Морские вирусы - Marine viruses

| Часть серии по |
| Планктон |
|---|
 |
Морские вирусы определяются их средой обитания как вирусы которые находятся в морская среда, то есть в соленая вода морей или океанов или солоноватый вода прибрежных эстуарии. Вирусы маленькие инфекционные агенты это может только копировать внутри жизни клетки из хозяин организм, потому что для этого им нужна репликационная машина хоста.[4] Они могут заразить все виды формы жизни, от животные и растения к микроорганизмы, в том числе бактерии и археи.[5]
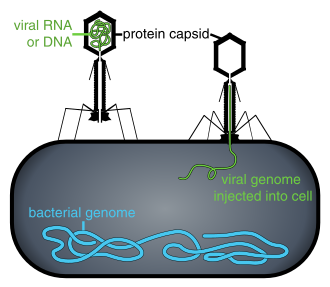
Находясь вне клетки или в процессе ее заражения, вирусы существуют в виде независимых частиц, называемых вирионы. Вирион содержит геном (длинная молекулы которые несут генетическую информацию в виде ДНК или РНК ) в окружении капсид (а белок шерсть, защищающая генетический материал). Формы этих вирусных частиц варьируются от простых спиральный и икосаэдр формы для одних видов вирусов в более сложные структуры для других. У большинства видов вирусов вирионы слишком малы, чтобы их можно было увидеть оптический микроскоп. Средний вирион составляет примерно одну сотую линейного размера среднего вириона. бактерия.
Чайная ложка морской воды обычно содержит около десяти миллионов морских вирусов. Большинство этих вирусов бактериофаги которые заражают и уничтожают морские бактерии и контролировать рост фитопланктон в основе морская пищевая сеть. Бактериофаги безвредны для растений и животных, но необходимы для регулирования морских экосистем. Они поставляют ключевые механизмы для переработка углерода океана и питательные вещества. В процессе, известном как вирусный шунт, органические молекулы, высвобождаемые из мертвых бактериальных клеток, стимулируют рост свежих бактерий и водорослей. В частности, разрушение бактерий вирусами (лизис ) было показано, что усиливает круговорот азота и стимулировать рост фитопланктона. Вирусная активность также влияет на биологический насос, процесс, который секвестр углерода в глубоком океане. Увеличивая количество дыхательных путей в океанах, вирусы косвенно ответственны за снижение количества углекислого газа в атмосфере примерно на 3%. гигатонны углерода в год
Морские микроорганизмы составляют около 70% от общего количества морская биомасса. Подсчитано, что морские вирусы убивают 20% этой биомассы каждый день. Вирусы - главные агенты, ответственные за быстрое уничтожение вредных цветение водорослей которые часто убивают других морская жизнь. Количество вирусов в океанах уменьшается дальше от берега и глубже в воду, где меньше организмов-хозяев. Вирусы - важное естественное средство передача генов между разными видами, что увеличивает генетическое разнообразие и движет эволюцией. Считается, что вирусы играли центральную роль в ранней эволюции до появления разнообразия бактерий, архей и эукариот, во времена последнего универсального общего предка жизни на Земле. Вирусы по-прежнему являются одной из крупнейших областей неизученного генетического разнообразия на Земле.
Задний план


Вирусы теперь признаны древними, и их происхождение предшествовало расхождению жизни в три домена.[6] Они встречаются везде, где есть жизнь, и, вероятно, существовали с момента появления живых клеток.[7] Происхождение вирусов в эволюционная история жизни неясны, потому что они не образуют окаменелости. Молекулярные методы используются для сравнения ДНК или РНК вирусов и являются полезным средством исследования того, как они возникли.[8] Некоторые вирусы могут иметь развился от плазмиды - фрагменты ДНК, которые могут перемещаться между клетками - в то время как другие могли развиться из бактерий. В эволюции вирусы являются важным средством горизонтальный перенос генов, что увеличивает генетическое разнообразие.[9]
Мнения расходятся относительно того, являются ли вирусы формой жизнь или органические структуры, взаимодействующие с живыми организмами.[10] Некоторые считают их формой жизни, потому что они несут генетический материал, воспроизводятся, создавая множество копий самих себя посредством самосборки, и развиваются через естественный отбор. Однако им не хватает ключевых характеристик, таких как клеточная структура, которая обычно считается необходимой для жизни. Поскольку они обладают некоторыми, но не всеми такими качествами, вирусы были описаны как репликаторы.[11] и как «организмы на краю жизни».[12]
Существование вирусов в океане было обнаружено благодаря электронная микроскопия и эпифлуоресцентная микроскопия экологических проб воды, а затем через метагеномный отбор образцов некультивируемых вирусов.[13][14] Морские вирусы, хотя и микроскопические, и практически незамеченные ученые до недавнего времени они были самыми многочисленными и разнообразными биологическими объектами в океане. Количество вирусов оценивается в 1030 в океане, или от 1 до 100 000x106 на миллилитр.[4] Количественная оценка морских вирусов первоначально проводилась с использованием просвечивающей электронной микроскопии, но была заменена эпифлуоресценцией или проточной цитометрии.[15]
Бактериофаги

Бактериофаги, часто просто звонят фаги, вирусы, которые паразит бактерии. Морские фаги паразит морские бактерии такие как цианобактерии.[16] Это разнообразная группа вирусов, которые являются наиболее многочисленными биологическими объектами в морской среде, потому что их хозяева, бактерии, как правило, численно доминируют в клеточной жизни в море. В океанах до десяти раз больше фагов, чем бактерий,[17] достигая уровня 250 миллионов бактериофагов на миллилитр морской воды.[18] Эти вирусы заражают определенные бактерии, связываясь с молекулы поверхностных рецепторов а затем вход в камеру. За короткий промежуток времени, в некоторых случаях всего за несколько минут, бактериальный полимераза начинает транслировать вирусную мРНК в белок. Эти белки становятся либо новыми вирионами внутри клетки, либо вспомогательными белками, которые помогают сборке новых вирионов, либо белками, участвующими в лизисе клетки. Вирусные ферменты способствуют разрушению клеточной мембраны, и есть фаги, которые могут реплицировать триста фагов через двадцать минут после инъекции.[19]
Бактерии защищаются от бактериофагов, производя ферменты, разрушающие чужеродную ДНК. Эти ферменты, называемые эндонуклеазы рестрикции, разрезать вирусную ДНК, которую бактериофаги вводят в бактериальные клетки.[20] Бактерии также содержат систему, которая использует CRISPR последовательности для сохранения фрагментов геномов вирусов, с которыми бактерии контактировали в прошлом, что позволяет им блокировать репликацию вируса посредством формы РНК-интерференция.[21][22] Эта генетическая система обеспечивает бактерии приобретенный иммунитет к инфекции.[23]
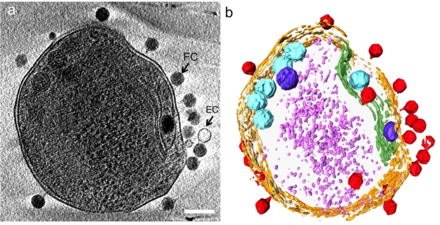
(б) то же изображение, визуализированное путем выделения клеточной стенки оранжевым цветом, плазматической мембраны светло-желтым, тилакоидной мембраны зеленым, карбоксисом голубым, полифосфатного тела синим, адсорбированных фагов по бокам или верхней части клетки красным цветом, и цитоплазматические гранулы (вероятно, в основном рибосомы) светло-фиолетового цвета.[24]

взаимодействие с морской Прохлорококк Бактерия MED4
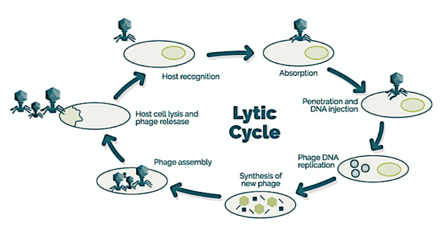
→ прикрепление: фаг прикрепляется к поверхности клетки-хозяина
→ проникновение: фаг вводит свою ДНК через клеточную мембрану
→ транскрипция: ДНК клетки-хозяина разрушается и метаболизм клетки
направлено на инициирование биосинтеза фага
→ биосинтез: ДНК фага реплицируется внутри клетки
→ созревание: реплицированный материал собирается в полностью сформированные вирусные фаги
→ лизис: новообразованные фаги высвобождаются из инфицированной клетки
(который сам разрушается в процессе) для поиска новых клеток-хозяев [25]
Микробы управляют преобразованием питательных веществ, которые поддерживают экосистемы Земли,[26] и вирусы, которые инфицируют эти микробы, модулируют как размер микробной популяции, так и ее разнообразие.[27][24] В цианобактерии Прохлорококк, самый распространенный оксигенный фототроф на Земле, вносит значительную долю в мировое производство первичного углерода и часто достигает плотности более 100000 клеток на миллилитр в олиготрофный и океаны умеренного пояса.[28] Следовательно, вирусная (цианофагальная) инфекция и лизис Прохлорококк представляют собой важную составляющую глобального цикл углерода. В дополнение к своей экологической роли в индукции смертности хозяина, цианофаги влияют на метаболизм и эволюцию своих хозяев, кооптируя и обмениваясь генами, включая основные гены фотосинтеза.[24]


В течение долгого времени, хвостатые фаги порядка Caudovirales казалось, что они доминируют в морских экосистемах по количеству и разнообразию организмов.[16] Однако в результате более поздних исследований оказалось, что нехвостые вирусы доминируют на многих глубинах и в океанических регионах.[29] Эти нехвостые фаги также инфицируют морские бактерии и включают семейства Corticoviridae,[30] Inoviridae,[31] Микровирусы [32] и Autolykiviridae.[33][34][35][36]
Вирусы архей
Вирусы архей воспроизводятся внутри археи: это двухцепочечные ДНК-вирусы необычной, а иногда и уникальной формы.[37][38] Эти вирусы наиболее подробно изучены в теплолюбивый архей, особенно отряды Sulfolobales и Thermoproteales.[39] Защита от этих вирусов включает вмешательство РНК со стороны повторяющаяся ДНК последовательности в геномах архей, которые связаны с генами вирусов.[40][41] Большинство архей имеют системы CRISPR – Cas в качестве адаптивной защиты от вирусов. Это позволяет архее сохранять участки вирусной ДНК, которые затем используются для нацеливания и устранения последующих заражений вирусом с использованием процесса, аналогичного вмешательству РНК.[42]
Грибковые вирусы
Миковирусы, также известные как микофаги, представляют собой вирусы, поражающие грибы. Инфекция грибковых клеток отличается от инфицирования клеток животных. У грибов жесткая клеточная стенка из хитина, поэтому большинство вирусов могут попасть внутрь этих клеток только после травмы клеточной стенки.[43]
- Увидеть; Nerva, L .; Ciuffo, M .; Валлино, М .; Margaria, P .; Varese, G.C .; Gnavi, G .; Турина, М. (2016). «Множественные подходы к обнаружению и характеристике вирусных и плазмидных симбионтов из коллекции морских грибов». Вирусные исследования. 219: 22–38. Дои:10.1016 / j.virusres.2015.10.028. HDL:2318/1527617. PMID 26546154.
Вирусы эукариотов

Морские протисты
К 2015 году около 40 вирусов, поражающих морские протисты были выделены и исследованы, большинство из них - вирусы микроводорослей.[44] Геномы этих морских протист-вирусов очень разнообразны.[45][46] Морские водоросли могут быть заражены вирусами в семье Phycodnaviridae. Они большие (100–560 кб) двухцепочечные ДНК-вирусы с участием икосаэдр фасонные капсиды. К 2014 году в семействе было идентифицировано 33 вида, разделенных на шесть родов,[47][48] который принадлежит к супергруппе больших вирусов, известных как нуклеоцитоплазматические большие ДНК-вирусы. В 2014 году были опубликованы данные, свидетельствующие о наличии некоторых разновидностей Phycodnaviridae может заразить людей, а не только виды водорослей, как считалось ранее.[49] Большинство родов этого семейства попадают в клетку-хозяина через клеточный рецептор. эндоцитоз и реплицироваться в ядре.

Phycodnaviridae играют важную экологическую роль, регулируя рост и продуктивность своих водорослей-хозяев. Виды водорослей, такие Гетеросигма акашиво и род Хризохромулина может образовывать густые цветы, которые могут нанести ущерб рыболовству и привести к убыткам в аквакультуре.[50] Вирус гетеросигмы акашиво (HaV) было предложено использовать в качестве микробного агента для предотвращения повторения токсичных красных приливов, производимых этим видом водорослей.[51] В кокколитовирус Вирус Эмилиании Хаксли 86, гигантский двухцепочечный ДНК-вирус, заражает повсеместно кокколитофора Эмилиания Хаксли.[47][48] Этот вирус имеет один из крупнейших известных геномов среди морских вирусов.[52] Phycodnaviridae вызывают гибель и лизис пресноводных и морских водорослей, высвобождая органический углерод, азот и фосфор в воду, обеспечивая питательными веществами микробный цикл.[53]
Отношение количества вирусов к прокариотам, VPR, часто используется как индикатор взаимосвязи между вирусами и хозяевами. Исследования использовали VPR, чтобы косвенно сделать вывод о влиянии вируса на микробную продуктивность, смертность и биогеохимический цикл в морской среде.[54] Однако, делая эти приближения, ученые предполагают, что VPR составляет 10: 1, то есть медиана наблюдаемого VPR на поверхности океана.[54][17] Фактический VPR сильно варьируется в зависимости от местоположения, поэтому VPR может не являться точным показателем вирусной активности или численности при лечении.[54][55]
Морские беспозвоночные
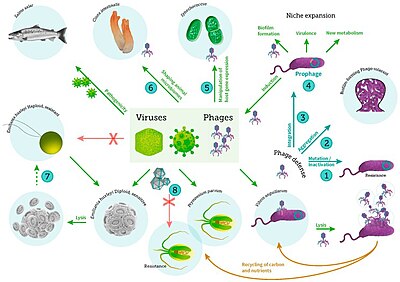
включая вирусные инфекции бактерий, фитопланктона и рыб[56]
Морские беспозвоночные подвержены вирусным заболеваниям.[57][58][59] Болезнь морской звезды это болезнь морская звезда и несколько других иглокожих, которые появляются спорадически, вызывая массовую смертность пораженных.[60] Это заболевание затронуло около 40 различных видов морских звезд. В 2014 году было высказано предположение, что болезнь связана с вирусом одноцепочечной ДНК, ныне известным как денсовирус, связанный с морскими звездами (SSaDV); тем не менее, болезнь, вызывающая истощение морских звезд, до конца не изучена.[61]
Морские позвоночные
Рыбы особенно подвержены инфекциям с рабдовирусы, которые отличаются от вируса бешенства, но связаны с ним. По крайней мере девять типов рабдовирусов вызывают экономически важные заболевания у таких видов, как лосось, щука, окунь, морской окунь, карп и треска. Симптомы включают анемию, кровотечение, летаргию и уровень смертности, на который влияет температура воды. В инкубаториях болезнь часто контролируется повышением температуры до 15–18 ° C.[62]:442–443 Как и все позвоночные, рыбы страдают от вирусы герпеса. Эти древние вирусы эволюционировали вместе со своими хозяевами и очень видоспецифичны.[62]:324 У рыб они вызывают раковые опухоли и незлокачественные образования, называемые гиперплазия.[62]:325
В 1984 г. инфекционная анемия лосося (ISAv) был обнаружен в Норвегии в Атлантический лосось инкубаторий. Восемьдесят процентов рыбы в очаге погибли. ISAv, вирусное заболевание, в настоящее время представляет собой серьезную угрозу для жизнеспособности разведения атлантического лосося.[63] Как следует из названия, это вызывает серьезные анемия инфицированных рыбы. В отличие от млекопитающих, красные кровяные тельца рыб имеют ДНК и могут заражаться вирусами. Стратегии управления включают разработку вакцины и повышение генетической устойчивости к болезни.[64]
морские млекопитающие также подвержены морским вирусным инфекциям. В 1988 и 2002 годах тысячи портовые тюлени были убиты в Европе вирус чумки фосин.[65] Многие другие вирусы, в том числе калицивирусы, герпесвирусы, аденовирусы и парвовирусы, циркулируют в популяциях морских млекопитающих.[66]
Гигантские морские вирусы
Большинство вирусов имеют длину от 20 до 300 нанометров. Это можно сравнить с длиной бактерий, которая начинается примерно с 400 нанометров. Это также гигантские вирусы, часто называют гирусыобычно около 1000 нанометров (одного микрона) в длину. Все гигантские вирусы относятся к филюм Nucleocytoviricota (NCLDV) вместе с поксвирусы Самый крупный из них - Тупанвирус. Этот вид гигантского вируса был обнаружен в 2018 году в глубоком океане, а также в содовом озере, и его общая длина может достигать 2,3 микрона.[67]

 Гигант мимивирус
Гигант мимивирус
Криоэлектронная микрофотография из CroV гигантский вирус [68]
масштабная линейка = 0,2 мкм


Открытие и последующая характеристика гигантских вирусов вызвали некоторые дискуссии относительно их эволюционного происхождения. Две основные гипотезы их происхождения заключаются в том, что либо они произошли от небольших вирусов, взяв ДНК из организмов-хозяев, либо они эволюционировали из очень сложных организмов в нынешнюю форму, которая не является самодостаточной для размножения.[69] От того, от какого сложного организма могли исходить гигантские вирусы, также ведутся споры. Одно из предположений состоит в том, что исходная точка фактически представляет четвертый домен жизни,[70][71] но это было в значительной степени сброшено со счетов.[72][73]
Вирофаги
Вирофаги представляют собой небольшие двухцепочечные ДНК-вирусы, которые полагаются на коинфекция из гигантские вирусы. Вирофаги полагаются на фабрику вирусной репликации соинфекционного гигантского вируса для своей собственной репликации. Одна из характеристик вирофагов заключается в том, что они имеют паразитический отношения с сопутствующим вирусом. Их зависимость от гигантского вируса для репликации часто приводит к дезактивации гигантских вирусов. Вирофаг может улучшить восстановление и выживаемость организма-хозяина. в отличие от других спутниковые вирусы, вирофаги имеют паразитический влияние на их соинфекционный вирус. Было замечено, что вирофаги делают гигантский вирус неактивным и тем самым улучшают состояние организма-хозяина.
Все известные вирофаги сгруппированы в семейство Lavidaviridae (от "большой вирус-зависимый или связанный" + -viridae ).[74] Первый вирофаг был обнаружен в градирни в Париже, Франция, в 2008 году. Он был обнаружен вместе с гигантским вирусом, заражающим его, Acanthamoeba castellanii мамавирус (ACMV). Вирофаг был назван Спутник и его репликация полностью зависела от коинфекции ACMV и его цитоплазматического репликационного аппарата. Было также обнаружено, что Sputnik оказывает ингибирующее действие на ACMV и улучшает выживаемость хозяина. Другие охарактеризованные вирофаги включают Спутник 2, Спутник 3, Замилон и Мавирус.[75][76]
Большинство этих вирофагов было обнаружено путем анализа метагеномный наборы данных. В метагеномном анализе последовательности ДНК обрабатываются множеством биоинформатических алгоритмов, которые выявляют определенные важные закономерности и характеристики. В этих наборах данных есть гигантские вирусы и вирофаги. Их разделяют последовательности от 17 до 20.kbp long, которые имеют сходство с уже секвенированными вирофагами. Эти вирофаги могут иметь линейные или кольцевые двухцепочечные геномы ДНК.[77] Вирофаги в культуре имеют икосаэдрические частицы капсида размером от 40 до 80 нанометров.[78] Частицы вирофагов настолько малы, что для их просмотра необходимо использовать электронную микроскопию. Анализ на основе метагеномных последовательностей был использован для прогнозирования около 57 полных и частичных геномов вирофагов.[79] а в декабре 2019 года - идентифицировать 328 высококачественных (полных или почти полных) геномов из различных мест обитания, включая кишечник человека, ризосферу растений и земные недра, из 27 различных таксономических клад.[80]



Гигантский морской вирус CroV заражает и вызывает смерть лизис морской зоофлагелляты Кафетерий roenbergensis.[82] Это влияет на экологию побережья, потому что Кафетерий roenbergensis питается бактериями, обнаруженными в воде. Когда мало Кафетерий roenbergensis из-за обширных инфекций CroV популяция бактерий увеличивается в геометрической прогрессии.[83] Влияние CroV на естественных популяциях С. roenbergensis остается неизвестным; однако было обнаружено, что вирус очень специфичен для хозяина и не заражает другие близкородственные организмы.[84] Cafeteria roenbergensis также инфицировано вторым вирусом, Мавирусный вирофаг при коинфекции CroV.[75] Этот вирус препятствует репликации CroV, что приводит к выживанию С. roenbergensis клетки. Мавирус способен интегрироваться в геном клеток С. roenbergensis и тем самым обеспечить неприкосновенность населения.[76]
Роль морских вирусов
Хотя морские вирусы только недавно были тщательно изучены, уже известно, что они играют важную роль во многих экосистемных функциях и циклах. Морские вирусы предлагают ряд важных экосистемные услуги и имеют важное значение для регулирования морских экосистем.[3] Морские бактериофаги и другие вирусы, по-видимому, влияют на биогеохимические циклы глобально, предоставлять и регулировать микробное биоразнообразие, круговорот углерода через морской пищевые сети, и необходимы для предотвращения бактериального демографические взрывы.[85]
Вирусный шунт

Преобладающими хозяевами вирусов в океане являются морские микроорганизмы, например бактерии.[13] Бактериофаги безвредны для растений и животных и необходимы для регулирования морских и пресноводных экосистем.[87] являются важными факторами смертности фитопланктон, основа пищевая цепочка в водных средах.[88] Они заражают и уничтожают бактерии в водных микробных сообществах и являются одним из важнейших механизмов переработка углерода круговорот питательных веществ в морской среде. Органические молекулы, высвобождаемые из мертвых бактериальных клеток, стимулируют рост свежих бактерий и водорослей в процессе, известном как вирусный шунт.[89]
Таким образом, считается, что морские вирусы играют важную роль в круговороте питательных веществ, повышая эффективность биологический насос. Вирусы вызывают лизис живых клеток, то есть они разрушают клеточные мембраны. Это высвобождает такие соединения, как аминокислоты и нуклеиновых кислот, которые, как правило, перерабатываются у поверхности.
Вирусная активность также увеличивает способность биологического насоса секвестр углерода в глубоком океане.[66] Лизис высвобождает больше неудобоваримого богатого углеродом материала, подобного тому, который содержится в стенках клеток, который, вероятно, экспортируется в более глубокие воды. Таким образом, материал, который переносится вирусным шунтом в более глубокие воды, вероятно, более богат углеродом, чем материал, из которого он был получен.[90][91] Увеличивая количество дыхательных путей в океанах, вирусы косвенно ответственны за снижение количества углекислого газа в атмосфере примерно на три гигатонны углерода в год.[66] Было показано, что лизис бактерий вирусами также усиливает круговорот азота и стимулирует рост фитопланктона.[92]
В вирусный шунт путь - это механизм, предотвращающий (прокариотический и эукариотический ) морской микроб твердое органическое вещество (POM) от перехода вверх трофические уровни переработав их в растворенное органическое вещество (DOM), которые легко усваиваются микроорганизмами. Вирусное шунтирование помогает поддерживать разнообразие в микробной экосистеме, предотвращая доминирование одного вида морских микробов в микросреде.[93] РОВ, рециркулируемое путем вирусного шунтирования, сопоставимо с количеством, вырабатываемым другими основными источниками морского РОВ.[94]


Вирусы являются наиболее распространенным биологическим объектом в морской среде.[5] В среднем в чайной ложке морской воды их около десяти миллионов.[96] Большинство этих вирусов бактериофаги заражение гетеротрофных бактерий и цианофаги заражение цианобактерий.Вирусы легко заражают микроорганизмы в микробной петле из-за их относительной численности по сравнению с микробами.[97] Прокариотическая и эукариотическая смертность способствует повторному использованию углеродных питательных веществ за счет лизис клеток. Есть также свидетельства регенерации азота (особенно аммония). Эта переработка питательных веществ помогает стимулировать рост микробов.[98] До 25% первичной продукции фитопланктона в мировом океане может быть повторно использовано в микробной петле посредством вирусного шунтирования.[99]
Ограничение цветения водорослей
Микроорганизмы составляют около 70% морской биомассы.[100] Подсчитано, что вирусы убивают 20% этой биомассы каждый день, и что вирусов в океанах в 15 раз больше, чем бактерий и архей. Вирусы - главные агенты, ответственные за быстрое уничтожение вредных цветение водорослей,[101] которые часто убивают других морских обитателей.[102] Ученые изучают потенциал морских цианофаги использоваться для предотвращения или обращения вспять эвтрофикация. Количество вирусов в океанах уменьшается дальше от берега и глубже в воду, где меньше организмов-хозяев.[103]
Перенос генов
Морские бактериофаги часто содержат вспомогательные метаболические гены, гены, производные от хозяина, как считается, поддерживают репликацию вируса, дополняя метаболизм хозяина во время вирусной инфекции.[104] Эти гены могут влиять на несколько биогеохимических циклов, включая углерод, фосфор, серу и азот.[105][106][107][108]
Вирусы - важное естественное средство передача генов между разными видами, что увеличивает генетическое разнообразие и движет эволюцией.[9] Считается, что вирусы играли центральную роль в ранней эволюции, еще до того, как появились разнообразные бактерии, археи и эукариоты, во время последний универсальный общий предок жизни на Земле.[109] Вирусы по-прежнему являются одним из крупнейших резервуаров неизведанного генетического разнообразия на Земле.[103]
Морские места обитания
Вдоль побережья
Морские прибрежные ареалы находятся на стыке суши и океана. Вполне вероятно что РНК-вирусы играют важную роль в этих условиях.[110]
На поверхности океана

Среда обитания на поверхности моря находится на границе между атмосферой и океаном. В биопленочный среда обитания на поверхности океана является местом обитания поверхностных микроорганизмов, обычно называемых Neuston. Вирусы в микрослое, так называемые Вирионейстон, в последнее время стали интересны исследователям как загадочные биологические объекты в пограничных поверхностных слоях, которые могут иметь серьезные экологические последствия. Учитывая, что эта обширная граница раздела воздух-вода находится на пересечении основных процессов обмена воздух-вода, охватывающих более 70% глобальной площади поверхности, она, вероятно, будет иметь серьезные последствия для морские биогеохимические циклы, на микробная петля и газообмен, а также морская пищевая сеть структура, глобальное распространение переносимых по воздуху вирусов, происходящих из микрослоя морской поверхности, и здоровье человека.[111]
В толще воды
Активность морских вирусов представляет собой возможное объяснение парадокс планктона предложено Джордж Хатчинсон в 1961 г.[112] Парадокс планктона заключается в том, что многие виды планктона были идентифицированы в небольших регионах океана, где ограниченные ресурсы должны создавать конкурентное исключение, ограничивая количество сосуществующих видов.[112] Морские вирусы могут играть роль в этом эффекте, поскольку вирусная инфекция увеличивается по мере увеличения потенциального контакта с хозяевами.[4] Таким образом, вирусы могут контролировать популяции видов планктона, которые растут слишком многочисленными, что позволяет сосуществовать широкому разнообразию видов.[4]
В отложениях
Морские бактериофаги играют важную роль в глубокое море экосистемы. Есть между 5x1012 и 1x1013 фагов на квадратный метр в глубоководных отложениях, и их численность тесно коррелирует с количеством прокариоты найдено в отложениях. Они ответственны за гибель 80% прокариот, обнаруженных в отложениях, и почти все эти смерти вызваны клетками. лизис (разрываясь). Это позволяет превращать азот, углерод и фосфор из живых клеток в растворенные органические вещества и детрит, что способствует высокой скорости круговорота питательных веществ в глубоководных отложениях. Из-за важности глубоководных отложений в биогеохимических циклах морские бактериофаги влияют на углерод, азот и циклы фосфора. Для более точного выяснения этих влияний необходимо провести дополнительные исследования.[113]
В гидротермальных источниках
Вирусы являются частью микробное сообщество гидротермальных источников и их влияние на микробную экологию в этих экосистемах - растущая область исследований.[114] Вирусы - это самая многочисленная жизнь в океане, являющаяся самым большим резервуаром генетического разнообразия.[101] Поскольку их инфекции часто заканчиваются смертельным исходом, они представляют собой значительный источник смертности и, таким образом, имеют широкое влияние на биологические океанографические процессы. эволюция и биогеохимический цикл в океане.[103] Однако были обнаружены доказательства того, что вирусы, обнаруженные в жерловых средах обитания, приняли более мутуалистический чем паразитический эволюционной стратегии, чтобы выжить в экстремальной и нестабильной среде, в которой они существуют.[115] Было обнаружено, что в глубоководных гидротермальных жерлах содержится большое количество вирусов, что указывает на высокую вирусную продукцию.[116] Как и в других морских средах, глубоководные гидротермальные вирусы влияют на изобилие и разнообразие прокариоты и, следовательно, влияют на микробный биогеохимический цикл путем лизать их хозяева для репликации.[117] Однако, в отличие от их роли в качестве источника смертности и контроля над популяциями, вирусы также постулируются как повышающие выживаемость прокариот в экстремальных условиях окружающей среды, выступая в качестве резервуаров генетической информации. Следовательно, считается, что взаимодействие виросферы с микроорганизмами в условиях стресса окружающей среды способствует выживанию микроорганизмов за счет распространения генов хозяина через горизонтальный перенос генов.[118]
Полярные регионы
Распределение
Вирусы сильно зависят от хозяина.[119] Морской вирус с большей вероятностью заразит совместно встречающиеся организмы, живущие в том же регионе, где обитает вирус.[120] Следовательно, биогеография является важным фактором способности вириона инфицировать.
Знание об этой вариации вирусных популяций в зависимости от пространственно-временного и других градиентов окружающей среды подтверждается морфологией вируса, определенной с помощью просвечивающей электронной микроскопии (ПЭМ). Нехвостые вирусы, по-видимому, доминируют на многих глубинах и в океанических регионах, за ними следуют Caudovirales. миовирусы, подовирусы и сифовирусы.[29] Однако вирусы, принадлежащие к семействам Corticoviridae,[121] Inoviridae[122] и Микровирусы[123] также известно, что они заражают различные морские бактерии. Метагеномный Имеющиеся данные свидетельствуют о том, что микровирусы (фаги икосаэдрической оцДНК) особенно распространены в морских средах обитания.[123]
Метагеномные подходы к оценке вирусного разнообразия часто ограничены отсутствием эталонных последовательностей, в результате чего многие последовательности остаются без аннотации.[124] Однако вирусные контиги генерируются путем прямого секвенирования вирусной фракции, обычно генерируемой после фильтрации 0,02 мкм образца морской воды, или с помощью биоинформатических подходов для идентификации вирусных контигов или вирусных геномов из микробного метагенома. Новые инструменты для выявления предполагаемых вирусных контигов, такие как VirSorter[125] и VirFinder,[126] позволяют оценить характер вирусного обилия, круг хозяев и функциональное содержание морского бактериофага.[127][128]
Смотрите также
использованная литература
- ^ Bonnain, C .; Breitbart, M .; Бак, К. (2016). «Гипотеза ферроянского коня: взаимодействие железа и вируса в океане». Границы морских наук. 3: 82. Дои:10.3389 / fmars.2016.00082. S2CID 2917222.
- ^ Эти крошечные организмы имеют действительно странные формы National Geogaphic », 12 ноября 2016 г.
- ^ а б Шорс Т. (2008). Понимание вирусов. Джонс и Бартлетт Издательство. п. 5. ISBN 978-0-7637-2932-5.
- ^ а б c d Brussaard CP, Baudoux A, Rodríguez-Valera F (2016). Сталь ЖЖ, Кретою М.С. (ред.). Морские вирусы. Морской микробиом. Издательство Springer International. С. 155–183. Дои:10.1007/978-3-319-33000-6_5. ISBN 9783319329987.
- ^ а б c d Кунин Э.В., Сенкевич Т.Г., Доля В.В. (2006). «Древний вирусный мир и эволюция клеток». Биология Директ. 1: 29. Дои:10.1186/1745-6150-1-29. ЧВК 1594570. PMID 16984643.CS1 maint: несколько имен: список авторов (ссылка на сайт)
 Материал был скопирован из этого источника, который доступен под Международная лицензия Creative Commons Attribution 2.0.
Материал был скопирован из этого источника, который доступен под Международная лицензия Creative Commons Attribution 2.0. - ^ Mahy WJ и Van Regenmortel MHV (редакторы). Настольная энциклопедия общей вирусологии. Оксфорд: Academic Press; 2009 г. ISBN 0-12-375146-2. п. 28.
- ^ Айер Л. М., Баладжи С., Кунин Е. В., Аравинд Л. (2006). «Эволюционная геномика нуклео-цитоплазматических больших ДНК-вирусов». Вирусные исследования. 117 (1): 156–84. Дои:10.1016 / j.virusres.2006.01.009. PMID 16494962.CS1 maint: несколько имен: список авторов (ссылка на сайт)
- ^ Санхуан Р., Небот М. Р., Кирико Н., Мански Л. М., Белшоу Р. (октябрь 2010 г.). «Частота вирусных мутаций». Журнал вирусологии. 84 (19): 9733–48. Дои:10.1128 / JVI.00694-10. ЧВК 2937809. PMID 20660197.CS1 maint: несколько имен: список авторов (ссылка на сайт)
- ^ а б Canchaya C, Fournous G, Chibani-Chennoufi S, Dillmann ML, Brüssow H (2003). «Фаги как агенты латерального переноса генов». Текущее мнение в микробиологии. 6 (4): 417–24. Дои:10.1016 / S1369-5274 (03) 00086-9. PMID 12941415.CS1 maint: несколько имен: список авторов (ссылка на сайт)
- ^ Кунин Э.В., Старокадомский П (2016). «Живы ли вирусы? Парадигма репликатора проливает решающий свет на старый, но ошибочный вопрос». Исследования по истории и философии биологических и биомедицинских наук. 59: 125–134. Дои:10.1016 / j.shpsc.2016.02.016. ЧВК 5406846. PMID 26965225.
- ^ Кунин, Э. В .; Старокадомский П. (7 марта 2016 г.). «Живы ли вирусы? Парадигма репликатора проливает решающий свет на старый, но ошибочный вопрос». Исследования по истории и философии биологических и биомедицинских наук. 59: 125–34. Дои:10.1016 / j.shpsc.2016.02.016. ЧВК 5406846. PMID 26965225.
- ^ Рыбицки, EP (1990). «Классификация организмов на пороге жизни, или проблемы с вирусной систематикой». Южноафриканский научный журнал. 86: 182–186.
- ^ а б Манн Н.Х. (май 2005 г.). «Третий век фага». PLOS Биология. 3 (5): e182. Дои:10.1371 / journal.pbio.0030182. ЧВК 1110918. PMID 15884981.
- ^ Wommack KE, Hill RT, Muller TA, Colwell RR (апрель 1996 г.). «Влияние солнечного света на жизнеспособность и структуру бактериофагов». Прикладная и экологическая микробиология. 62 (4): 1336–41. Дои:10.1128 / AEM.62.4.1336-1341.1996. ЧВК 167899. PMID 8919794.
- ^ Мари Д., Брюссаард С.П., Тирхауг Р., Братбак Г., Воулот Д. (январь 1999 г.). «Подсчет морских вирусов в культуре и природных образцах методом проточной цитометрии». Прикладная и экологическая микробиология. 65 (1): 45–52. Дои:10.1128 / AEM.65.1.45-52.1999. ЧВК 90981. PMID 9872758.
- ^ а б Манн, Нью-Хэмпшир (2005-05-17). "Третий век фага". PLOS Биология. 3 (5): 753–755. Дои:10.1371 / journal.pbio.0030182. ЧВК 1110918. PMID 15884981.
- ^ а б Wommack KE, Colwell RR (март 2000 г.). «Вириопланктон: вирусы в водных экосистемах». Обзоры микробиологии и молекулярной биологии. 64 (1): 69–114. Дои:10.1128 / MMBR.64.1.69-114.2000. ЧВК 98987. PMID 10704475.
- ^ Bergh O, Børsheim KY, Bratbak G, Heldal M (август 1989 г.). «Большое количество вирусов, обнаруженных в водной среде». Природа. 340 (6233): 467–68. Bibcode:1989Натура.340..467Б. Дои:10.1038 / 340467a0. PMID 2755508. S2CID 4271861.
- ^ Шорс с. 595–97.
- ^ Бикл Т.А., Крюгер Д.Х. (июнь 1993 г.). «Биология рестрикции ДНК». Микробиологические обзоры. 57 (2): 434–50. Дои:10.1128 / MMBR.57.2.434-450.1993. ЧВК 372918. PMID 8336674.
- ^ Баррангу Р., Фремо С., Дево Х., Ричардс М., Боявал П., Муано С. и др. (Март 2007 г.). «CRISPR обеспечивает приобретенную устойчивость к вирусам у прокариот». Наука. 315 (5819): 1709–12. Bibcode:2007Научный ... 315.1709B. Дои:10.1126 / science.1138140. HDL:20.500.11794/38902. PMID 17379808. S2CID 3888761.
- ^ Brouns SJ, Jore MM, Lundgren M, Westra ER, Slijkhuis RJ, Snijders AP, et al. (Август 2008 г.). «Малые РНК CRISPR направляют противовирусную защиту прокариот». Наука. 321 (5891): 960–64. Bibcode:2008Sci ... 321..960B. Дои:10.1126 / science.1159689. ЧВК 5898235. PMID 18703739.
- ^ Mojica FJ, Родригес-Валера F (сентябрь 2016 г.). «Открытие CRISPR у архей и бактерий». Журнал FEBS. 283 (17): 3162–69. Дои:10.1111 / фев.13766. HDL:10045/57676. PMID 27234458.
- ^ а б c d Мурата, К .; Zhang, Q .; Galaz-Montoya, J.G .; Fu, C .; Coleman, M.L .; Osburne, M.S .; Schmid, M.F .; Салливан, МБ; Chisholm, S.W .; Чиу, В. (2017). «Визуализация адсорбции цианофага P-SSP7 на морских Прохлорококк". Научные отчеты. 7: 44176. Bibcode:2017НатСР ... 744176М. Дои:10.1038 / srep44176. ЧВК 5345008. PMID 28281671.
- ^ Как размножаются бактериофаги? Университет Барселоны. Проверено 12 июля 2020.
- ^ Фальковски, П.Г .; Fenchel, T .; Делонг, Э.Ф. (2008). «Микробные двигатели, управляющие биогеохимическими циклами Земли». Наука. 320 (5879): 1034–1039. Bibcode:2008Научный ... 320.1034F. Дои:10.1126 / science.1153213. PMID 18497287. S2CID 2844984.
- ^ Brum, J.R .; Салливан, М. (2015). «Принимая вызов: ускоренные темпы открытий меняют морскую вирусологию». Обзоры природы Микробиология. 13 (3): 147–159. Дои:10.1038 / nrmicro3404. PMID 25639680. S2CID 32998525.
- ^ Bouman, H.A .; Ulloa, O .; Scanlan, D.J .; Zwirglmaier, K .; Li, W.K .; Platt, T .; Стюарт, В .; Barlow, R .; Leth, O .; Clementson, L .; Лутц, В. (2006). «Океанографические основы глобального поверхностного распределения Прохлорококк экотипы ». Наука. 312 (5775): 918–921. Bibcode:2006Научный ... 312..918Б. Дои:10.1126 / science.1122692. PMID 16690867. S2CID 20738145.
- ^ а б Брам-младший, Шенк Р.О., Салливан МБ (сентябрь 2013 г.). «Глобальный морфологический анализ морских вирусов показывает минимальные региональные вариации и преобладание нехвостых вирусов». Журнал ISME. 7 (9): 1738–51. Дои:10.1038 / ismej.2013.67. ЧВК 3749506. PMID 23635867.
- ^ Крупович М., Бэмфорд Д.Х. (2007). «Предполагаемые профаги, связанные с литическим бесхвостым фагом морской дцДНК PM2, широко распространены в геномах водных бактерий». BMC Genomics. 8: 236. Дои:10.1186/1471-2164-8-236. ЧВК 1950889. PMID 17634101.
- ^ Сюэ Х, Сюй И, Буше И, Польз М.Ф. (2012). «Высокая частота нового нитчатого фага, VCYϕ, в популяции экологического холерного вибриона». Прикладная и экологическая микробиология. 78 (1): 28–33. Дои:10.1128 / AEM.06297-11. ЧВК 3255608. PMID 22020507.
- ^ Ру С., Крупович М., Пуле А., Деброа Д., Эно Ф. (2012). «Эволюция и разнообразие вирусного семейства Microviridae посредством коллекции из 81 нового полного генома, собранных из виромных прочтений». PLOS ONE. 7 (7): e40418. Bibcode:2012PLoSO ... 740418R. Дои:10.1371 / journal.pone.0040418. ЧВК 3394797. PMID 22808158.
- ^ Кауфман, Кэтрин М .; Хуссейн, Фатима А .; Ян, радость; Аревало, Филипп; Браун, Юлия М .; Чанг, Уильям К .; Ванинсберге, Дэвид; Эльшербини, Джозеф; Sharma, Radhey S .; Катлер, Майкл Б .; Келли, Либуша; Польз, Мартин Ф. (2018). «Основная линия нехвостых вирусов дцДНК как непризнанных убийц морских бактерий». Природа. 554 (7690): 118–122. Bibcode:2018Натура.554..118K. Дои:10.1038 / природа25474. PMID 29364876. S2CID 4462007.
- ^ Ученые обнаружили новый тип вируса в Мировом океане: Autolykiviridae, on: sci-news, 25 января 2018 г.
- ^ В океане только что обнаружили ранее неизвестные вирусы со странной ДНК, по: наукапредупреждение, 25 января 2018
- ^ NCBI: Autolykiviridae (семейство) - неклассифицированные вирусы дцДНК
- ^ Лоуренс С.М., Менон С., Эйлерс Б.Дж., Ботнер Б., Хаят Р., Дуглас Т., Янг М.Дж. (2009). «Структурно-функциональные исследования вирусов архей». Журнал биологической химии. 284 (19): 12599–603. Дои:10.1074 / jbc.R800078200. ЧВК 2675988. PMID 19158076.
- ^ Прангишвили Д., Фортер П, Гаррет Р. А. (2006). «Вирусы архей: объединяющая точка зрения». Обзоры природы Микробиология. 4 (11): 837–48. Дои:10.1038 / nrmicro1527. PMID 17041631. S2CID 9915859.CS1 maint: несколько имен: список авторов (ссылка на сайт)
- ^ Прангишвили Д., Гаррет Р.А. (2004). «Исключительно разнообразные морфотипы и геномы гипертермофильных вирусов кренархей». Сделки Биохимического Общества. 32 (Pt 2): 204–8. Дои:10.1042 / BST0320204. PMID 15046572.
- ^ Mojica FJ, Díez-Villaseñor C, García-Martínez J, Soria E (февраль 2005 г.). «Промежуточные последовательности регулярно расположенных прокариотических повторов происходят из чужеродных генетических элементов». Журнал молекулярной эволюции. 60 (2): 174–82. Bibcode:2005JMolE..60..174M. Дои:10.1007 / s00239-004-0046-3. PMID 15791728. S2CID 27481111.
- ^ Макарова К.С., Гришин Н.В., Шабалина С.А., Вольф Ю.И., Кунин Е.В. (март 2006 г.). «Предполагаемая иммунная система прокариот, основанная на РНК-интерференции: компьютерный анализ предсказанного ферментативного механизма, функциональные аналогии с эукариотической РНКи и гипотетические механизмы действия». Биология Директ. 1: 7. Дои:10.1186/1745-6150-1-7. ЧВК 1462988. PMID 16545108.
- ^ ван дер Ост Дж, Вестра ER, Джексон Р.Н., Виденхефт Б. (июль 2014 г.). «Раскрытие структурных и механических основ систем CRISPR-Cas». Обзоры природы Микробиология. 12 (7): 479–92. Дои:10.1038 / nrmicro3279. ЧВК 4225775. PMID 24909109.
- ^ Диммок Нью-Джерси, Истон А.Дж., Леппард К. (2007). Введение в современную вирусологию (Шестое изд.). Блэквелл Паблишинг. п.70. ISBN 978-1-4051-3645-7.
- ^ Томару Й, Кимура К. и Нагасаки К. (2015) «Вирусы морских протистов». В: Ohtsuka S, Suzaki T, Horiguchi T, Suzuki N, Not F (ред.) Морские протисты страницы 501–517. Спрингер, Токио. Дои:10.1007/978-4-431-55130-0_20. ISBN 978-4-431-55130-0.
- ^ Хайман, Пол; Абедон, Стивен Т. (2012). «Мелкие блохи: вирусы микроорганизмов». Scientifica. 2012: 1–23. Дои:10.6064/2012/734023. ЧВК 3820453. PMID 24278736.. Материал был скопирован из этого источника, который доступен под Международная лицензия Creative Commons Attribution 3.0.
- ^ Шорт, С. (2012). «Экология вирусов, поражающих эукариотические водоросли». Экологическая микробиология. 14 (9): 2253–2271. Дои:10.1111 / j.1462-2920.2012.02706.x. PMID 22360532.
- ^ а б «Вирусная зона». ExPASy. Получено 15 июн 2015.
- ^ а б ICTV. «Таксономия вирусов: выпуск 2014 г.». Получено 15 июн 2015.
- ^ Желток, RH; и другие. (2014). «Хлоровирус ATCV-1 является частью ротоглоточного вирома человека и связан с изменениями когнитивных функций у людей и мышей». Proc Natl Acad Sci U S A. 111 (45): 16106–16111. Bibcode:2014PNAS..11116106Y. Дои:10.1073 / pnas.1418895111. ЧВК 4234575. PMID 25349393.
- ^ Томару, Юдзи; Шираи, Йоко; Нагасаки, Кейдзо (1 августа 2008 г.). "Экология, физиология и генетика фикоднавируса, поражающего вредный цветущий рафидофит". Гетеросигма акашиво". Наука о рыболовстве. 74 (4): 701–711. Дои:10.1111 / j.1444-2906.2008.01580.x. S2CID 23152411.
- ^ Нагасаки, Кейдзо; Тарутани, Кенджи; Ямагути, Минео (1999-03-01). «Характеристики роста вируса Heterosigma akashiwo и его возможное использование в качестве микробиологического агента для борьбы с красным приливом». Прикладная и экологическая микробиология. 65 (3): 898–902. Дои:10.1128 / AEM.65.3.898-902.1999. ЧВК 91120. PMID 10049839.
- ^ Крупнейшие известные вирусные геномы Giantviruses.org. Дата обращения: 11 июня 2020 г.
- ^ Сиги, Дэвид (2005-09-27). Пресноводная микробиология: биоразнообразие и динамические взаимодействия микроорганизмов в водной среде. Джон Вили и сыновья. ISBN 9780470026472.
- ^ а б c Вигингтон СН, Сондреггер Д.Л., Брюссард С.П., Бучан А., Финке Дж.Ф., Фурман Дж., Леннон Дж. Т., Мидделбо М., Саттл, Калифорния, Сток C, Уилсон WH (2015-08-26). «Повторное изучение взаимосвязи между численностью вирусов и микробных клеток в мировом океане». bioRxiv: 025544. bioRxiv 10.1101/025544. Дои:10.1101/025544.
- ^ Парикка К.Дж., Le Romancer M, Wauters N, Jacquet S (май 2017 г.). «Расшифровка отношения вируса к прокариотам (VPR): понимание взаимоотношений вирус-хозяин в различных экосистемах». Биологические обзоры Кембриджского философского общества. 92 (2): 1081–1100. Дои:10.1111 / brv.12271. PMID 27113012. S2CID 3463306.
- ^ Middelboe, M .; Брюссар, К. (2017). «Морские вирусы: ключевые игроки в морских экосистемах». Вирусы. 9 (10): 302. Дои:10.3390 / v9100302. ЧВК 5691653. PMID 29057790.
- ^ Т. Джонсон, П. (1984). «Вирусные болезни морских беспозвоночных». Helgoländer Meeresuntersuchungen. 37 (1–4): 65–98. Bibcode:1984HM ..... 37 ... 65J. Дои:10.1007 / BF01989296. S2CID 30161955.
- ^ Renault T (2011 г.) «Вирусы, поражающие морских моллюсков» В: Hurst CJ (Ed) Исследования вирусной экологии, Том 2: Системы-хозяева животных, John Wiley & Sons. ISBN 9781118024584.
- ^ Арзул, И .; Corbeil, S .; Морга, Б .; Рено, Т. (2017). «Вирусы, поражающие морских моллюсков» (PDF). Журнал патологии беспозвоночных. 147: 118–135. Дои:10.1016 / j.jip.2017.01.009. PMID 28189502.
- ^ Доусони, Соластер. «Виды морских звезд, пораженные синдромом истощения». Pacificrockyintertidal.org Seastarwasting.org (n.d.): n. стр. Экология и эволюционная биология. Интернет.
- ^ "Синдром истощения морской звезды | МАРИН". eeb.ucsc.edu. Получено 2018-06-03.
- ^ а б c Мерфи, ФА; Гиббс, EPJ; Horzinek, MC; Студдарт MJ (1999). Ветеринарная вирусология. Бостон: Academic Press. ISBN 978-0-12-511340-3.
- ^ Нью-Брансуик поможет Чили победить болезнь Информация и услуги по рыбной ловле
- ^ Информационный бюллетень - Исследования аквакультуры атлантического лосося В архиве 29 декабря 2010 г. Wayback Machine Рыболовство и океаны Канады. Проверено 12 мая 2009 года.
- ^ Холл А.Дж., Джепсон П.Д., Гудман С.Дж., Харконен Т. (2006). «Вирус чумы фокинии в Северном и Европейском морях - данные и модели, природа и воспитание». Биологическое сохранение. 131 (2): 221–29. Дои:10.1016 / j.biocon.2006.04.008.
- ^ а б c Suttle CA (октябрь 2007 г.). «Морские вирусы - основные игроки в глобальной экосистеме». Обзоры природы Микробиология. 5 (10): 801–12. Дои:10.1038 / nrmicro1750. PMID 17853907. S2CID 4658457.
- ^ Абрахао, Йонатас; Сильва, Лорена; Сильва, Людмила Сантос; Халил, Жак Яакуб Боу; Родригес, Родриго; Арантес, Талита; Ассис, Фелипе; Боратто, Пауло; Андраде, Мигель; Крун, Эрна Геессиен; Рибейро, Бергманн; Бергье, Иван; Селигманн, Эрве; Гиго, Эрик; Колсон, Филипп; Левассер, Энтони; Кремер, Гвидо; Рауль, Дидье; Скола, Бернар Ла (27 февраля 2018 г.). «Хвостатый гигант Тупанвирус обладает наиболее полным трансляционным аппаратом из известной виросферы». Nature Communications. 9 (1): 749. Bibcode:2018НатКо ... 9..749A. Дои:10.1038 / s41467-018-03168-1. ЧВК 5829246. PMID 29487281.
- ^ Xiao, C .; Фишер, М.Г .; Bolotaulo, D.M .; Ulloa-Rondeau, N .; Avila, G.A .; Саттл, К.А. (2017). «Крио-ЭМ реконструкция капсида вируса Cafeteria roenbergensis предлагает новый путь сборки гигантских вирусов». Научные отчеты. 7 (1): 5484. Bibcode:2017НатСР ... 7.5484X. Дои:10.1038 / s41598-017-05824-w. ЧВК 5511168. PMID 28710447.
- ^ Bichell RE. «В гигантских вирусных генах намекает на их загадочное происхождение». Все учтено.
- ^ Ван Эттен Дж. Л. (июль – август 2011 г.). «Гигантские вирусы». Американский ученый. 99 (4): 304–311. Дои:10.1511/2011.91.304.
- ^ Лежандр М., Арслан Д., Абергель С., Клавери Дж. М. (январь 2012 г.). «Геномика мегавируса и неуловимая четвертая область жизни». Коммуникативная и интегративная биология. 5 (1): 102–6. Дои:10.4161 / cib.18624. ЧВК 3291303. PMID 22482024.
- ^ Шульц Ф., Ютин Н., Иванова Н. Н., Ортега Д. Р., Ли Т. К., Вирхейлиг Дж., Даймс Х., Хорн М., Вагнер М., Дженсен Г. Дж., Кирпидес Н. С., Кунин Е. В., Войке Т. (апрель 2017 г.). «Гигантские вирусы с расширенным набором компонентов системы перевода» (PDF). Наука. 356 (6333): 82–85. Bibcode:2017Научный ... 356 ... 82S. Дои:10.1126 / science.aal4657. PMID 28386012. S2CID 206655792.
- ^ Бэкстрём Д., Ютин Н., Йоргенсен С.Л., Дхарамши Дж., Хома Ф., Заремба-Недведска К., Спанг А., Вольф Ю.И., Кунин Е.В., Эттема Т.Дж. (март 2019 г.). «Геномы вирусов из глубоководных отложений расширяют океанический мегавиром и поддерживают независимое происхождение вирусного гигантизма». мБио. 10 (2): e02497-02418. Дои:10,1128 / мБио.02497-18. ЧВК 6401483. PMID 30837339.
- ^ Duponchel, S; Фишер, MG (март 2019 г.). «Да здравствуют лавидавирусы! Пять особенностей вирофагов, паразитирующих на гигантских ДНК-вирусах». Патогены PLOS. 15 (3): e1007592. Дои:10.1371 / journal.ppat.1007592. ЧВК 6428243. PMID 30897185.
- ^ а б Фишер М.Г., Саттл, Калифорния (апрель 2011 г.). «Вирофаг в происхождении больших транспозонов ДНК». Наука. 332 (6026): 231–4. Bibcode:2011Наука ... 332..231F. Дои:10.1126 / science.1199412. PMID 21385722. S2CID 206530677.
- ^ а б Фишер М.Г., Хакл (декабрь 2016 г.). «Интеграция генома хозяина и гигантская вирус-индуцированная реактивация мавируса вирофага». Природа. 540 (7632): 288–91. Bibcode:2016Натура. 540..288F. Дои:10.1038 / природа20593. PMID 27929021. S2CID 4458402.
- ^ Кацуракис, Арис; Асвад, Амр (2014). «Происхождение гигантских вирусов, вирофагов и их родственников в геномах хозяина». BMC Биология. 12: 2–3. Дои:10.1186 / s12915-014-0051-у. ЧВК 4096385. PMID 25184667.
- ^ Крупович, Март; Кун, Йенс; Фишер, Меттиас (осень 2015 г.). «Система классификации вирофагов и спутниковых вирусов» (PDF). Архив вирусологии. 161 (1): 233–247. Дои:10.1007 / s00705-015-2622-9. PMID 26446887. S2CID 14196910 - через Springer.
- ^ Ру, Саймон; Чан, Леонг-Кит; Иган, Роб; Malmstrom, Rex R .; McMahon, Katherine D .; Салливан, Мэтью Б. (2017). «Экогеномика вирофагов и их гигантских вирусных хозяев, оцененная с помощью метагеномики временных рядов». Nature Communications. 8 (1): 858. Bibcode:2017НатКо ... 8..858R. Дои:10.1038 / s41467-017-01086-2. ISSN 2041-1723. ЧВК 5636890. PMID 29021524.
- ^ Паэс-Эспино, Дэвид; Чжоу, Цзинли; Ру, Саймон; Найфах, Стивен; Павлопулос, Георгиос А .; Шульц, Фредерик; McMahon, Katherine D .; Уолш, Дэвид; Войке, Таня; Иванова Наталья Н .; Элоэ-Фадрош, Эмили А .; Триндж, Сюзанна Дж .; Кирпидес, Никос К. (10 декабря 2019 г.). «Разнообразие, эволюция и классификация вирофагов, обнаруженных с помощью глобальной метагеномики». Микробиом. 7 (1): 157. Дои:10.1186 / s40168-019-0768-5. ЧВК 6905037. PMID 31823797.
- ^ Duponchel, S; Фишер, MG (2019). «Да здравствуют лавидавирусы! Пять особенностей вирофагов, паразитирующих на гигантских ДНК-вирусах». PLOS Pathog. 15 (3): e1007592. Дои:10.1371 / journal.ppat.1007592. ЧВК 6428243. PMID 30897185.. Материал был скопирован из этого источника, который доступен под Международная лицензия Creative Commons Attribution 4.0.
- ^ Фишер, М. Г .; Allen, M. J .; Wilson, W. H .; Саттл, К. А. (2010). «Гигантский вирус с замечательным набором генов заражает морской зоопланктон» (PDF). Труды Национальной академии наук. 107 (45): 19508–19513. Bibcode:2010PNAS..10719508F. Дои:10.1073 / pnas.1007615107. ЧВК 2984142. PMID 20974979.
- ^ Маттиас Г. Фишер; Майкл Дж. Аллен; Уильям Х. Уилсон; Кертис А. Саттл (2010). «Гигантский вирус с замечательным набором генов заражает морской зоопланктон» (PDF). Труды Национальной академии наук. 107 (45): 19508–19513. Bibcode:2010PNAS..10719508F. Дои:10.1073 / pnas.1007615107. ЧВК 2984142. PMID 20974979.
- ^ Массана, Рамон; Хавьер Дель Кампо; Кристиан Динтер; Рубен Соммаруга (2007). «Гибель популяции морских гетеротрофных жгутиконосцев. Кафетерий roenbergensis вирусной инфекцией ». Экологическая микробиология. 9 (11): 2660–2669. Дои:10.1111 / j.1462-2920.2007.01378.x. PMID 17922751. S2CID 30191542.
- ^ Уолдор М.К., Фридман Д.И., Адхья С.Л., ред. (2005). Фаги: их роль в бактериальном патогенезе и биотехнологии. Вашингтон, округ Колумбия: ASM Press. стр.450. ISBN 978-1-55581-307-9.
- ^ Кезе, Лаура; Гойер, Яна К. (2018). «Ответ фитопланктона на изменение морского климата - введение». YOUMARES 8 - Океаны через границы: учимся друг у друга. С. 55–71. Дои:10.1007/978-3-319-93284-2_5. ISBN 978-3-319-93283-5.
- ^ Weitz JS, Вильгельм SW (2013). «Море вирусов». Ученый. 27 (7): 35–39.
- ^ Suttle CA (сентябрь 2005 г.). «Вирусы в море». Природа. 437 (7057): 356–61. Bibcode:2005Натура.437..356С. Дои:10.1038 / природа04160. PMID 16163346. S2CID 4370363.
- ^ Вильгельм С.В., Саттл Калифорния (1999). «Вирусы и круговорот питательных веществ в море: вирусы играют решающую роль в структуре и функциях водных пищевых сетей». Бионаука. 49 (10): 781–88. Дои:10.2307/1313569. JSTOR 1313569.
- ^ Suttle CA (октябрь 2007 г.). «Морские вирусы - основные игроки в глобальной экосистеме». Обзоры природы Микробиология. 5 (10): 801–12. Дои:10.1038 / nrmicro1750. PMID 17853907. S2CID 4658457.
- ^ Suttle CA (сентябрь 2005 г.). «Вирусы в море». Природа. 437 (7057): 356–61. Bibcode:2005Натура.437..356С. Дои:10.1038 / природа04160. PMID 16163346. S2CID 4370363.
- ^ Шелфорд Э.Дж., Саттл, Калифорния (2018). «Опосредованный вирусами перенос азота от гетеротрофных бактерий к фитопланктону». Биогеонауки. 15 (3): 809–15. Bibcode:2018BGeo ... 15..809S. Дои:10.5194 / bg-15-809-2018.
- ^ Weinbauer, Markus G .; и другие. (2007). «Синергетические и антагонистические эффекты вирусного лизиса и выпаса протистана на бактериальную биомассу, производство и разнообразие». Экологическая микробиология. 9 (3): 777–788. Дои:10.1111 / j.1462-2920.2006.01200.x. PMID 17298376.
- ^ Робинсон, Кэрол и Нагаппа Рамаях. «Скорость микробного гетеротрофного метаболизма ограничивает микробный углеродный насос». Американская ассоциация развития науки, 2011 г.
- ^ Heinrichs, M.E., Mori, C. и Dlugosch, L. (2020) «Сложные взаимодействия между водными организмами и их химической средой, выясненные с разных точек зрения». В: YOUMARES 9-Океаны: наши исследования, наше будущее , страницы 279–297. Springer. Дои:10.1007/978-3-030-20389-4_15.
- ^ Давила-Рамос С., Кастелан-Санчес Х.Г., Мартинес-Авила Л., Санчес-Карбенте, доктор медицины, Перальта Р., Эрнандес-Мендоса А и др. (2019). «Обзор вирусной метагеномики в экстремальных условиях». Границы микробиологии. 10: 2403. Дои:10.3389 / fmicb.2019.02403. ЧВК 6842933. PMID 31749771.
- ^ Фурман, Джед А. (1999). «Морские вирусы и их биогеохимические и экологические эффекты». Природа. 399 (6736): 541–548. Bibcode:1999Натура.399..541F. Дои:10.1038/21119. ISSN 0028-0836. PMID 10376593. S2CID 1260399.
- ^ Цай, Ань-И, Гво-Цзин Гун и Ю-Вэнь Хуан. «Важность вирусного шунта в круговороте азота в Synechococcus Spp. Рост в субтропических прибрежных водах западной части Тихого океана». Наземные, атмосферные и океанические науки25.6 (2014).
- ^ Вильгельм, Стивен У .; Саттл, Кертис А. (1999). «Вирусы и круговорот питательных веществ в море: вирусы играют решающую роль в структуре и функциях водных пищевых сетей». Бионаука. 49 (10): 781–788. Дои:10.2307/1313569. JSTOR 1313569.
- ^ Бар-он, YM; Филлипс, Р. Майло, Р. (2018). «Распределение биомассы на Земле». PNAS. 115 (25): 6506–6511. Дои:10.1073 / pnas.1711842115. ЧВК 6016768. PMID 29784790.
- ^ а б Саттл, Кертис А. (2005). «Вирусы в море». Природа. 437 (7057): 356–361. Bibcode:2005Натура.437..356С. Дои:10.1038 / природа04160. ISSN 0028-0836. PMID 16163346. S2CID 4370363.
- ^ www.cdc.gov. Вредоносное цветение водорослей: Red Tide: Home [Проверено 19 декабря 2014 г.].
- ^ а б c Саттл, Кертис А. (октябрь 2007 г.). «Морские вирусы - основные игроки в глобальной экосистеме». Обзоры природы Микробиология. 5 (10): 801–812. Дои:10.1038 / nrmicro1750. ISSN 1740-1526. PMID 17853907. S2CID 4658457.
- ^ Брейтбарт М., Томпсон Л., Саттл С., Салливан М. (2007-06-01). «Изучение огромного разнообразия морских вирусов» (PDF). Океанография. 20 (2): 135–139. Дои:10.5670 / oceanog.2007.58.
- ^ Гурвиц Б.Л., У'Рен Дж.М. (июнь 2016 г.). «Перепрограммирование метаболизма вирусов в морских экосистемах». Текущее мнение в микробиологии. 31: 161–168. Дои:10.1016 / j.mib.2016.04.002. PMID 27088500.
- ^ Hurwitz BL, Hallam SJ, Sullivan MB (ноябрь 2013 г.). «Метаболическое перепрограммирование вирусами в солнечном и темном океане». Геномная биология. 14 (11): R123. Дои:10.1186 / gb-2013-14-11-r123. ЧВК 4053976. PMID 24200126.
- ^ Анантараман К., Духайме МБ, Брейер Дж. А., Вендт К. А., Тонер Б. М., Дик Дж. Дж. (Май 2014 г.). «Гены окисления серы в различных глубоководных вирусах». Наука. 344 (6185): 757–60. Bibcode:2014Научный ... 344..757A. Дои:10.1126 / science.1252229. HDL:1912/6700. PMID 24789974. S2CID 692770.
- ^ Ру С., Хоули А. К., Торрес Белтран М., Скофилд М., Швентек П., Степанаускас Р. и др. (Август 2014 г.). «Экология и эволюция вирусов, заражающих некультивируемые бактерии SUP05, выявленные одноклеточной и метагеномикой». eLife. 3: e03125. Дои:10.7554 / elife.03125. ЧВК 4164917. PMID 25171894.
- ^ Фортер П., Филипп Х (1999). «Последний универсальный общий предок (LUCA), простой или сложный?». Биологический бюллетень. 196 (3): 373–5, обсуждение 375–7. Дои:10.2307/1542973. JSTOR 1542973. PMID 11536914.
- ^ Калли, A.I .; Lang, A.S .; Саттл, К.А. (2006). «Метагеномный анализ прибрежных РНК-вирусных сообществ». Наука. 312 (5781): 1795–1798. Bibcode:2006Научный ... 312.1795C. Дои:10.1126 / science.1127404. PMID 16794078. S2CID 20194876.
- ^ а б Ральфф, Янина (2019). "Вирионейстон: обзор вирусно-бактериальных ассоциаций на границах раздела воздух-вода". Вирусы. 11 (2): 191. Дои:10.3390 / v11020191. ЧВК 6410083. PMID 30813345.. Материал был скопирован из этого источника, который доступен под Международная лицензия Creative Commons Attribution 4.0.
- ^ а б Хатчинсон Г.Е. (1961). «Парадокс планктона». Американский натуралист. 95 (882): 137–145. Дои:10.1086/282171. JSTOR 2458386.
- ^ Дановаро Р., Делль'Анно А., Коринальдези С., Маганьини М., Благородный Р., Тамбурини С., Вайнбауэр М. (август 2008 г.). «Серьезное вирусное воздействие на функционирование донных глубоководных экосистем». Природа. 454 (7208): 1084–7. Bibcode:2008 Натур.454.1084D. Дои:10.1038 / природа07268. PMID 18756250. S2CID 4331430.
- ^ Андерсон, Рика Э .; Brazelton, Уильям Дж .; Баросс, Джон А. (2011). «Влияют ли вирусы на генетический ландшафт глубинной биосферы?». Границы микробиологии. 2: 219. Дои:10.3389 / fmicb.2011.00219. ISSN 1664-302X. ЧВК 3211056. PMID 22084639.
- ^ Андерсон, Рика Э .; Sogin, Mitchell L .; Баросс, Джон А. (2014-10-03). «Эволюционные стратегии вирусов, бактерий и архей в экосистемах гидротермальных жерл, выявленные с помощью метагеномики». PLOS ONE. 9 (10): e109696. Bibcode:2014PLoSO ... 9j9696A. Дои:10.1371 / journal.pone.0109696. ISSN 1932-6203. ЧВК 4184897. PMID 25279954.
- ^ Ортманн, Алиса С.; Саттл, Кертис А. (август 2005 г.). «Высокая численность вирусов в системе глубоководных гидротермальных источников указывает на микробную смертность, опосредованную вирусами». Deep Sea Research Part I: Oceanographic Research Papers. 52 (8): 1515–1527. Bibcode:2005DSRI ... 52.1515O. Дои:10.1016 / j.dsr.2005.04.002. ISSN 0967-0637.
- ^ Брейтбарт, Мья (2012-01-15). «Морские вирусы: правда или действие». Ежегодный обзор морской науки. 4 (1): 425–448. Bibcode:2012 ОРУЖИЕ .... 4..425Б. Дои:10.1146 / annurev-marine-120709-142805. ISSN 1941-1405. PMID 22457982.
- ^ Голденфельд, Найджел; Woese, Карл (январь 2007 г.). «Следующая революция биологии». Природа. 445 (7126): 369. arXiv:q-bio / 0702015. Bibcode:2007Натура.445..369Г. Дои:10.1038 / 445369a. ISSN 0028-0836. PMID 17251963. S2CID 10737747.
- ^ Leggett HC, Buckling A, Long GH, Boots M (октябрь 2013 г.). «Генерализм и эволюция вирулентности паразитов». Тенденции в экологии и эволюции. 28 (10): 592–6. Дои:10.1016 / j.tree.2013.07.002. PMID 23968968.
- ^ Флорес CO, Valverde S, Weitz JS (март 2013 г.). «Многоуровневая структура и географические факторы перекрестного заражения морских бактерий и фагов». Журнал ISME. 7 (3): 520–32. Дои:10.1038 / ismej.2012.135. ЧВК 3578562. PMID 23178671.
- ^ Крупович М., Бэмфорд Д.Х. (июль 2007 г.). «Предполагаемые профаги, связанные с литическим бесхвостым фагом морской дцДНК PM2, широко распространены в геномах водных бактерий». BMC Genomics. 8: 236. Дои:10.1186/1471-2164-8-236. ЧВК 1950889. PMID 17634101.
- ^ Сюэ Х, Сюй И, Буше И, Польз М.Ф. (январь 2012 г.). «Высокая частота появления нового нитчатого фага, VCY φ, в популяции холерного вибриона в окружающей среде». Прикладная и экологическая микробиология. 78 (1): 28–33. Дои:10.1128 / AEM.06297-11. ЧВК 3255608. PMID 22020507.
- ^ а б Ру С., Крупович М., Пуле А., Деброа Д., Эно Ф. (2012). «Эволюция и разнообразие вирусного семейства Microviridae посредством коллекции из 81 нового полного генома, собранных из виромных прочтений». PLOS ONE. 7 (7): e40418. Bibcode:2012PLoSO ... 740418R. Дои:10.1371 / journal.pone.0040418. ЧВК 3394797. PMID 22808158.
- ^ Гурвиц Б.Л., Салливан МБ (2013). «Виром Тихого океана (POV): набор метагеномных данных морских вирусов и связанные с ними кластеры белков для количественной вирусной экологии». PLOS ONE. 8 (2): e57355. Bibcode:2013PLoSO ... 857355H. Дои:10.1371 / journal.pone.0057355. ЧВК 3585363. PMID 23468974.
- ^ Ру С, Эно Ф, Гурвиц Б.Л., Салливан МБ (28 мая 2015 г.). «VirSorter: извлечение вирусного сигнала из данных микробного генома». PeerJ. 3: e985. Дои:10.7717 / peerj.985. ЧВК 4451026. PMID 26038737.
- ^ Рен Дж., Альгрен Н.А., Лу Й.Й., Фурман Дж. А., Сан Ф. (июль 2017 г.). «VirFinder: новый инструмент на основе k-mer для идентификации вирусных последовательностей на основе собранных метагеномных данных». Микробиом. 5 (1): 69. Дои:10.1186 / s40168-017-0283-5. ЧВК 5501583. PMID 28683828.
- ^ Паез-Эспино Д., Элоэ-Фадрош Е.А., Павлопулос Г.А., Томас А.Д., Хантеманн М., Михайлова Н. и др. (Август 2016 г.). «Открытие вирома Земли». Природа. 536 (7617): 425–30. Bibcode:2016Натура.536..425P. Дои:10.1038 / природа19094. PMID 27533034. S2CID 4466854.
- ^ Coutinho FH, Silveira CB, Gregoracci GB, Thompson CC, Edwards RA, Brussaard CP и др. (Июль 2017 г.). «Морские вирусы, обнаруженные с помощью метагеномики, проливают свет на вирусные стратегии в океанах». Nature Communications. 8: 15955. Bibcode:2017НатКо ... 815955C. Дои:10.1038 / ncomms15955. ЧВК 5504273. PMID 28677677.
| Группы |  | |
|---|---|---|
| Микробиология | ||
| Экология | ||
| Растения | ||
| морской | ||
| Связанные с людьми | ||
| Методы | ||
| Другой | ||
| ||
| Компоненты |  | |
|---|---|---|
| Жизненный цикл вирусов | ||
| Генетика | ||
| Хозяин | ||
| Другой | ||
| ||