Морские грибы - Marine fungi
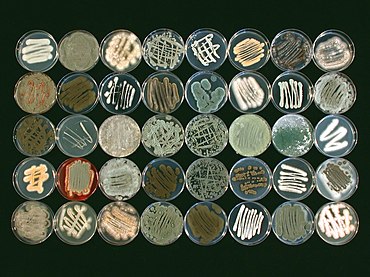
Морские грибы находятся разновидность из грибы которые живут в морской или же эстуарий среды. Они не таксономический группы, но имеют общую среду обитания. Облигатные морские грибы растут исключительно в морская среда обитания при полном или эпизодическом погружении в морскую воду. Факультативные морские грибы обычно обитают в наземных или пресноводных средах обитания, но способны жить или даже жить. спорулирующий в морской среде обитания. Описано около 444 видов морских грибов, в том числе семь родов и десять видов базидиомицеты, а также 177 родов и 360 видов аскомицеты. Остальные морские грибы хитриды и митоспорический или бесполые грибы.[2] Многие виды морских грибов известны только по спорам, и, вероятно, большое количество видов еще предстоит открыть.[3] Фактически, считается, что описано менее 1% всех видов морских грибов из-за трудностей с нацеливанием ДНК морских грибов и трудностей, возникающих при попытках выращивания культур морских грибов.[4] Невозможно культивировать многие из этих грибов, но их природу можно изучить, изучив образцы морской воды и рДНК анализ обнаруженного грибкового материала.[3]
Различные морские среды обитания поддерживают очень разные сообщества грибов. Грибы можно найти в нишах, начиная от океанских глубин и прибрежных вод до мангровые заросли болота и лиманы с низким уровнем солености.[5] Морские грибы могут быть сапробный или же паразитический на животных, сапробных или паразитарных на водоросли, сапробия для растений или сапробия для мертвой древесины.[2]
Обзор
Наземные грибы играют решающую роль в круговороте питательных веществ и пищевых сетях и могут формировать сообщества макроорганизмов как паразитов и мутуалистов. Хотя оценки количества видов грибов на планете колеблются от 1,5 до более 5 миллионов, вероятно, до сих пор идентифицировано менее 10% грибов. На сегодняшний день относительно небольшой процент описанных видов связан с морской средой, при этом около 1100 видов извлечены исключительно из морской среды. Тем не менее, грибы были обнаружены почти во всех исследованных морских средах обитания, от поверхности океана до километров под океанскими отложениями. Предполагается, что грибы вносят вклад в популяционные циклы фитопланктона и биологический углеродный насос, а также активны в химическом составе морских отложений. Многие грибы были идентифицированы как комменсалы или патогены морских животных (например, кораллов и губок), растений и водорослей. Несмотря на их различные роли, о разнообразии этой основной ветви эукариотической жизни в морских экосистемах или их экологических функциях известно очень мало.[6]
Грибы представляют собой большую и разнообразную группу микроорганизмов в микробиологических сообществах морской среды и играют важную роль в круговороте питательных веществ.[7] Они делятся на две основные группы; обязать морские грибы и факультативный морские грибы.[8] Облигатные морские грибы приспособлены к размножению в водной среде, в то время как факультативные морские грибы могут расти как в водной, так и в наземной среде.[8] Морские грибы называются грибами морского происхождения, если их факультативное или облигатное состояние не определено.[9]
Морские виды грибов встречаются как сапробы, паразиты, или же симбионты и колонизировать широкий спектр субстратов, таких как губки, кораллы, мангровые заросли, морские травы и водоросли.[10][11][9]
Факторы, которые влияют на то, присутствуют ли морские грибы в каком-либо конкретном месте, включают температуру воды, ее соленость, движение воды, наличие подходящих субстраты для колонизации - наличие пропагул в воде, межвидовая конкуренция, загрязнение и содержание кислорода в воде.[5]
Некоторые морские грибы, попавшие в море из наземных местообитаний, включают виды, которые зарываются в песчинки и живут в порах. Другие живут внутри каменистых кораллов и могут стать патогенными, если коралл подвергнется стрессу из-за повышения температуры моря.[3][12]
В 2011 г. филогения морских грибов выяснено путем анализа их малая единица рибосомная ДНК последовательности. Было обнаружено тридцать шесть новых морских линий, большинство из которых были хитриды но также некоторые нитчатые и многоклеточные грибы. Большинство найденных видов были аскомицетичный и базидиомицет дрожжи.[13]
Вторичные метаболиты, продуцируемые морскими грибами, имеют высокий потенциал для использования в биотехнологический, медицинские и промышленные применения.[14]
Эволюция
В отличие от растения и животные, ранняя летопись окаменелостей грибов скудна. Поскольку грибы не биоминерализовать, они не сразу попадают в летопись окаменелостей. Окаменелости грибов трудно отличить от окаменелостей других микробов, и их легче всего идентифицировать, если они напоминают сохранившийся грибы.[15]
Самые ранние окаменелости, обладающие чертами, характерными для грибов, относятся к Палеопротерозой эра, некоторые 2,400 миллион лет назад (Ма ). Эти многоклеточные бентосный у организмов были нитчатые структуры, способные к анастомоз, в котором рекомбинируются ветви гиф.[16] Другие недавние исследования (2009 г.) оценивают прибытие грибных организмов примерно через 760–1060 млн лет назад на основе сравнений скорости эволюции в близкородственных группах.[17]

Для большей части Палеозой Эра (542–251 млн лет назад), грибы, по всей видимости, были водными и состояли из организмов, похожих на современные Читриды наличие жгутика споры.[18] Филогенетический Анализы показывают, что жгутик был утерян на ранней стадии эволюции грибов, и, следовательно, у большинства видов грибов жгутик отсутствует.[19] Данные анализа ДНК показывают, что все грибы произошли от одного общего предка, по крайней мере, 600 миллионов лет назад. Вероятно, что эти самые ранние грибы жили в воде и имели жгутики. Грибы переселились на сушу примерно в то же время, что и растения, по крайней мере, около 460 миллионов лет назад.[20] Хотя грибы опистоконцы - группа эволюционно связанных организмов, в целом характеризующихся одним задним жгутик - все типы, кроме хитриды потеряли задние жгутики.[21]
Эволюционная адаптация от водного к наземному образу жизни потребовала диверсификации экологических стратегий получения питательных веществ, в том числе паразитизм, сапробизм, и развитие мутуалистический такие отношения как микориза и лихенизация.[22] Недавние (2009 г.) исследования показывают, что экологическое состояние предков Аскомикота был сапробизм, и это независимое лихенизация события произошли несколько раз.[23]
Рост грибов как гифы на твердых субстратах или в виде отдельных клеток в водной среде адаптирован для эффективного извлечения питательных веществ, поскольку эти формы роста имеют высокий отношение площади поверхности к объему.[24] Гифы специально приспособлены для роста на твердых поверхностях и вторжения субстраты и ткани.[25] Они могут проявлять большие проникающие механические силы; например, многие патогены растений, включая Magnaporthe grisea, образуют структуру, называемую аппрессориум которые эволюционировали, чтобы прокалывать ткани растений.[26] Давление, создаваемое аппрессорием, направленное против растения. эпидермис, может превышать 8 мегапаскали (1200 фунтов на квадратный дюйм).[26] Нитчатый гриб Paecilomyces lilacinus использует аналогичную структуру для проникновения в яйца нематоды.[27]
Грибы считались частью царство растений до середины 20 века. К середине 20 века грибы считались отдельным королевством, и недавно признанное королевство Грибы становится третьим крупным царством многоклеточных эукариоты с королевством Plantae и королевство Animalia, отличительной чертой этих царств является способ их получения.[28]
Морские растения
Мангровые заросли

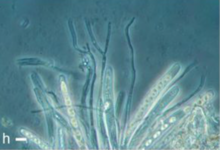
Наибольшее количество известных видов морских грибов происходит от мангровые заросли болота.[2] В одном исследовании блоки мангрового дерева и куски коряги Avicennia alba, Bruguiera cylindrica и Rhizophora apiculata были исследованы для выявления одревесневший (разлагающие дерево) грибы, которые они принимали. Также протестированы были Nypa fruticans, мангровая пальма и Acanthus ilicifolius, растение часто ассоциируется с мангровыми зарослями. Было обнаружено, что каждый материал имеет свои собственные характерные грибы, наибольшее разнообразие которых наблюдается на мангровых пальмах. Предполагалось, что это произошло потому, что соленость была ниже в эстуариях и ручьях, где Нипа росли, и поэтому грибам требовалась меньшая степень адаптации, чтобы там процветать. Некоторые из этих видов были тесно связаны с грибами наземных пальм. Другие исследования показали, что коряги содержат больше видов грибов, чем открытые образцы древесины аналогичного вида. Опад из листьев мангрового дерева также поддерживал большое грибковое сообщество, которое отличалось от такового на древесине и живом материале. Однако немногие из них были многоклеточными высшими морскими грибами.[5]
Другие растения


В морская улитка Littoraria irrorata повреждает растения Спартина в прибрежных морских болотах, где он обитает, что позволяет спорам приливных аскомицетных грибов колонизировать растение. Улитка ест грибок, а не траву. Этот мутуализм между улиткой и грибком считается первым примером содержания беспозвоночных животных вне класса Насекомое.[30]
Eelgrass, Зостера Марина, иногда влияет водоросли истощающая болезнь. Основной причиной этого, по-видимому, являются патогенные штаммы протист Лабиринтула зостерае, но считается, что грибковые патогены также вносят свой вклад и могут предрасполагать к болезни.[31][32]
Дерево

Многие морские грибы очень специфичны в отношении того, какие виды плавающей и подводной древесины они колонизируют. Населяют целый ряд видов грибов бук пока дуб поддерживает другое сообщество. Когда ростки гриба попадают на подходящий кусок дерева, они будут расти, если нет других грибов. Если древесина уже заселена другим видом грибов, рост будет зависеть от того, производит ли этот гриб противогрибковые химические вещества и сможет ли новичок противостоять им. Химические свойства колонизирующих грибов также влияют на сообщества животных, которые на них пасутся: в одном исследовании, когда гифы пяти разных видов морских грибов скармливались нематоды, один вид содержал менее половины количества нематод на мг гиф, чем другие.[33]
Обнаружение грибов в древесине может включать инкубацию при подходящей температуре в подходящей водной среде в течение периода от шести месяцев до восемнадцати месяцев.[33]
Лишайники

Лишайники находятся мутуалистический ассоциации между грибы, обычно аскомицет с базидиомицетом,[34] и водоросль или цианобактерии. Несколько лишайников, в том числе Arthopyrenia halodytes, Pharcidia ламинариикола, Pharcidia rhachiana и Turgidosculum ulvae, встречаются в морской среде.[2] Многие другие происходят в зона брызг, где они занимают разные вертикальные зоны в зависимости от их устойчивости к погружению.[35] Лишайник -подобные окаменелости были обнаружены в Формация Доушантуо в Китае около 600 миллионов лет назад.[36]
Грибы из Verrucariales также образуют морские лишайники с бурые водоросли Petroderma maculiforme,[37] и иметь симбиотические отношения с водоросли подобно (камыш ) и Blidingia minima, где водоросли являются доминирующими компонентами. Считается, что грибы помогают водорослям сопротивляться высыханию на воздухе.[38][39] Кроме того, лишайники также могут использовать желто-зеленые водоросли (Гетерококк ) как их симбиотический партнер.[40]
Лишеподобные окаменелости, состоящие из кокковидных клеток (цианобактерии ?) и тонкие нити (мукоромикотинан Гломеромикота ?) перминерализованы в морских фосфорит из Формация Доушантуо на юге Китая. Считается, что этим окаменелостям от 551 до 635 миллионов лет или Эдиакарский.[41] Эдиакарский акритархи также имеют много общего с Гломеромикотан везикулы и споры.[42] Также утверждалось, что Эдиакарские окаменелости включая Дикинсония,[43] были лишайники,[44] хотя это утверждение является спорным.[45] Эндосимбиотический Гломеромикота сравнимо с жизнью Геосифон может вернуться в Протерозойский в форме 1500 миллионов лет Городиския[46] и возраст 2200 миллионов лет Дискагма.[47] Обнаружение этих окаменелостей предполагает, что морские грибы развили симбиотические партнерские отношения с фотоавтотрофами задолго до эволюции сосудистых растений.
Водоросли и фитопланктон
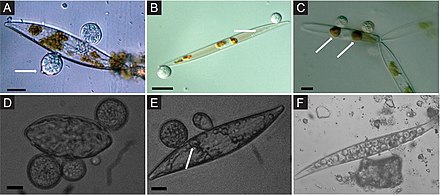
Морские грибы, связанные с водорослями, в значительной степени не исследованы, несмотря на их экологическую роль и потенциальное промышленное применение. Например, было показано, что грибы, связанные с водорослями, производят много биоактивных веществ. вторичные метаболиты.[49][50][51] Грибы, полученные из водорослей, могут быть связаны с различными водорослями, включая коричневый (например., Agarum clathratum, Фукус sp., Ламинария sp., Саргассум sp.), зеленый (например., Ульва sp., Энтероморфа sp., Флабеллия sp.), или красный (например. Хондрус sp., Дилси sp., Керамиум sp.) водоросли.[52][53][54][55][56][9]
Почти треть всех известных видов морских грибов связана с водорослями.[57] Наиболее часто описываемые грибы, связанные с водорослями, относятся к Аскомикота и представлены широким разнообразием родов, таких как Акремониум, Альтернариоз, Аспергиллы, Кладоспорий, Phoma, Пенициллий, Триходермия, Эмерицеллопсис, Ретрозий, Спатулоспора, Pontogenia и Сигмоидея.[58][59][60][61][62][63][64][9]
Rhyzophydium littoreum морпех Читрид, примитивный гриб, поражающий зеленые водоросли в эстуариях. Он получает питательные вещества из водорослей-хозяев и способствует плаванию. зооспоры которые должны выжить в открытой воде, в среде с низким содержанием питательных веществ, пока не встретится новый хозяин.[33] Другой грибок, Ascochyta salicorniae, найдено растущим на водоросли расследуется за его действия против малярия,[65] а инфекционное заболевание, передаваемое комарами людей и других животных.
Беспозвоночные
В Американский лобстер (Homarus americanus), как и многие другие морские ракообразные, насиживает яйца под сегментами хвоста. Здесь они подвергаются воздействию переносимых водой микроорганизмов, в том числе грибов, в течение длительного периода своего развития. У омара есть симбиотический отношения с грамотрицательная бактерия обладающий противогрибковыми свойствами. Эта бактерия растет над яйцами и защищает их от заражения патогенными грибами. оомицет Lagenidium callinectes. Метаболит, продуцируемый бактерией, является тирозол, 4-гидроксифенэтиловый спирт, антибиотик вещество также вырабатывается некоторыми наземными грибами. Аналогично креветка в устьях рек, Палемон макродактилис, имеет симбиотическую бактерию, которая производит 2,3-индолендион, вещество, которое также токсично для оомицетов. Lagenidium callinectes.[66]
Позвоночные

Киты, морские свиньи и дельфины восприимчивы к грибковым заболеваниям, но они мало исследованы в этой области. Сообщается о смертности от грибковых заболеваний среди косаток, содержащихся в неволе; считается, что предрасполагающим фактором мог быть стресс из-за условий содержания в неволе. Передача инфекции среди животных в открытом море может естественным образом ограничить распространение грибковых заболеваний. Инфекционные грибы, известные от косаток, включают: Aspergillus fumigatus, грибковые микроорганизмы албиканс и Saksenaea vasiformis. Грибковые инфекции у других китообразные включают Coccidioides immitis, Криптококк neoformans, Лобоа лобой, Ризопус sp., Aspergillus flavus, Blastomyces dermatitidus, Кладофиалофора бантиана, Histoplasma capsulatum, Мукор sp., Sporothrix schenckii и Трихофитон sp.[67]
Лососевые выращиваемые в садках в морской среде, могут быть поражены рядом различных грибковых инфекций. Экзофиала лососевая вызывает инфекцию, при которой рост гиф в почках вызывает вздутие живота. Клеточная реакция рыбы направлена на изоляцию грибка, отгораживая его. Рыбы также подвержены грибковым заболеваниям. оомицеты включая Бранхиомицеты который поражает жабры различных рыб, и Сапролегния который атакует поврежденные ткани.[68]
Морские отложения
Ascomycota, Basidiomycota и Chytridiomycota наблюдались в отложениях на глубине от 0 до 1740 метров под дном океана. В одном исследовании были проанализированы пробы морских отложений между этими глубинами и выделены все наблюдаемые грибы. Изоляты показали, что наибольшее разнообразие подповерхностных грибов было обнаружено на глубине от 0 до 25 метров ниже морского дна. Fusarium oxysporum и Rhodotorula mucilaginosa быть самым известным. В целом, аскомикоты являются доминирующим подповерхностным типом.[69] Почти все восстановленные виды грибов также наблюдались в наземных отложениях, причем источники спор указывают на их наземное происхождение.[69][70]
Вопреки предыдущим представлениям, глубинные морские грибы активно растут и прорастают, и некоторые исследования показывают повышенную скорость роста при высоком гидростатическом давлении. Хотя методы, с помощью которых морские грибы могут выжить в экстремальных условиях морского дна и ниже, в значительной степени неизвестны, Saccharomyces cerevisiae проливает свет на адаптации, которые делают это возможным. Этот гриб укрепляет свою внешнюю мембрану, чтобы выдерживать более высокое гидростатическое давление.[69]
Некоторые морские грибы, обитающие в донных отложениях, участвуют в биогеохимических процессах. Fusarium oxysporum и Fusarium solani являются денитрификаторами как в морской, так и в наземной среде.[69][71] Некоторые из них совместно денитрифицируют, превращая азот в закись азота и диазот.[70] Третьи перерабатывают органические вещества, включая углеводы, белки и липиды. Грибы океанической корки, подобные тем, которые встречаются вокруг гидротермальных источников, разлагают органическое вещество и играют различные роли в круговороте марганца и мышьяка.[6]
Морские грибы, связанные с отложениями, сыграли важную роль в разложении нефти, разлитой из Катастрофа Deepwater Horizons в 2010. Аспергиллы, Пенициллий, и Фузариум виды, среди прочего, могут разлагать высокомолекулярные углеводороды, а также помогать бактериям, разлагающим углеводороды.[6]
Арктические морские грибы

Морские грибы наблюдались даже на севере, до Северного Ледовитого океана. Chytridiomycota, доминирующий паразитический грибной организм в арктических водах, пользуется преимуществами цветения фитопланктона в солевых каналах, вызванного повышением температуры и повышенным проникновением света через лед. Эти грибы паразитируют на диатомовых водорослях, тем самым контролируя цветение водорослей и возвращая углерод обратно в пищевую сеть микробов. Цветение в Арктике также создает благоприятную среду для других паразитических грибов. Уровень освещенности и сезонные факторы, такие как температура и соленость, также контролируют активность хитридов независимо от популяций фитопланктона. В периоды низких температур и уровней фитопланктона, Ауреобазидиум и Кладоспорий популяции догоняют популяцию хитридов в каналах рассола.[73]
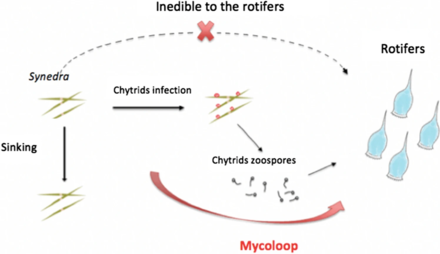
Пищевые сети и миколуп



Человеческое использование
Переработчики биомассы
Медицинское
Морские грибы производят противовирусные и антибактериальные соединения в виде метаболитов, более 1000 из которых реализованы и могут использоваться в качестве противоопухолевых, антидиабетических и противовоспалительных средств.[78][79]
Противовирусные свойства морских грибов были реализованы в 1988 году после того, как их соединения были успешно использованы для лечения вируса гриппа H1N1. Было показано, что помимо H1N1 противовирусные соединения, выделенные из морских грибов, оказывают вирулицидное действие на ВИЧ, простой герпес 1 и 2, Вирус репродуктивного и респираторного синдрома свиней, и Респираторно-синцитиальный вирус. Большинство этих противовирусных метаболитов были выделены из видов Аспергиллы, Пенициллий, Кладоспорий, Stachybotrys, и Неосартория. Эти метаболиты подавляют способность вируса к репликации, тем самым замедляя распространение инфекции.[78]
Грибы, ассоциированные с мангровыми деревьями, оказывают заметное антибактериальное действие на несколько распространенных патогенных бактерий человека, включая: Золотистый стафилококк и Синегнойная палочка. Высокая конкуренция между организмами в мангровых нишах приводит к увеличению антибактериальных веществ, вырабатываемых этими грибами в качестве защитных агентов.[80] Пенициллий и Аспергиллы виды являются крупнейшими продуцентами антибактериальных соединений среди морских грибов.[81]
Недавно было показано, что различные виды глубоководных морских грибов производят противораковые метаболиты. В одном исследовании было обнаружено 199 новых цитотоксических соединений с противораковым потенциалом. Помимо цитотоксических метаболитов, эти соединения имеют структуры, способные нарушать активируемые раком теломеразы через связывание ДНК. Другие препятствуют топоизомераза фермент от продолжения помощи в восстановлении и репликации раковых клеток.[79]
Смотрите также
Рекомендации
- ^ Paz, Z .; Комонь-Желязовская, М .; Дружинина, И. С .; Aveskamp, M. M .; Шнайдерман, А .; Aluma, Y .; Carmeli, S .; Илан, М .; Ярден, О. (30 января 2010 г.). «Разнообразие и потенциальные противогрибковые свойства грибов, связанных со средиземноморской губкой». Грибковое разнообразие. 42 (1): 17–26. Дои:10.1007 / s13225-010-0020-х. S2CID 45289302.
- ^ а б c d Виды высших морских грибов В архиве 2013-04-22 в Wayback Machine Университет Миссисипи. Проверено 5 февраля 2012.
- ^ а б c «Классная новая газета: морские грибы». Преподавание биологии. Архивировано из оригинал 16 марта 2012 г.[самостоятельно опубликованный источник? ]
- ^ Гладфельтер, Эми S .; Джеймс, Тимоти Й .; Поправьте, Энтони С. (март 2019 г.). «Морские грибы». Текущая биология. 29 (6): R191 – R195. Дои:10.1016 / j.cub.2019.02.009. PMID 30889385.
- ^ а б c Э. Б. Гарет Джонс (2000). «Морские грибы: некоторые факторы, влияющие на биоразнообразие» (PDF). Грибковое разнообразие. 4: 53–73.
- ^ а б c d Amend, A., Burgaud, G., Cunliffe, M., Edgcomb, VP, Ettinger, CL, Gutiérrez, MH, Heitman, J., Hom, EF, Ianiri, G., Jones, AC, и Kagami, M. ( 2019) «Грибы в морской среде: открытые вопросы и нерешенные проблемы». MBio, 10(2): e01189-18. Дои:10,1128 / мБио.01189-18.
 Материал был скопирован из этого источника, который доступен под Международная лицензия Creative Commons Attribution 4.0.
Материал был скопирован из этого источника, который доступен под Международная лицензия Creative Commons Attribution 4.0. - ^ Tisthammer, K.H .; Cobian, G.M .; Поправьте, A.S. Глобальная биогеография морских грибов определяется окружающей средой. Fungal Ecol. 2016, 19, 39–46.
- ^ а б Рагхукумар, С. Морская среда и роль грибов. В грибах в прибрежных и океанических морских экосистемах: морские грибы; Springer International Publishing: Cham, Швейцария, 2017; С. 17–38.
- ^ а б c d Патышакулиева А., Фалькоски Д.Л., Вибенга А., Тиммерманс К. и Де Вриз Р.П. (2020) «Грибы, полученные из макроводорослей, обладают высокой способностью разрушать полимеры водорослей». Микроорганизмы, 8(1): 52. Дои:10.3390 / микроорганизмы8010052. Материал был скопирован из этого источника, который доступен под Международная лицензия Creative Commons Attribution 4.0.
- ^ Jones, E.B.G .; Suetrong, S .; Sakayaroj, J .; Bahkali, A.H .; Abdel-Wahab, M.A .; Боеут, Т .; Панг, К.-Л. Классификация морских Ascomycota, Basidiomycota, Blastocladiomycota и Chytridiomycota. Грибные ныряльщики. 2015, 73, 1–72.
- ^ Debbab, A .; Aly, A.H .; Прокш, П. Мангровые грибковые эндофиты - химическое и биологическое восприятие. Грибные ныряльщики. 2013, 61, 1–27.
- ^ Holmquist, G.U .; Х. В. Уокер и Стар Х. М. (1983). «Влияние температуры, pH, активности воды и противогрибковых агентов на рост Aspergillus flavus и A. parasiticus". Журнал пищевой науки. 48 (3): 778–782. Дои:10.1111 / j.1365-2621.1983.tb14897.x.
- ^ Richards, Thomas A .; Джонс, Мередит Д.М.; Леонард, Гай; Басс, Дэвид (15 января 2012 г.). «Морские грибы: их экология и молекулярное разнообразие». Ежегодный обзор морской науки. 4 (1): 495–522. Bibcode:2012РУЖИЕ .... 4..495р. Дои:10.1146 / аннурьев-марин-120710-100802. PMID 22457985.
- ^ Морские грибы Проверено 6 февраля 2012.
- ^ Донохью MJ; Крафт J (2004). Сборка Древа Жизни. Оксфорд (Оксфордшир), Великобритания: Oxford University Press. п. 187. ISBN 978-0-19-517234-8.
- ^ Бенгтсон, Стефан; Расмуссен, Биргер; Иварссон, Магнус; Мюлинг, Джанет; Броман, Курт; Мароне, Федерика; Стампанони, Марко; Беккер, Андрей (24 апреля 2017 г.). «Грибоподобные окаменелости мицелия в везикулярном базальте возрастом 2,4 миллиарда лет». Природа Экология и эволюция. 1 (6): 0141. Дои:10.1038 / s41559-017-0141. ISSN 2397-334X. PMID 28812648. S2CID 25586788.
- ^ Lucking R, Huhndorf S, Pfister D, Plata ER, Lumbsch H (2009). «Грибки эволюционировали прямо на ходу». Микология. 101 (6): 810–822. Дои:10.3852/09-016. PMID 19927746. S2CID 6689439.
- ^ Джеймс Т.Ю .; и другие. (2006). «Реконструкция ранней эволюции грибов с использованием филогении с шестью генами». Природа. 443 (7113): 818–22. Bibcode:2006Натура.443..818J. Дои:10.1038 / природа05110. PMID 17051209. S2CID 4302864.
- ^ Лю Ю.Дж., Ходсон М.С., Холл Б.Д. (2006). «Потеря жгутика произошла только один раз в грибковой линии: филогенетическая структура Kingdom Fungi выведена из генов субъединиц РНК-полимеразы II». BMC Эволюционная биология. 6 (1): 74. Дои:10.1186/1471-2148-6-74. ЧВК 1599754. PMID 17010206.
- ^ «СК12-Фундамент». flexbooks.ck12.org. Получено 2020-05-19.
- ^ Стинкамп Е.Т., Райт Дж., Балдауф С.Л. (январь 2006 г.). «Протестированное происхождение животных и грибов». Молекулярная биология и эволюция. 23 (1): 93–106. Дои:10.1093 / molbev / msj011. PMID 16151185.
- ^ Тейлор и Тейлор, стр. 84–94 и 106–107.
- ^ Schoch CL; Sung GH; Лопес-Хиральдес Ф; и другие. (2009). «Древо жизни Ascomycota: филогенетическая филогения проясняет происхождение и эволюцию основных репродуктивных и экологических черт». Систематическая биология. 58 (2): 224–39. Дои:10.1093 / sysbio / syp020. PMID 20525580.
- ^ Мосс СТ (1986). Биология морских грибов. Кембридж, Великобритания: Издательство Кембриджского университета. п. 76. ISBN 978-0-521-30899-1.
- ^ Пеньяльва М.А., Арст Х.Н. (сентябрь 2002 г.). «Регулирование экспрессии генов с помощью pH окружающей среды в мицелиальных грибах и дрожжах». Обзоры микробиологии и молекулярной биологии. 66 (3): 426–46, содержание. Дои:10.1128 / MMBR.66.3.426-446.2002. ЧВК 120796. PMID 12208998.
- ^ а б Говард Р.Дж., Феррари М.А., Роуч Д.Х., Деньги Н.П. (декабрь 1991 г.). «Проникновение грибка в твердые субстраты с огромным тургорным давлением». Труды Национальной академии наук Соединенных Штатов Америки. 88 (24): 11281–4. Bibcode:1991PNAS ... 8811281H. Дои:10.1073 / пнас.88.24.11281. ЧВК 53118. PMID 1837147.
- ^ Деньги НП (1998). «Механика инвазивного роста грибов и значение тургора при заражении растений». Молекулярная генетика специфических для хозяев токсинов при болезнях растений: материалы 3-го Международного симпозиума Тоттори по специфическим для хозяина токсинам, Дайсен, Тоттори, Япония, 24–29 августа 1997 г.. Нидерланды: Kluwer Academic Publishers. С. 261–271. ISBN 978-0-7923-4981-5.
- ^ «Грибок - Эволюция и филогения грибов». Энциклопедия Британника. Получено 2020-05-19.
- ^ Фраяр, С.С., Хэлеуотерс, Д., Кэтчесайд, Д. (2019) "Аннабелла австралийская ген. & sp. nov. (Helotiales, Cordieritidaceae) из мангровых зарослей Южной Австралии ". Микологический прогресс, 18(7): 973–981. Дои:10.1007 / s11557-019-01499-x.
- ^ Силлиман Б. Р. и С. Ю. Ньюэлл (2003). «Грибное хозяйство в улитке». PNAS. 100 (26): 15643–15648. Bibcode:2003ПНАС..10015643С. Дои:10.1073 / pnas.2535227100. ЧВК 307621. PMID 14657360.
- ^ Анализ болезней архипелага Сан-Хуан В архиве 2013-09-12 в Wayback Machine Лаборатория морских водорослей Friday Harbour Laboratories. Проверено 6 февраля 2012.
- ^ Коротко, Фредерик Т .; Роберт Г. Коулз (2001-11-06). Глобальные методы исследования водорослей. п. 414. ISBN 9780080525617.
- ^ а б c Мосс, Стивен Т. (1986). Биология морских грибов. С. 65–70. ISBN 9780521308991.
- ^ Сприбиль, Тоби; Туовинен, Веера; Resl, Philipp; Вандерпул, Дэн; Волински, Хеймо; Эйме, М. Кэтрин; Шнайдер, Кевин; Stabentheiner, Эдит; Тооме-Хеллер, Мерже; Тор, Горан; Майрхофер, Гельмут; Йоханнессон, Ханна; Маккатчеон, Джон П. (29 июля 2016 г.). «Базидиомицетные дрожжи в коре аскомицетных макролишайников». Наука. 353 (6298): 488–492. Bibcode:2016Научный ... 353..488С. Дои:10.1126 / science.aaf8287. ЧВК 5793994. PMID 27445309.
- ^ Пресноводные и морские лишайниковые грибы Проверено 6 февраля 2012.
- ^ Юань X, Сяо С., Тейлор Т.Н.; Сяо; Тейлор (2005). «Симбиоз типа лишайников 600 миллионов лет назад». Наука. 308 (5724): 1017–20. Bibcode:2005Наука ... 308.1017Y. Дои:10.1126 / science.1111347. PMID 15890881. S2CID 27083645.CS1 maint: несколько имен: список авторов (связь)
- ^ Sanders, W. B .; Moe, R. L .; Аскасо, К. (2004). «Приливной морской лишайник, образованный пиреномицетным грибом Verrucaria tavaresiae (Ascomycotina) и бурой водорослью Petroderma maculiforme (Phaeophyceae): организация таллома и взаимодействие симбионтов - NCBI». Американский журнал ботаники. 91 (4): 511–22. Дои:10.3732 / ajb.91.4.511. PMID 21653406.
- ^ «Взаимоотношения грибов и водорослей - Музей Нью-Брансуика». В архиве из оригинала 18 сентября 2018 г.. Получено 4 октября 2018.
- ^ Миллер, Кэти Энн; Перес-Ортега, Серхио. «Оспаривание концепции лишайников: Turgidosculum ulvae - Кембридж». Лихенолог. 50 (3): 341–356. Дои:10.1017 / S0024282918000117. В архиве из оригинала 7 октября 2018 г.. Получено 7 октября 2018.
- ^ Рыбалка, Н .; Wolf, M .; Андерсен, Р. А .; Фридл, Т. (2013). «Конгруэнтность хлоропластов - Эволюционная биология BMC - BioMed Central». BMC Эволюционная биология. 13: 39. Дои:10.1186/1471-2148-13-39. ЧВК 3598724. PMID 23402662.
- ^ Юань X, Сяо С., Тейлор Т. Н. (2005). «Симбиоз типа лишайников 600 миллионов лет назад». Наука. 308 (5724): 1017–1020. Bibcode:2005Наука ... 308.1017Y. Дои:10.1126 / science.1111347. PMID 15890881. S2CID 27083645.
- ^ Retallack G.J. (2015). «Акритарх свидетельствует о позднедокембрийской адаптивной радиации грибов» (PDF). Botanica Pacifica. 4 (2): 19–33. Дои:10.17581 / bp.2015.04203. В архиве (PDF) из оригинала 22 декабря 2016 г.. Получено 22 декабря 2016.
- ^ Retallack GJ. (2007). «Рост, распад и уплотнение погребения Дикинсонии, культовой ископаемой эдиакарской останки». Алчеринга: Австралазийский журнал палеонтологии. 31 (3): 215–240. Дои:10.1080/03115510701484705. S2CID 17181699.
- ^ Retallack GJ. (1994). «Были ли эдиакарские ископаемые лишайники?». Палеобиология. 20 (4): 523–544. Дои:10,1017 / с0094837300012975. JSTOR 2401233.
- ^ Свитек Б (2012). «Жизнь спорное утверждение ставит на земле 65 миллионов лет раньше». Природа. Дои:10.1038 / природа.2012.12017. S2CID 130305901. В архиве из оригинала на 1 января 2013 г.. Получено 2 января 2013.
- ^ Retallack, G.J .; Dunn, K.L .; Саксби, Дж. (2015). «Проблемная мезопротерозойская окаменелость Городиския из национального парка Глейшер, Монтана, США». Докембрийские исследования. 226: 125–142. Bibcode:2013PreR..226..125R. Дои:10.1016 / j.precamres.2012.12.005.
- ^ Retallack, G.J .; Krull, E.S .; Thackray, G.D .; Паркинсон, Д. (2013). «Проблемные окаменелости в форме урн из палеопротерозойских (2,2 млрд лет) палеопочв в Южной Африке». Докембрийские исследования. 235: 71–87. Bibcode:2013PreR..235 ... 71R. Дои:10.1016 / j.precamres.2013.05.015.
- ^ Hassett, BT; Грейдингер, Р. (2016). «Читриды доминируют в сообществах арктических морских грибов». Environ Microbiol. 18 (6): 2001–2009. Дои:10.1111/1462-2920.13216. PMID 26754171.
- ^ Overy, D.P .; Bayman, P .; Kerr, R.G .; Биллз, Г.Ф. Оценка обнаружения натуральных продуктов из морских (sensu strictu) и морских грибов. Микология 2014, 5, 145–167.
- ^ Flewelling, A.J .; Currie, J .; Gray, C.A .; Джонсон, Дж. Эндофиты морских макроводорослей: многообещающие источники новых натуральных продуктов. Curr. Sci. 2015, 109, 88–111.
- ^ Miao, F .; Zuo, J .; Лю, X .; Джи, Н. Альгицидная активность вторичных метаболитов эндофитных грибов, происходящих из морских макроводорослей. J. Oceanol. Лимнол. 2019, 37, 112–121.
- ^ Gnavi, G .; Garzoli, L .; Poli, A .; Prigione, V .; Burgaud, G .; Варезе, Г. Культурная микобиота Flabellia petiolata: первое исследование морских грибов, связанных со средиземноморскими зелеными водорослями. PLoS ONE 2017, 12, e0175941.
- ^ Kohlmeyer, J .; Фолькманн-Кольмейер, Б. Иллюстрированный ключ к нитчатым высшим морским грибам. Бот. Мар.1991, 34, 1–61.
- ^ Стэнли, С.Дж. Наблюдения за сезонностью морских эндофитных и паразитических грибов. Может. J. Bot. 1992, 70, 2089–2096.
- ^ Zuccaro, A .; Schoch, C.L .; Spatafora, J.W .; Kohlmeyer, J .; Draeger, S .; Митчелл, Дж. Обнаружение и идентификация грибов, тесно связанных с бурыми водорослями Fucus serratus. Appl. Environ. Microbiol. 2008, 74, 931–941.
- ^ Lee, S .; Парк, M.S .; Lee, H .; Kim, J.J .; Eimes, J.A .; Лим, Ю.В. Разнообразие грибов и активность ферментов, связанных с макроводорослями Agarum clathratum. Микобиология 2019, 47, 50–58.
- ^ Балабанова, Л .; Слепченко, Л .; Сын, О .; Текутьева, Л. Биотехнологический потенциал морских грибов, разлагающих полимерные субстраты растений и водорослей. Передний. Microbiol. 2018, 9, 15–27.
- ^ Kohlmeyer, J .; Фолькманн-Кольмейер, Б. Иллюстрированный ключ к нитчатым высшим морским грибам. Бот. Мар.1991, 34, 1–61.
- ^ Стэнли, С.Дж. Наблюдения за сезонностью морских эндофитных и паразитических грибов. Может. J. Bot. 1992, 70, 2089–2096. [Google Scholar] [CrossRef]
- ^ Zuccaro, A .; Schoch, C.L .; Spatafora, J.W .; Kohlmeyer, J .; Draeger, S .; Митчелл, Дж. Обнаружение и идентификация грибов, тесно связанных с бурыми водорослями Fucus serratus. Appl. Environ. Microbiol. 2008, 74, 931–941.
- ^ Lee, S .; Парк, M.S .; Lee, H .; Kim, J.J .; Eimes, J.A .; Лим, Ю.В. Разнообразие грибов и активность ферментов, связанных с макроводорослями Agarum clathratum. Микобиология 2019, 47, 50–58.
- ^ Zuccaro, A .; Саммербелл, R.C .; Gams, W .; Schroers, H.-J .; Митчелл, Дж. Новый вид Acremonium, связанный с Fucus spp., И его родство с филогенетически отличной морской кладой Emericellopsis. Stud. Mycol. 2004, 50, 283–297.
- ^ Kohlmeyer, J .; Кольмейер, Э. Морская микология: высшие грибы; Эльзевир: Амстердам, Нидерланды, 2013.
- ^ Wainwright, B.J .; Bauman, A.G .; Zahn, G.L .; Todd, P.A .; Хуанг, Д. Характеристика грибного биоразнообразия и сообществ, связанных с рифовой макроводорослью Sargassum ilicifolium, выявляет дифференциацию грибного сообщества в соответствии с географическим местоположением и структурой водорослей. Март Biodivers. 2019, 49, 2601–2608.
- ^ Osterhage C .; Р. Каминский; Г. Кениг и А. Д. Райт (2000). "Аскосалипирролидинон А, противомикробный алкалоид, из облигатного морского грибка" Ascochyta salicorniae". Журнал органической химии. 65 (20): 6412–6417. Дои:10.1021 / jo000307g. PMID 11052082.
- ^ Гил-Тернес, М. София и Уильям Феникал (1992). "Эмбрионы Homarus americanus защищены эпибиотическими бактериями ». Биологический бюллетень. 182 (1): 105–108. Дои:10.2307/1542184. JSTOR 1542184. PMID 29304709.
- ^ Gaydos, Joseph K .; Balcomb, Kenneth C .; Осборн, Ричард В .; Dierauf, Лесли. Обзор потенциальных угроз инфекционных заболеваний для южных обитателей косаток (Orcinus orca) (PDF) (Отчет). Архивировано из оригинал (PDF) на 2016-03-04. Получено 2012-09-28.
- ^ Грибковые инфекции выращиваемого лосося и форели Проверено 6 февраля 2012.
- ^ а б c d Реду, Ванесса; Наварри, Марион; Месле-Кладьер, Лоуренс; Барбье, Жорж; Бурго, Гаэтан (15 мая 2015 г.). Брахейдж, А.А. (ред.). «Видовое богатство и адаптация морских грибов из глубоководных отложений». Прикладная и экологическая микробиология. 81 (10): 3571–3583. Дои:10.1128 / AEM.04064-14. ISSN 0099-2240. ЧВК 4407237. PMID 25769836.
- ^ а б Мутон, Марнел; Постма, Фердинанд; Wilsenach, Jac; Бота, Альфред (август 2012). «Разнообразие и характеристика культивируемых грибов из морских отложений, собранных в заливе Св. Елены, Южная Африка». Микробная экология. 64 (2): 311–319. Дои:10.1007 / s00248-012-0035-9. ISSN 0095-3628. PMID 22430506. S2CID 9724720.
- ^ Shoun, H .; Танимото, Т. (15.06.1991). «Денитрификация грибком Fusarium oxysporum и участие цитохрома Р-450 в восстановлении дыхательных нитритов». Журнал биологической химии. 266 (17): 11078–11082. ISSN 0021-9258. PMID 2040619.
- ^ Килиас, Эстель С .; Юнгес, Леандро; Шупраха, Лука; Леонард, Гай; Метфиес, Катя; Ричардс, Томас А. (2020). «Распространение хитридовых грибов и их совместное появление с диатомовыми водорослями коррелируют с таянием морского льда в Северном Ледовитом океане». Биология коммуникации. 3 (1): 183. Дои:10.1038 / с42003-020-0891-7. ЧВК 7174370. PMID 32317738. S2CID 216033140. Материал был скопирован из этого источника, который доступен под Международная лицензия Creative Commons Attribution 4.0.
- ^ Hassett, B.T .; Градингер, Р. (2016). «Читриды доминируют в сообществах арктических морских грибов». Экологическая микробиология. 18 (6): 2001–2009. Дои:10.1111/1462-2920.13216. ISSN 1462-2920. PMID 26754171.
- ^ Кагами М., Мики Т. и Такимото Г. (2014) «Mycoloop: хитриды в водных пищевых сетях». Границы микробиологии, 5: 166. Дои:10.3389 / fmicb.2014.00166. Материал был скопирован из этого источника, который доступен под Международная лицензия Creative Commons Attribution 3.0.
- ^ Gutierrez MH, Jara AM, Pantoja S (2016) «Грибковые паразиты заражают морских диатомовых водорослей в апвеллинговой экосистеме системы течения Гумбольдта в центральной части Чили». Environ Microbiol, 18(5): 1646–1653. Дои:10.1111/1462-2920.13257.
- ^ Френкен, Т., Алацид, Э., Бергер, С.А., Борн, Э.С., Герфагнон, М., Гроссарт, Х.П., Гселл, А.С., Ибелингс, Б.В., Кагами, М., Кюппер, Ф.С. и Летчер, П. (2017) «Интеграция хитридовых грибковых паразитов в экологию планктона: пробелы в исследованиях и потребности». Экологическая микробиология, 19(10): 3802–3822. Дои:10.1111/1462-2920.13827. Материал был скопирован из этого источника, который доступен под Международная лицензия Creative Commons Attribution 4.0.
- ^ Барранко, В.С., Ван дер Меер, М.Т., Кагами, М., Ван ден Вингаерт, С., Ван де Ваал, Д.Б., Ван Донк, Э. и Гселл, А.С. (2020 г. («Трофическая позиция, соотношение элементов и перенос азота в планктонной пищевой цепи хозяин – паразит – потребитель, включая грибкового паразита». Oecologia: 1–14. Дои:10.1007 / s00442-020-04721-w. Материал был скопирован из этого источника, который доступен под Международная лицензия Creative Commons Attribution 4.0.
- ^ а б Могхадамтуси, Сохейл; Никзад, Соня; Кадир, Хабса; Абубакар, Сазалы; Занди, Кейван (22 июля 2015 г.). «Потенциальные противовирусные средства от морских грибов: обзор». Морские препараты. 13 (7): 4520–4538. Дои:10.3390 / md13074520. ISSN 1660-3397. ЧВК 4515631. PMID 26204947.
- ^ а б Дешмук, Сунил К .; Пракаш, Вед; Ранджан, Нихар (2018-01-05). «Морские грибы: источник потенциальных противораковых соединений». Границы микробиологии. 8: 2536. Дои:10.3389 / fmicb.2017.02536. ISSN 1664-302X. ЧВК 5760561. PMID 29354097.
- ^ Чжоу, Сунлинь; Ван, Мин; Фэн, Ци; Линь, Иньин; Чжао, Хуангэ (декабрь 2016 г.). «Исследование биологической активности морских грибов из различных местообитаний в прибрежных регионах». SpringerPlus. 5 (1): 1966. Дои:10.1186 / с40064-016-3658-3. ISSN 2193-1801. ЧВК 5108748. PMID 27933244.
- ^ Сюй, Лицзянь; Мэн, Вэй; Цао, Конг; Ван, Цзянь; Шань, Вэньцзюнь; Ван, Цингуй (2015-06-02). «Антибактериальные и противогрибковые соединения морских грибов». Морские препараты. 13 (6): 3479–3513. Дои:10.3390 / md13063479. ISSN 1660-3397. ЧВК 4483641. PMID 26042616.
дальнейшее чтение
- Гарет Джонс, Э. Б. и Панг, Ка-Лай (2012) Морские грибы: и грибоподобные организмы Морская и пресноводная ботаника Вальтер де Грюйтер. ISBN 9783110264067.
- Рагхукумар, Чандралата (2012) Биология морских грибов Springer. ISBN 9783642233425.
- Рагхукумар, Сешагири (2017) Грибы в прибрежных и океанических морских экосистемах Springer. ISBN 9783319543048.