые животные, такие как хамелеоны и анолис, имеют высокоразвитую фоновую адаптационную реакцию, способную очень быстро генерировать различные цвета. Они адаптировали способность изменять цвет в зависимости от температуры, настроения, уровня стресса и социальных сигналов, а не просто имитировать окружающую среду.
Разработка

Во время позвоночных эмбриональное развитие, хроматофоры - это один из многих типов клеток, образующихся в нервный гребень, парная полоса клеток, возникающая на краях нервная трубка. Эти клетки обладают способностью мигрировать на большие расстояния, позволяя хроматофорам заселять многие органы тела, включая кожу, глаза, ухо и мозг. Было обнаружено, что меланофоры и иридофоры рыб содержат регуляторные белки гладкой мускулатуры [кальпонин] и Caldesmon.[33] Покидая нервный гребень волнообразно, хроматофоры проходят дорсолатеральный путь через дерму, попадая в эктодерма через небольшие дыры в базальная пластинка, или вентромедиальный путь между сомиты и нервная трубка. Исключением являются меланофоры пигментированного эпителия сетчатки глаза. Они не происходят от нервного гребня. Вместо этого, выпячивание нервной трубки генерирует оптический стакан, что, в свою очередь, образует сетчатка.[нужна цитата ]
Когда и как мультипотентный клетки-предшественники хроматофоров (называемые хроматобласты) развиваются в свои дочерние подтипы - область постоянных исследований. У эмбрионов рыбок данио известно, например, что через 3 дня после оплодотворение каждый из классов клеток взрослых рыб - меланофоры, ксантофоры и иридофоры - уже присутствует. Исследования с использованием мутантных рыб показали, что факторы транскрипции Такие как Комплект, sox10, и митф важны для контроля дифференциации хроматофора.[34] Если эти белки являются дефектными, хроматофоры могут отсутствовать частично или полностью, что приводит к лейкистический беспорядок.
Практическое применение
Хроматофоры иногда используются в прикладных исследованиях. Например, личинки рыбок данио используются для изучения того, как хроматофоры организуются и взаимодействуют, чтобы точно сформировать регулярный узор с горизонтальными полосами, как у взрослых рыб.[35] Это рассматривается как полезный модель система для понимания паттернов в эволюционная биология развития поле. Биология хроматофора также использовалась для моделирования состояния или заболевания человека, включая меланома и альбинизм. В последнее время ген, ответственный за меланофор-специфический золотой штамм рыбок данио, Slc24a5, как было показано, имеет человеческий эквивалент, который сильно коррелирует с цвет кожи.[36]
Хроматофоры также используются как биомаркер слепоты у хладнокровных видов, так как животные с определенными дефектами зрения не могут фоново адаптироваться к световой среде.[32] Считается, что человеческие гомологи рецепторов, которые опосредуют перемещение пигмента в меланофорах, участвуют в таких процессах, как аппетит подавление и дубление, что делает их привлекательными целями для наркотики.[26] Поэтому фармацевтические компании разработали биологический анализ для быстрой идентификации потенциальных биологически активных соединений с использованием меланофоров из Африканская когтистая лягушка.[37] Другие ученые разработали методы использования меланофоров в качестве биосенсоры,[38] и для быстрого обнаружения болезней (на основании открытия, что токсин коклюша блокирует агрегацию пигментов в меланофорах рыб).[39] Потенциал военный были предложены варианты применения изменения цвета, опосредованного хроматофором, в основном как тип активный камуфляж, который мог бы как в каракатица сделать объекты почти невидимыми.[40][41]
Хроматофоры головоногих моллюсков

Колеид головоногие моллюски (включая осьминогов, кальмары и каракатица ) имеют сложные многоклеточные органы, которые они используют для быстрого изменения цвета, создавая большое разнообразие ярких цветов и узоров. Каждая хроматофорная единица состоит из одной хроматофорной клетки и множества мышц, нервов, глиальный, и клетки оболочки.[42] Внутри клетки хроматофора гранулы пигмента заключены в эластичный мешок, называемый цитоэластическим саккулюсом. Чтобы изменить цвет, животное искажает форму или размер саккулюса мышечным сокращением, изменяя его полупрозрачность, отражательная способность или непрозрачность. Это отличается от механизма, используемого у рыб, амфибий и рептилий, в том, что изменяется форма саккулюса, а не перемещение пигментных пузырьков внутри клетки. Однако достигается аналогичный эффект.
Осьминоги и большинство каракатица[43] может управлять хроматофорами в сложных волнообразных хроматических дисплеях, что приводит к множеству быстро меняющихся цветовых схем. Считается, что нервы, управляющие хроматофорами, расположены в головном мозге по схеме, изоморфной структуре хроматофоров, каждый из которых они контролируют. Это означает, что шаблон изменения цвета функционально соответствует шаблону активация нейронов. Это может объяснить, почему, когда нейроны активируются в каскаде повторяющихся сигналов, можно наблюдать волны изменения цвета.[44] Подобно хамелеонам, головоногие моллюски используют физиологические изменение цвета для социального взаимодействия. Они также являются одними из самых опытных в маскировке, имея возможность сочетать как цветовое распределение, так и текстура их местной среды с поразительной точностью.
Смотрите также
Примечания
- ^ Скотт М. Бобак и Линн М. Сиферман (2010). "Различия в окраске и изменение окраски у островных и материковых удавов (Удав)". Журнал герпетологии. 44 (4): 506–515. Дои:10.1670/09-026.1.
- ^ Аристотель. Historia Animalium. IX, 622а: 2-10. Около 400 г. до н. Э. Цитируется у Лучианы Боррелли, Франческа Герарди, Грациано Фиорито. Каталог рисунков тела головоногих моллюсков. Издательство Firenze University Press, 2006. Абстрактный Книги Google
- ^ Сангиованни, G (1819 г.). "Descrizione di un specificolare sistema di organi cromoforo espansivo-dermoideo e dei fenomeni che esso producti, scoperto nei molluschi cefaloso". G. Enciclopedico Napoli. 9: 1–13.
- ^ Дарвин, Чарльз (1860 г.). «Глава 1. Повадки морского слизня и каракатицы». Журнал исследований естественной истории и геологии стран, посещенных во время кругосветного путешествия H.M.S. «Бигль» под командованием капитана Фитц Роя, Р.Н.. Джон Мюррей, Лондон. п. 7.
- ^ а б Багнара, JT (1966). Цитология и цитофизиология немеланофорных пигментных клеток. Международный обзор цитологии. 20. С. 173–205. Дои:10.1016 / S0074-7696 (08) 60801-3. ISBN 978-0-12-364320-9. PMID 5337298.
- ^ Фокс, DL. Биохромы животных и структурные цвета: физические, химические, распределительные и физиологические особенности цветных тел в животном мире. Калифорнийский университет Press, Беркли, 1976. ISBN 0-520-02347-1
- ^ Мацумото, Дж. (1965). «Исследования тонкой структуры и цитохимических свойств эритрофоров меченого хвоста», Xiphophorus helleri, с особым акцентом на их пигментные гранулы (птериносомы) ". J Cell Biol. 27 (3): 493–504. Дои:10.1083 / jcb.27.3.493. ЧВК 2106771. PMID 5885426.
- ^ Bagnara JT. Сравнительная анатомия и физиология пигментных клеток в тканях не млекопитающих. В: Пигментная система: физиология и патофизиология, Oxford University Press, 1998. ISBN 0-19-509861-7
- ^ Тейлор, JD. (1969). «Влияние интермедина на ультраструктуру иридофоров амфибий». Gen Comp Endocrinol. 12 (3): 405–16. Дои:10.1016/0016-6480(69)90157-9. PMID 5769930.
- ^ Моррисон, Р. (1995). «Метод просвечивающей электронной микроскопии (ПЭМ) для определения структурных цветов, отраженных иридофорами ящериц». Пигментные клетки Res. 8 (1): 28–36. Дои:10.1111 / j.1600-0749.1995.tb00771.x. PMID 7792252.
- ^ а б c d Фуджи Р. (2000). «Регуляция подвижной активности хроматофоров рыб». Пигментные клетки Res. 13 (5): 300–19. Дои:10.1034 / j.1600-0749.2000.130502.x. PMID 11041206.
- ^ Ито, S; Вакамацу, К. (2003). «Количественный анализ эумеланина и феомеланина у людей, мышей и других животных: сравнительный обзор». Пигментные клетки Res. 16 (5): 523–31. Дои:10.1034 / j.1600-0749.2003.00072.x. PMID 12950732.
- ^ Bagnara, J.T .; Тейлор, JD; Прота, Г. (1973). «Изменение цвета, необычные меланосомы и новый пигмент от листовых лягушек». Наука. 182 (4116): 1034–5. Дои:10.1126 / science.182.4116.1034. PMID 4748673.
- ^ Багнара, Дж. (2003). "Загадки Птерорходина, красного меланосомного пигмента древесных лягушек". Исследование пигментных клеток. 16 (5): 510–516. Дои:10.1034 / j.1600-0749.2003.00075.x. PMID 12950730.
- ^ Schwalm, PA; Старрет, PH; McDiarmid, RW (1977). «Инфракрасное отражение у неотропических лягушек, сидящих на листьях». Наука. 196 (4295): 1225–7. Дои:10.1126 / science.860137. PMID 860137. S2CID 45014819.
- ^ Года М., Охата М., Икома Х, Фудзиёси Ю., Сугимото М., Фуджи Р. (2011). «Красновато-фиолетовая окраска кожных покровов из-за новых дихроматических хроматофоров костистых рыб Pseudochromis diadema». Пигментная клетка Melanoma Res. 24 (4): 614–7. Дои:10.1111 / j.1755-148X.2011.00861.x. PMID 21501419.
- ^ Bagnara, JT; Тейлор, JD; Хэдли, Мэн (1968). «Дермальный хроматофор». J Cell Biol. 38 (1): 67–79. Дои:10.1083 / jcb.38.1.67. ЧВК 2107474. PMID 5691979.
- ^ Палаццо, RE; Линч, TJ; Lo, SJ; Тейлор, JD; Tchen, TT (1989). «Перестройки птериносом и цитоскелета, сопровождающие дисперсию пигмента в ксантофорах золотой рыбки». Цитоскелет клеточного мотиля. 13 (1): 9–20. Дои:10.1002 / см. 970130103. PMID 2543509.
- ^ Porras, MG; Де Луф, А; Брейер, М; Аречига, Х (2003). "Procambarus clarkii". Пептиды. 24 (10): 1581–9. Дои:10.1016 / j.peptides.2003.08.016. PMID 14706537.
- ^ Дьякон, SW; Серпинская, АС; Vaughan, PS; Лопес Фанаррага, М; Вернос, I; Vaughan, KT; Гельфанд В.И. (2003). «Динактин необходим для двунаправленного транспорта органелл». Журнал клеточной биологии. 160 (3): 297–301. Дои:10.1083 / jcb.200210066. ЧВК 2172679. PMID 12551954.
- ^ Мейер-Рохов, В.Б. (2001). Хроматофоры рыб как сенсоры раздражителей окружающей среды - Книга = Сенсорная биология челюстей рыб; редакторы Капур Б.Г. и Хара Т.Дж .; Science Publishers Enfield (NH), США. С. 317–334.
- ^ Аспенгрен, S; Sköld, HN; Кирога, G; Mårtensson, L; Валлин, М (2003). «Норадреналин- и мелатонин-опосредованная регуляция агрегации пигментов в меланофорах рыб». Пигментные клетки Res. 16 (1): 59–64. Дои:10.1034 / j.1600-0749.2003.00003.x. PMID 12519126.
- ^ а б Логан, Д. В .; Ожог, SF; Джексон, Эй Джей (2006). «Регулирование пигментации меланофоров рыбок данио». Исследование пигментных клеток. 19 (3): 206–213. Дои:10.1111 / j.1600-0749.2006.00307.x. PMID 16704454.
- ^ Логан, DW; Брайсон-Ричардсон, RJ; Тейлор, MS; Currie, P; Джексон, Эй Джей (2003). «Последовательная характеристика рецепторов меланокортина костистых рыб». Ann N Y Acad Sci. 994: 319–30. Дои:10.1111 / j.1749-6632.2003.tb03196.x. PMID 12851332.
- ^ Sugden, D; Дэвидсон, К; Hough, KA; Тех, М.Т. (2004). «Мелатонин, рецепторы мелатонина и меланофоры: трогательная история». Пигментные клетки Res. 17 (5): 454–60. Дои:10.1111 / j.1600-0749.2004.00185.x. PMID 15357831.
- ^ а б Логан, DW; Брайсон-Ричардсон, RJ; Pagán, KE; Тейлор, MS; Карри, Полицейский; Джексон, Эй Джей (2003). «Структура и эволюция рецепторов меланокортина и MCH у рыб и млекопитающих». Геномика. 81 (2): 184–91. Дои:10.1016 / S0888-7543 (02) 00037-X. PMID 12620396.
- ^ Вальверде, П; Хили, E; Джексон, я; Рис, JL; Тоди, AJ (1995). «Варианты гена рецептора меланоцит-стимулирующего гормона связаны с рыжими волосами и светлой кожей у людей». Нат Жене. 11 (3): 328–30. Дои:10.1038 / ng1195-328. PMID 7581459.
- ^ Ричардсон, Дж; Лундегаард, PR; Рейнольдс, Нидерланды; Дорин-младший; Портеус, диджей; Джексон, Эй Джей; Паттон, EE (2008). «Путь mc1r регуляции дисперсии меланосом рыбок данио». Данио. 5 (4): 289–95. Дои:10.1089 / зеб.2008.0541. PMID 19133827.
- ^ Снайдер, Дж; Lin, F; Захеди, Н. Родионов, В; Ю, СС; Гросс, СП (2004). «Внутриклеточный транспорт на основе актина: насколько далеко вы продвинетесь, зависит от того, как часто вы переключаетесь». Труды Национальной академии наук Соединенных Штатов Америки. 101 (36): 13204–9. Дои:10.1073 / pnas.0403092101. ЧВК 516548. PMID 15331778.
- ^ Родионов В.И.; Надежда, Эй Джей; Свиткина, ТМ; Борисы, Г.Г. (1998). «Функциональная координация подвижности на основе микротрубочек и актина в меланофорах». Текущая биология. 8 (3): 165–8. Дои:10.1016 / S0960-9822 (98) 70064-8. PMID 9443917.
- ^ Кашина, АС; Семенова И.В. Иванов, П.А.; Потехина Е.С.; Заляпин, я; Родионов, В.И. (2004). «Протеинкиназа А, регулирующая внутриклеточный транспорт, образует комплексы с молекулярными моторами на органеллах». Текущая биология. 14 (20): 1877–81. Дои:10.1016 / j.cub.2004.10.003. PMID 15498498.
- ^ а б Neuhauss, SC. (2003). «Поведенческие генетические подходы к развитию и функционированию зрительной системы у рыбок данио» (PDF). J Neurobiol. 54 (1): 148–60. Дои:10.1002 / neu.10165. PMID 12486702.[мертвая ссылка ]
- ^ Meyer-Rochow, VB; Ройэла, М. (2002). «Кальпонин, кальдесмон и хроматофоры: соединение гладких мышц». Microsc. Res. Технология. 58 (6): 504–513. Дои:10.1002 / jemt.10169. PMID 12242708.
- ^ Келш, Р.Н.; Шмид, Б; Эйзен, Дж. С. (2000). «Генетический анализ развития меланофоров у эмбрионов рыбок данио». Дев Биол. 225 (2): 277–93. Дои:10.1006 / dbio.2000.9840. PMID 10985850.
- ^ Келш, Р.Н. (2004). «Генетика и эволюция пигментных паттернов у рыб». Исследование пигментных клеток. 17 (4): 326–36. Дои:10.1111 / j.1600-0749.2004.00174.x. PMID 15250934. S2CID 15112204.
- ^ Ламасон, Р.Л .; Мохидин, Массачусетс; Mest, JR; Вонг, AC; Нортон, HL; Арос, MC; Юринец, MJ; Мао, X; и другие. (2005). «SLC24A5, предполагаемый катионообменник, влияет на пигментацию у рыбок данио и людей». Наука. 310 (5755): 1782–6. Дои:10.1126 / science.1116238. PMID 16357253. S2CID 2245002.
- ^ Jayawickreme, CK; Sauls, H; Bolio, N; Руан, Дж; Мойер, М; Burkhart, W; Marron, B; Римеле, Т; Шаффер, Дж (1999). «Использование клеточного анализа в формате газона для быстрого скрининга библиотеки пептидов на основе 442 368 гранул». J Pharmacol Toxicol Методы. 42 (4): 189–97. Дои:10.1016 / S1056-8719 (00) 00083-6. PMID 11033434.
- ^ Андерссон, Т.П .; Филиппини, Д; Суська, А; Йоханссон, TL; Свенссон, ИП; Лундстрем, я (2005). «Лягушачьи меланофоры, культивируемые на флуоресцентных микрошариках: биосенсинг на основе биомимики». Биосенс Биоэлектрон. 21 (1): 111–20. Дои:10.1016 / j.bios.2004.08.043. PMID 15967358.
- ^ Karlsson, JO; Андерссон, Р.Г.; Askelöf, P; Elwing, H; Гранстрем, М; Grundström, N; Лундстрем, I; Оман, Л. (1991). «Агрегационная реакция меланофоров изолированной рыбьей чешуи: очень быстрый и точный диагноз коклюша». FEMS Microbiol Lett. 66 (2): 169–75. Дои:10.1111 / j.1574-6968.1991.tb04860.x. PMID 1936946.
- ^ Хансфорд, Дэйв (6 августа 2008 г.). «Каракатицы меняют цвет, меняют форму, чтобы ускользнуть от хищников». Национальная география Новости. Веллингтон, Новая Зеландия.
[...] каракатицы вместо этого полагались на невидимость, талант, который может найти применение в человеческих технологиях. Норман сказал, что военные проявили интерес к камуфляжу для каракатиц с целью однажды включить аналогичные механизмы в солдатскую форму.
- ^ Ли И. Нанотрубки для обработки зашумленных сигналов Кандидатская диссертация. 2005; Университет Южной Калифорнии.
- ^ Клони, РА; Флори, Э (1968). «Ультраструктура хроматофорных органов головоногих моллюсков». Zeitschrift für Zellforschung und Mikroskopische Anatomie. 89 (2): 250–80. Дои:10.1007 / BF00347297. PMID 5700268.
- ^ Лаан, А; Гутник, Т; Куба, MJ; Лоран, Г. (2014). «Поведенческий анализ бегущих волн каракатицы и его значение для нейронного контроля». Текущая биология. 24 (15): 1737–1742. Дои:10.1016 / j.cub.2014.06.027. PMID 25042589.
- ^ Демски, LS (1992). «Хроматофорные системы костистых и головоногих моллюсков: анализ конвергентных систем с ориентацией на уровни». Мозг, поведение и эволюция. 40 (2–3): 141–56. Дои:10.1159/000113909. PMID 1422807.
внешняя ссылка
- «Палитра природы - как животные производят цвет» (PDF). (1,20 МБ)
- Видеозапись адаптации фона осьминога
- Видеозапись формирования паттерна хроматофора кальмаров
- Веб-проект "Древо жизни": Хроматофоры головоногих моллюсков

Головоногие моллюски анатомия | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ракушка |
|    | ||||||||||||
| Мантия & воронка |
| |||||||||||||
| Голова & конечности |
| |||||||||||||
| Общий | ||||||||||||||
Стадии развития: Порождать → Параларва (Стадия доратопса ) → Ювенильный → Подростковый → Взрослый • Окаменелости яиц • Протоконх (зародышевый панцирь) | ||||||||||||||