Бактериофаг - Bacteriophage



А бактериофаг (/бækˈтɪərяoʊжeɪdʒ/), также известный неофициально как фаг (/жeɪdʒ/), это вирус который заражает и размножается внутри бактерии и археи. Этот термин произошел от слова «бактерии» и Греческий φαγεῖν (фагеин), что означает «пожирать». Бактериофаги состоят из белки который инкапсулировать а ДНК или же РНК геном, и могут иметь простую или сложную структуру. Их геномы могут кодировать всего четыре гена (например, MS2 ) и целых сотни гены. Фаги размножаются внутри бактерии после инъекции их генома в ее цитоплазма.
Бактериофаги - одни из самых распространенных и разнообразных организмов в биосфера.[1] Бактериофаги - это повсеместно распространенные вирусы, которые встречаются везде, где существуют бактерии. По оценкам, их более 1031 бактериофагов на планете больше, чем у всех других организмов на Земле, включая бактерии, вместе взятые.[2] Одним из самых плотных естественных источников фагов и других вирусов является морская вода, где до 9х108 вирионы на миллилитр были обнаружены в микробные маты на поверхности,[3] и до 70% морские бактерии могут быть инфицированы фагами.[4]
Фаги используются с конца 20 века как альтернатива антибиотики в бывшем Советский союз и Центральной Европе, а также во Франции.[5][6] Они рассматриваются как возможное лечение против мультирезистентный штаммы многих бактерий (см. фаговая терапия ).[7] С другой стороны, фаги Inoviridae было показано, что усложняет биопленки участвует в пневмония и кистозный фиброз и для защиты бактерий от лекарств, предназначенных для искоренения болезней, тем самым способствуя развитию стойкой инфекции.[8]
Классификация
Бактериофаги широко распространены в биосфере, имеют разные геномы и образ жизни. Фаги классифицируются по Международный комитет по таксономии вирусов (ICTV) согласно морфология и нуклеиновая кислота.

| Заказ | Семья | Морфология | Нуклеиновая кислота | Примеры |
|---|---|---|---|---|
| Белфривиралес | Turriviridae | Обернутый, изометрический | Линейная дцДНК | |
| Caudovirales | Ackermannviridae | Неокутанный, сократительный хвост | Линейная дцДНК | |
| Myoviridae | Безобутальный сократительный хвост | Линейная дцДНК | Т4, Му, P1, P2 | |
| Siphoviridae | Хвост без оболочки, не сокращающийся (длинный) | Линейная дцДНК | λ, Т5, HK97, N15 | |
| Podoviridae | Хвост без оболочки, не сокращающийся (короткий) | Линейная дцДНК | T7, Т3, Φ29, P22 | |
| Halopanivirales | Sphaerolipoviridae | Обернутый, изометрический | Линейная дцДНК | |
| Haloruvirales | Pleolipoviridae | Обволакивающий, плеоморфный | Круговая оцДНК, круговая дцДНК или линейная дцДНК | |
| Kalamavirales | Tectiviridae | Без оболочки, изометрическая | Линейная дцДНК | |
| Левивиралес | Левивириды | Без оболочки, изометрическая | Линейная оцРНК | MS2, Qβ |
| Ligamenvirales | Lipothrixviridae | Обволакивающий, стержневидный | Линейная дцДНК | Нитчатый вирус Acidianus 1 |
| Rudiviridae | Без оболочки, стержневидный | Линейная дцДНК | Палочковидный вирус Sulfolobus islandicus 1 | |
| Миндивиралес | Cystoviridae | Обволакивающий, сферический | Сегментированная дцРНК | |
| Petitvirales | Микровирусы | Без оболочки, изометрическая | Круговая оцДНК | ΦX174 |
| Tubulavirales | Inoviridae | Без оболочки, нитчатый | Круговая оцДНК | M13 |
| Винавиралес | Corticoviridae | Без оболочки, изометрическая | Круговая дцДНК | PM2 |
| Не назначен | Ampullaviridae | Обернутый, в форме бутылки | Линейная дцДНК | |
| Bicaudaviridae | Без оболочки, в форме лимона | Круговая дцДНК | ||
| Clavaviridae | Без оболочки, стержневидный | Круговая дцДНК | ||
| Finnlakeviridae | дцДНК | Кувырок[9] | ||
| Fuselloviridae | Без оболочки, в форме лимона | Круговая дцДНК | ||
| Globuloviridae | Обернутый, изометрический | Линейная дцДНК | ||
| Guttaviridae | Без оболочки, яйцевидной формы | Круговая дцДНК | ||
| Plasmaviridae | Обволакивающий, плеоморфный | Круговая дцДНК | ||
| Portogloboviridae | Обернутый, изометрический | Круговая дцДНК | ||
| Спиравиры | Без развертки, стержневидный | Круговая оцДНК | ||
| Tristromaviridae | Обволакивающий, стержневидный | Линейная дцДНК |
Было высказано предположение, что члены Picobirnaviridae заражают бактерии, но не млекопитающих.[10]
Еще одна предлагаемая семья: "Autolykiviridae "(дцДНК).[11]
История
В 1896 г. Эрнест Хэнбери Ханкин сообщил, что что-то в водах Ганг и Ямуна реки в Индия был отмечен антибактериальный действие против холера и он мог проходить через очень тонкий фарфоровый фильтр.[12] В 1915 г. Британский бактериолог Фредерик Творт, суперинтендант Лондонского института Брауна, обнаружил небольшой агент, заражающий и убивающий бактерии. Он считал, что агент должен быть одним из следующих:
- этап в жизненный цикл бактерий
- ан фермент производятся самими бактериями, или
- вирус, который вырос и уничтожил бактерии[13]
Исследования Творта были прерваны началом Первая Мировая Война, а также дефицит финансирования и открытия антибиотиков.

Независимо, Французский канадец микробиолог Феликс д'Эрелль, работая в Институт Пастера в Париж, объявил 3 сентября 1917 г., что он открыл «невидимый антагонистический микроб дизентерия bacillus ». Для д'Эрелля не возникало никаких сомнений относительно природы его открытия:« В мгновение ока я понял: причиной моих чистых пятен был невидимый микроб… вирус, паразитирующий на бактериях ».[14] Д'Эрелль назвал вирус бактериофагом, поедающим бактерии (от греч. фагеин означает «пожирать»). Он также записал драматический рассказ о человеке, страдающем дизентерией, которому бактериофаги вернули хорошее здоровье.[15] Именно Д'Эрелль провел много исследований бактериофагов и ввел концепцию фаговая терапия.[16]
Более полувека спустя, в 1969 году, Макс Дельбрюк, Альфред Херши, и Сальвадор Лурия были награждены Нобелевская премия по физиологии и медицине за открытия репликации вирусов и их генетической структуры.[17]
Использует
Фаговая терапия
Было обнаружено, что фаги являются антибактериальными агентами и использовались в первых Советский Республика Грузия (впервые здесь Георгий Элиава с помощью одного из первооткрывателей бактериофагов, Феликс д'Эрель ) в 1920-х и 1930-х годах для лечения бактериальных инфекций. Они имели широкое применение, в том числе обращение с солдатами в Красная армия. Однако на Западе от них отказались для широкого использования по нескольким причинам:
- Были открыты и широко распространены антибиотики. Их было легче изготовить, хранить и выписывать.
- Были проведены медицинские испытания фагов, но из-за недостатка понимания возникли вопросы о достоверности этих испытаний.[18]
- Публикация исследований в Советском Союзе была в основном в русский или же Грузинские языки и в течение многих лет за ним не следили на международном уровне.
Использование фагов продолжалось с конца Холодная война в России,[19] Грузия и другие регионы Центральной и Восточной Европы. Первый регулируемый, рандомизированный, двойной слепой клиническое испытание была опубликована в журнале Journal of Wound Care в июне 2009 года, в котором оценивалась безопасность и эффективность коктейля с бактериофагами для лечения инфицированных венозных язв на ногах у людей.[20] FDA одобрило исследование как клиническое испытание фазы I. Результаты исследования продемонстрировали безопасность терапевтического применения бактериофагов, но не показали эффективности. Авторы объяснили, что использование определенных химикатов, которые входят в стандартный уход за ранами (например, лактоферрин или серебро) могли повлиять на жизнеспособность бактериофага.[20] Вскоре после этого было проведено еще одно контролируемое клиническое испытание в Западной Европе (лечение ушных инфекций, вызванных Синегнойная палочка) был опубликован в журнале Clinical Otolaryngology в августе 2009 года.[21] В исследовании сделан вывод о том, что препараты бактериофага безопасны и эффективны для лечения хронических инфекций уха у людей. Кроме того, были проведены многочисленные экспериментальные клинические испытания на животных и другие экспериментальные клинические испытания, оценивающие эффективность бактериофагов при различных заболеваниях, таких как инфицированные ожоги и раны, а также инфекции легких, связанные с кистозным фиброзом.[21]
Между тем, исследователи бактериофагов разрабатывают искусственно созданные вирусы для преодоления устойчивость к антибиотикам, и конструирование фаговых генов, ответственных за кодирование ферментов, которые разрушают матрикс биопленок, структурные белки фага и ферменты, ответственные за лизис стенки бактериальной клетки.[3][4][5] Были получены результаты, показывающие, что фаги Т4, небольшие по размеру и короткохвостые, могут быть полезны при обнаружении Кишечная палочка в теле человека.[22]
Терапевтическая эффективность коктейля фагов оценивалась на модели мышей с носовой инфекцией с множественной лекарственной устойчивостью (МЛУ). А. бауманний. Мыши, получавшие коктейль с фагами, показали в 2,3 раза более высокую выживаемость, чем мыши, не получавшие лечения, через семь дней после заражения.[23] В 2017 г. пациент с поджелудочной железой с компромиссом по МЛУ. А. бауманний был прописан несколько антибиотиков, несмотря на это, в течение четырех месяцев состояние пациента продолжало ухудшаться. Без эффективных антибиотиков пациенту была проведена фаговая терапия с использованием фагового коктейля, содержащего девять различных фагов, эффективность которых против МЛУ была подтверждена. А. бауманний. После этого лечения нисходящая клиническая траектория пациента изменилась и выздоровела.[24]
Д'Эрелль «быстро узнал, что бактериофаги встречаются везде, где процветают бактерии: в сточных водах, в реках, собирающих сточные воды из труб, и в стуле выздоравливающих пациентов».[25] Сюда входят реки, которые традиционно считались исцеляющими, в том числе и индийские. Ганг Река.[26]
Другой
Пищевая промышленность - С 2006 г. Управление по санитарному надзору за качеством пищевых продуктов и медикаментов США (FDA) и Министерство сельского хозяйства США (USDA) одобрило несколько продуктов с бактериофагами. LMP-102 (Intralytix) был одобрен для обработки готовых к употреблению (RTE) птицы и мясных продуктов. В том же году FDA одобрило LISTEX (разработанный и произведенный Микрео ) использование бактериофагов на сыре для уничтожения Listeria monocytogenes бактерии, чтобы дать им общепризнанно безопасным (GRAS) статус.[27] В июле 2007 года один и тот же бактериофаг был одобрен для использования во всех пищевых продуктах.[28] В 2011 году Министерство сельского хозяйства США подтвердило, что LISTEX является средством для обработки чистой этикетки и включено в USDA.[29] Исследования в области безопасности пищевых продуктов продолжаются, чтобы выяснить, являются ли литические фаги жизнеспособным вариантом для борьбы с другими патогенами пищевого происхождения в различных пищевых продуктах.
Молочная промышленность - Бактериофаги, присутствующие в окружающей среде, могут вызывать сбои ферментации сырных заквасок. Чтобы избежать этого, можно использовать заквасочные культуры со смешанными штаммами и режимы ротации культур.[30]
Диагностика - В 2011 году FDA одобрило первый продукт на основе бактериофагов для использования в диагностике in vitro.[31] В тесте на культуру крови KeyPath MRSA / MSSA используется смесь бактериофагов для обнаружения Золотистый стафилококк в положительных посевах крови и определить метициллин сопротивление или восприимчивость. Тест возвращает результаты примерно через пять часов, по сравнению с двумя-тремя днями для стандартных методов идентификации микробов и определения чувствительности. Это был первый ускоренный тест на чувствительность к антибиотикам, одобренный FDA.[32]
Противодействие биологическому оружию и токсинам - Государственные органы на Западе в течение нескольких лет стремились Грузия и бывший Советский союз за помощь в использовании фагов для противодействия биологическому оружию и токсинам, таким как сибирская язва и ботулизм.[33] Исследования продолжаются среди исследовательских групп в США. Другие применения включают распыление в садоводстве для защиты растений и овощных продуктов от гниения и распространения бактериальных заболеваний. Другие области применения бактериофагов - это биоциды для поверхностей из окружающей среды, например, в больницах, и в качестве профилактических средств лечения катетеров и медицинских устройств перед их использованием в клинических условиях. В настоящее время существует технология нанесения фагов на сухие поверхности, например, на униформу, занавески или даже хирургические швы. Клинические испытания, опубликованные в Клиническая отоларингология[21] показать успехи в ветеринарном лечении домашних собак с отит.
В СЕПТИК Метод обнаружения и идентификации бактерий использует ионную эмиссию и ее динамику во время фаговой инфекции и обеспечивает высокую специфичность и скорость обнаружения.[34]
Фаговый дисплей представляет собой другое использование фагов с участием библиотеки фагов с вариабельным пептидом, связанным с поверхностным белком. Каждый фаговый геном кодирует вариант белка, отображаемый на его поверхности (отсюда и название), обеспечивая связь между вариантом пептида и его кодирующим геном. Варианты фагов из библиотеки могут быть выбраны по их аффинности связывания с иммобилизованной молекулой (например, токсином ботулизма) для ее нейтрализации. Связанные отобранные фаги могут быть размножены путем повторного заражения чувствительного бактериального штамма, что позволяет им извлекать закодированные в них пептиды для дальнейшего изучения.[35]
Открытие противомикробных препаратов - Фаговые белки часто обладают антимикробной активностью и могут служить пептидомиметики, т.е. препараты, имитирующие пептиды.[36] Фаг-лигандная технология использует фаговые белки для различных целей, таких как связывание бактерий и бактериальных компонентов (например, эндотоксин ) и лизис бактерий.[37]
Фундаментальные исследования - Бактериофаги важны модельные организмы для изучения принципов эволюция и экология.[38]
Репликация

Бактериофаги могут иметь литический цикл или лизогенный цикл. С литические фаги такой как Фаг Т4, бактериальные клетки разрушаются (лизируются) и разрушаются после немедленной репликации вириона. Как только клетка разрушается, потомство фага может найти новых хозяев для заражения. Литические фаги больше подходят для фаговая терапия. Некоторые литические фаги подвергаются феномену, известному как ингибирование лизиса, когда законченное фаговое потомство не будет немедленно лизироваться из клетки, если внеклеточные концентрации фага высоки. Этот механизм не идентичен механизму умеренный фаг бездействует и обычно носит временный характер.
Напротив, лизогенный цикл не приводит к немедленному лизису клетки-хозяина. Фаги, способные подвергаться лизогении, известны как умеренные фаги. Их вирусный геном будет интегрироваться с ДНК хозяина и реплицироваться вместе с ней относительно безвредно или даже может стать плазмида. Вирус остается бездействующим до тех пор, пока условия хозяина не ухудшатся, возможно, из-за истощения питательных веществ, тогда эндогенный фаги (известные как профаги ) становятся активными. В этот момент они запускают репродуктивный цикл, что приводит к лизису клетки-хозяина. Поскольку лизогенный цикл позволяет клетке-хозяину продолжать выживать и воспроизводиться, вирус реплицируется во всех потомках клетки. Примером бактериофага, который, как известно, следует лизогенному циклу и литическому циклу, является фаг лямбда из E. coli.[39]
Иногда профаги могут приносить пользу бактериям-хозяевам, пока они находятся в спящем состоянии, добавляя новые функции к бактериям. геном, в явлении, называемом лизогенная конверсия. Примеры - преобразование безвредных штаммов Коринебактерии дифтерии или же Холерный вибрион бактериофагами, к высоковирулентным, вызывающим дифтерия или же холера, соответственно.[40][41] Были предложены стратегии борьбы с некоторыми бактериальными инфекциями путем нацеливания на эти кодирующие токсин профаги.[42]
Привязанность и проникновение
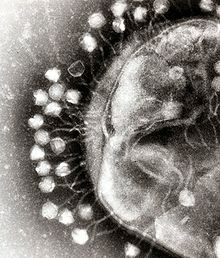
Бактериальные клетки защищены клеточной стенкой из полисахариды, которые являются важными факторами вирулентности, защищающими бактериальные клетки как от иммунной защиты хозяина, так и от антибиотики.[43] Чтобы проникнуть в клетку-хозяин, бактериофаги прикрепляются к специфическим рецепторам на поверхности бактерий, включая липополисахариды, тейхоевые кислоты, белки, или даже жгутики. Эта специфичность означает, что бактериофаг может инфицировать только определенные бактерии, несущие рецепторы, с которыми они могут связываться, что, в свою очередь, определяет круг хозяев фага. Ферменты, разлагающие полисахариды, такие как эндолизины, представляют собой связанные с вирионом белки, которые ферментативно разрушают внешний слой капсулы их хозяев на начальном этапе строго запрограммированного процесса инфицирования фагом. .[44] Поскольку вирионы фагов не перемещаются независимо, они должны полагаться на случайные встречи с правильными рецепторами в растворе, такими как кровь, лимфатическая циркуляция, орошение, почвенная вода и т. Д.
Миовирусные бактериофаги используют шприц для подкожных инъекций -подобное движение для введения своего генетического материала в клетку. После контакта с соответствующим рецептором хвостовые волокна изгибаются, чтобы приблизить базовую пластину к поверхности клетки. Это называется обратимым связыванием. После полного присоединения инициируется необратимое связывание, и хвост сокращается, возможно, с помощью АТФ, присутствует в хвосте,[4] инъекция генетического материала через бактериальную мембрану.[45] Инъекция осуществляется посредством своего рода изгибающего движения в стержне, когда он идет в сторону, сокращается ближе к ячейке и толкается обратно вверх. У подовирусов отсутствует удлиненная хвостовая оболочка, как у миовируса, поэтому вместо этого они используют свои маленькие зубчатые волокна хвоста ферментативно, чтобы разрушить часть клеточной мембраны перед вставкой своего генетического материала.
Синтез белков и нуклеиновой кислоты
Через несколько минут бактериальный рибосомы начать перевод вирусной мРНК в белок. Для фагов на основе РНК РНК-репликаза синтезируется на ранней стадии процесса. Белки изменяют бактериальную РНК-полимераза поэтому он предпочтительно транскрибирует вирусную мРНК. Нормальный синтез белков и нуклеиновых кислот в организме хозяина нарушается, и вместо этого он вынужден производить вирусные продукты. Эти продукты становятся частью новых вирионов внутри клетки, вспомогательных белков, которые вносят вклад в сборку новых вирионов, или белков, участвующих в клетке. лизис. В 1972 г. Уолтер Фирс (Гентский университет, Бельгия ) был первым, кто установил полную нуклеотидную последовательность гена, а в 1976 г. - вирусного генома бактериофаг MS2.[46] Некоторые бактериофаги дцДНК кодируют рибосомные белки, которые, как считается, модулируют трансляцию белка во время фаговой инфекции.[47]
Сборка вириона
В случае Фаг Т4, в создании новых вирусных частиц участвуют вспомогательные белки. Сначала собираются опорные плиты, а потом на них надстраиваются хвосты. Головные капсиды, сконструированные отдельно, самопроизвольно собираются вместе с хвостами. ДНК эффективно упакована в головах. Весь процесс занимает около 15 минут.
Выпуск вирионов
Фаги могут высвобождаться посредством лизиса клеток, экструзии или, в некоторых случаях, почкования. Лизис хвостатыми фагами достигается ферментом, называемым эндолизин, который атакует и разрушает клеточную стенку пептидогликан. Совершенно другой тип фага, нитчатый фаг, заставляют клетку-хозяин постоянно секретировать новые вирусные частицы. Освободившиеся вирионы описываются как свободные и, если они не являются дефектными, способны заразить новую бактерию. Почкование связано с определенными Микоплазма фаги. В отличие от высвобождения вириона, фаги, проявляющие лизогенный цикла не убивают хозяина, а, скорее, становятся постоянными жителями, поскольку профаг.
Коммуникация
Исследования 2017 года показали, что бактериофаг Φ3T вырабатывает короткий вирусный белок, который сигнализирует другим бактериофагам о том, что они бездействуют, а не убивают бактерию-хозяин. Арбитриум - так назвали этот белок открывшие его исследователи.[48][49]
Структура генома
Учитывая миллионы различных фагов в окружающей среде, геномы фагов бывают самых разных форм и размеров. РНК-фаг, такой как MS2 имеют самый маленький геном, всего в несколько килобаз. Однако некоторые ДНК-фаги, такие как Т4 могут иметь большие геномы с сотнями генов; размер и форма капсид варьируется вместе с размером генома.[50] Наиболее крупные геномы бактериофагов достигают размера 735 т.п.н.[51]
Геномы бактериофагов могут быть мозаика, т.е. геном многих видов фагов, по-видимому, состоит из множества отдельных модулей. Эти модули могут быть обнаружены у других видов фагов в другом расположении. Микобактериофаги, бактериофаги с микобактериальный хозяева предоставили прекрасные примеры этого мозаицизма. У этих микобактериофагов генетический ассортимент может быть результатом повторяющихся случаев сайт-специфическая рекомбинация и незаконная рекомбинация (результат приобретения фаговым геномом генетических последовательностей бактериального хозяина).[52] Эволюционные механизмы, формирующие геномы бактериальных вирусов, различаются в зависимости от семейства и зависят от типа нуклеиновой кислоты, характеристик структуры вириона, а также от режима жизненного цикла вируса.[53]
Системная биология
Фаги часто оказывают сильное воздействие на хозяев. Как следствие, картина транскрипции инфицированной бактерии может значительно измениться. Например, заражение Синегнойная палочка под действием умеренного фага PaP3 изменил экспрессию 38% (2160/5633) генов своего хозяина. Многие из этих эффектов, вероятно, являются косвенными, поэтому задача состоит в том, чтобы определить прямые взаимодействия между бактериями и фагами.[54]
Было предпринято несколько попыток сопоставить белок-белковые взаимодействия среди фага и их хозяина. Например, было обнаружено, что лямбда бактериофага взаимодействует со своим хозяином, Кишечная палочка, на 31 взаимодействие. Однако масштабное исследование выявило 62 взаимодействия, большинство из которых были новыми. Опять же, значение многих из этих взаимодействий остается неясным, но эти исследования показывают, что, скорее всего, существует несколько ключевых взаимодействий и множество косвенных взаимодействий, роль которых остается не охарактеризованной.[55]
В окружающей среде
Метагеномика позволил обнаруживать бактериофаги в воде, что ранее было невозможно.[56]
Также бактериофаги использовались в гидрологический трассировка и моделирование в река системы, особенно там, где поверхностные воды и грунтовые воды происходят взаимодействия. Использование фагов предпочтительнее более традиционных краситель маркер, потому что они значительно меньше поглощаются при прохождении через грунтовые воды и легко обнаруживаются при очень низких концентрациях.[57] Незагрязненная вода может содержать примерно 2 × 108 бактериофагов на мл.[58]
Считается, что бактериофаги вносят значительный вклад в горизонтальный перенос генов в естественной среде, в основном через трансдукция, но также через трансформация.[59] Метагеномика исследования также показали, что виромы из различных сред содержат гены устойчивости к антибиотикам, в том числе те, которые могут множественная лекарственная устойчивость.[60]
Модельные бактериофаги
Следующие бактериофаги широко изучаются:
Смотрите также
- Вирофаг, вирусы, заражающие другие вирусы
- Бактериоядное животное
- CrAssphage
- ДНК-вирусы
- Экология фагов
- Монографии по фагам (исчерпывающий список фаговых и связанных с фагами монографий, с 1921 г. по настоящее время)
- Полифаг
- РНК-вирусы
- Трансдукция
- Вириома
- CRISPR
- Фагемид
Рекомендации
- ^ а б МакГрат С. и ван Синдерен Д. (редакторы). (2007). Бактериофаг: генетика и молекулярная биология (1-е изд.). Caister Academic Press. ISBN 978-1-904455-14-1.
- ^ «Новая фаговая терапия спасает пациента с бактериальной инфекцией с множественной лекарственной устойчивостью». UC Health - Калифорнийский университет в Сан-Диего. Получено 13 мая 2018.
- ^ а б Wommack, K. E .; Колвелл, Р. Р. (2000). «Вириопланктон: вирусы в водных экосистемах». Обзоры микробиологии и молекулярной биологии. 64 (1): 69–114. Дои:10.1128 / MMBR.64.1.69-114.2000. ЧВК 98987. PMID 10704475.
- ^ а б c Прескотт, Л. (1993). Микробиология, Wm. C. Brown Publishers, ISBN 0-697-01372-3
- ^ а б BBC Horizon (1997): Вирус, который лечит - Документальный фильм об истории фаговой медицины в России и на Западе
- ^ Боррелл, Брендан (август 2012 г.). «Научный разговор: Фаговый фактор». Scientific American. С. 80–83.
- ^ Кин, Э. С. (2012). «Фаговая терапия: концепция лечения». Границы микробиологии. 3: 238. Дои:10.3389 / fmicb.2012.00238. ЧВК 3400130. PMID 22833738.
- ^ «Бактерии и бактериофаги вступают в сговор, образуя клинически фрустрирующие биопленки».
- ^ Элина Лаанто, Сари Мянтинен, Луиджи Де Колибус, Дженни Марьякангас, Эшли Гиллум, Дэвид И. Стюарт, Янне Дж. Равантти, Юха Хуйсконен, Лотта-Риина Сундберг: Вирус, обнаруженный в северном озере, связывает вирусы оцДНК и дцДНК. В: Proceedings of the National Academy of Sciences 114 (31), июль 2017 г., DOI: 10.1073 / pnas.1703834114
- ^ Кришнамурти SR, Ван Д. (2018). «Обширная консервация прокариотических сайтов связывания рибосом в известных и новых пикобирнавирусах». Вирусология. 516: 108–114. Дои:10.1016 / j.virol.2018.01.006. PMID 29346073.
- ^ Кэтрин М. Кауфман, Фатима А. Хуссейн, Джой Ян, Филип Аревало, Джулия М. Браун, Уильям К. Чанг, Дэвид ВанИнсберге, Джозеф Эльшербини, Радхи С. Шарма, Майкл Б. Катлер, Либуша Келли, Мартин Ф. Польц: Основная линия нехвостых вирусов дцДНК как нераспознанных убийц морских бактерий. В: Nature Vol. 554. С. 118–122. 24 января 2018 г. DOI: 10.1038 / природа25474
- ^ Ханкин Э. Х. (1896). "Бактерицидное действие воды" Жамна и Ганж на вибрионе холеры ". Annales de l'Institut Pasteur (На французском). 10: 511–523.
- ^ Творт, Ф. У. (1915). «Исследование природы ультрамикроскопических вирусов». Ланцет. 186 (4814): 1241–1243. Дои:10.1016 / S0140-6736 (01) 20383-3.
- ^ д'Эрель, Феликс (1917). "Sur un microbe invisible antagoniste des bacilles dysentériques" (PDF). Comptes Rendus de l'Académie des Sciences de Paris. 165: 373–5. В архиве (PDF) из оригинала 11 мая 2011 г.. Получено 5 сентября 2010.
- ^ д'Эрель, Феликс (1949). «Бактериофаг» (PDF). Новости науки. 14: 44–59. Получено 5 сентября 2010.
- ^ Кин ЕС (2012). «Феликс д'Эрелль и наше микробное будущее». Будущая микробиология. 7 (12): 1337–1339. Дои:10.2217 / fmb.12.115. PMID 23231482.
- ^ "Нобелевская премия по физиологии и медицине 1969 г.". Нобелевский фонд. Получено 28 июля 2007.
- ^ Каттер, Элизабет; Де Вос, Даниэль; Гвасалия, Гурам; Алавидзе, Земфира; Гогохия, Лаша; Куль, Сара; Абедон, Стивен (1 января 2010 г.). «Фаготерапия в клинической практике: лечение инфекций человека». Текущая фармацевтическая биотехнология. 11 (1): 69–86. Дои:10.2174/138920110790725401. PMID 20214609. S2CID 31626252.
- ^ Сергей Головин Бактериофаги: убийцы в роли спасителей // Наука и жизнь. - 2017. - № 6. - С. 26–33
- ^ а б Rhoads, DD; Wolcott, RD; Кусковский, Массачусетс; Wolcott, BM; Уорд, LS; Сулаквелидзе, А (июнь 2009 г.). «Бактериофаговая терапия венозных язв ног у людей: результаты исследования безопасности фазы I». Журнал ухода за ранами. 18 (6): 237–8, 240–3. Дои:10.12968 / jowc.2009.18.6.42801. PMID 19661847.
- ^ а б c Райт, А .; Hawkins, C.H .; Änggård, E.E .; Харпер, Д. (Август 2009 г.). "Контролируемое клиническое испытание терапевтического препарата бактериофага при хроническом отите, вызванном устойчивостью к антибиотикам. Синегнойная палочка; предварительный отчет об эффективности ". Клиническая отоларингология. 34 (4): 349–357. Дои:10.1111 / j.1749-4486.2009.01973.x. PMID 19673983.
- ^ Тавил, Нэнси (апрель 2012 г.). «Обнаружение поверхностного плазмонного резонанса E. coli и устойчивых к математике бактериофагов S. aureus». PLOS Genetics. 3 (5): e78. Дои:10.1371 / journal.pgen.0030078. ЧВК 1877875. PMID 17530925.
- ^ Ча, Кёнгеун; О, Hynu K .; Jang, Jae Y .; Джо, Юнёль; Kim, Won K .; Ha, Geon U .; Ko, Kwan S .; Мён, Хиджун (10 апреля 2018 г.). «Характеристика двух новых бактериофагов, заражающих Acinetobacter baumannii с множественной лекарственной устойчивостью (МЛУ), и оценка их терапевтической эффективности in vivo». Границы микробиологии. 9: 696. Дои:10.3389 / fmicb.2018.00696. ISSN 1664-302X. ЧВК 5932359. PMID 29755420.
- ^ Schooley, Роберт Т .; Бисвас, Бисваджит; Гилл, Джейсон Дж .; Эрнандес-Моралес, Адриана; Ланкастер, Джейкоб; Арендодатель, Лорен; Барр, Джереми Дж .; Рид, Шэрон Л .; Ровер, Форест (октябрь 2017 г.). «Разработка и использование персонализированных лечебных коктейлей на основе бактериофагов для лечения пациента с распространенной устойчивой инфекцией Acinetobacter baumannii». Противомикробные препараты и химиотерапия. 61 (10). Дои:10.1128 / AAC.00954-17. ISSN 0066-4804. ЧВК 5610518. PMID 28807909.
- ^ Кучмент, Анна (2012), Забытое лекарство: прошлое и будущее фаговой терапии, Springer, стр. 11, ISBN 978-1-4614-0250-3
- ^ Дересинский, Стан (15 апреля 2009 г.). «Бактериофаговая терапия: использование более мелких блох» (PDF). Клинические инфекционные болезни. 48 (8): 1096–1101. Дои:10.1086/597405. PMID 19275495.
- ^ FDA / CFSAN США: Ответное письмо агентства, уведомление GRAS № 000198
- ^ (FDA / CFSAN США: Ответное письмо агентства, уведомление GRAS № 000218)
- ^ Директива FSIS 7120 В архиве 18 октября 2011 г. Wayback Machine
- ^ Атамер, Зейнеп; Самтлебе, Мейке; Неве, Хорст; Дж. Хеллер, Кнут; Хинрихс, Йорг (16 июля 2013 г.). «Обзор: устранение бактериофагов в сыворотке и сывороточных продуктах». Границы микробиологии. 4: 191. Дои:10.3389 / fmicb.2013.00191. ЧВК 3712493. PMID 23882262.
- ^ FDA 510 (k) Предварительное уведомление
- ^ FDA одобряет первый тест для быстрой диагностики и различения MRSA и MSSA. FDA (6 мая 2011 г.)
- ^ Вайсман, Дарья (25 мая 2007 г.) Изучение сибирской язвы в лаборатории советских времен при западном финансировании. Нью-Йорк Таймс
- ^ Добози-Кинг, М .; Seo, S .; Kim, J.U .; Young, R .; Cheng, M .; Киш, Л. (2005). «Быстрое обнаружение и идентификация бактерий: определение ионного каскада, запускаемого фагом (SEPTIC)» (PDF). Журнал биологической физики и химии. 5: 3–7. Дои:10.4024 / 1050501.jbpc.05.01.
- ^ Смит Г.П., Петренко В.А. (апрель 1997 г.). «Фаговый дисплей». Chem. Rev. 97 (2): 391–410. Дои:10.1021 / cr960065d. PMID 11848876.
- ^ Лю, Цзин; Дехби, Мохаммед; Моек, Грег; Архин, Фрэнсис; Бауда, Паскаль; Бержерон, Доминик; Каллехо, Марио; Ферретти, Винсент; Ха, Нхуан (февраль 2004 г.). «Открытие антимикробных препаратов с помощью геномики бактериофагов». Природа Биотехнологии. 22 (2): 185–191. Дои:10.1038 / nbt932. PMID 14716317. S2CID 9905115.
- ^ Технологические основы Фаго-лигандная технология
- ^ Кин, Э. С. (2014). "Компромиссы в жизнях бактериофагов". Бактериофаг. 4 (1): e28365. Дои:10.4161 / bact.28365. ЧВК 3942329. PMID 24616839.
- ^ Мейсон, Кеннет А., Джонатан Б. Лосос, Сьюзен Р. Сингер, Питер Х. Рэйвен и Джордж Б. Джонсон. (2011). Биология, п. 533. Макгроу-Хилл, Нью-Йорк. ISBN 978-0-07-893649-4.
- ^ Мокроусов I (2009). "Коринебактерии дифтерии: разнообразие генома, популяционная структура и перспективы генотипирования ». Инфекция, генетика и эволюция. 9 (1): 1–15. Дои:10.1016 / j.meegid.2008.09.011. PMID 19007916.
- ^ Чарльз Р.К., Райан Е.Т. (октябрь 2011 г.). «Холера в 21 веке». Современное мнение об инфекционных заболеваниях. 24 (5): 472–7. Дои:10.1097 / QCO.0b013e32834a88af. PMID 21799407. S2CID 6907842.
- ^ Кин, Э. С. (декабрь 2012 г.). «Парадигмы патогенеза: нацеливание на мобильные генетические элементы болезни». Границы клеточной и инфекционной микробиологии. 2: 161. Дои:10.3389 / fcimb.2012.00161. ЧВК 3522046. PMID 23248780.
- ^ Друлис-Кава, Зузанна; Майковска-Скробек, Гражина; MacIejewska, Барбара (2015). «Бактериофаги и белки фагового происхождения - подходы к применению». Современная лекарственная химия. 22 (14): 1757–1773. Дои:10.2174/0929867322666150209152851. ЧВК 4468916. PMID 25666799.
- ^ Габашвили, И .; Хан, С .; Hayes, S .; Сервер, П. (1997). «Полиморфизм бактериофага Т7». Журнал молекулярной биологии. 273 (3): 658–67. Дои:10.1006 / jmbi.1997.1353. PMID 9356254.
- ^ Maghsoodi, A .; Chatterjee, A .; Andricioaei, I .; Перкинс, Северная Каролина (25 ноября 2019 г.). «Как работает оборудование для инъекции фага Т4, включая энергию, силы и динамический путь». Труды Национальной академии наук. 116 (50): 25097–25105. Дои:10.1073 / pnas.1909298116. ISSN 0027-8424. ЧВК 6911207. PMID 31767752.
- ^ Fiers, W .; Contreras, R .; Duerinck, F .; Haegeman, G .; Iserentant, D .; Merregaert, J .; Мин Джоу, Вт .; Molemans, F .; Raeymaekers, A .; Van Den Berghe, A .; Volckaert, G .; Изебаерт, М. (1976). «Полная нуклеотидная последовательность РНК бактериофага MS2: первичная и вторичная структура гена репликазы». Природа. 260 (5551): 500–507. Bibcode:1976Натура.260..500F. Дои:10.1038 / 260500a0. PMID 1264203. S2CID 4289674.
- ^ Mizuno, CM; Guyomar, C; Roux, S; Lavigne, R; Родригес-Валера, Ф; Салливан, МБ; Gillet, R; Forterre, P; Крупович, М (2019). «Многочисленные культивируемые и некультивируемые вирусы кодируют рибосомные белки». Nature Communications. 10 (1): 752. Bibcode:2019НатКо..10..752M. Дои:10.1038 / s41467-019-08672-6. ЧВК 6375957. PMID 30765709.
- ^ Каллавей, Юэн (2017). «Вы говорите о вирусе? Фаги отправляют химические сообщения». Природа. Дои:10.1038 / природа.2017.21313.
- ^ Эрез, Зоар; Стейнбергер-Леви, Ида; Шамир, Майя; Дорон, Шани; Стокар-Авихаил, Авигайль; Пелег, Йоав; Меламед, Сара; Ливитт, Азита; Савидор, Алон; Олбек, Шира; Амитаи, Гил; Сорек, Ротем (26 января 2017 г.). «Связь между вирусами определяет решения о лизисе – лизогении». Природа. 541 (7638): 488–493. Bibcode:2017Натура.541..488E. Дои:10.1038 / природа21049. ISSN 0028-0836. ЧВК 5378303. PMID 28099413.
- ^ Черный, LW; Томас, Дж. А. (2012). Конденсированная структура генома. Успехи экспериментальной медицины и биологии. 726. С. 469–87. Дои:10.1007/978-1-4614-0980-9_21. ISBN 978-1-4614-0979-3. ЧВК 3559133. PMID 22297527.
- ^ Аль-Шайеб, Базем; Сачдева, Рохан; Чен Линь-Син; Уорд, Фред; Мунк, Патрик; Devoto, Audra; Кастель, Синди Дж .; Olm, Matthew R .; Баума-Грегсон, Кейт; Амано, Юки; Он, Кристина (февраль 2020 г.). «Клады огромных фагов со всех экосистем Земли». Природа. 578 (7795): 425–431. Дои:10.1038 / s41586-020-2007-4. ISSN 1476-4687. ЧВК 7162821. PMID 32051592.
- ^ Моррис П., Маринелли Л.Дж., Джейкобс-Сера Д., Хендрикс Р.В., Хатфул Г.Ф. (март 2008 г.). «Геномная характеристика микобактериофага Джайлза: доказательства приобретения фагом ДНК хозяина путем незаконной рекомбинации». Журнал бактериологии. 190 (6): 2172–82. Дои:10.1128 / JB.01657-07. ЧВК 2258872. PMID 18178732.
- ^ Крупович М., Прангишвили Д., Хендрикс Р.В., Бамфорд Д.Х. (декабрь 2011 г.). «Геномика бактериальных и архейных вирусов: динамика в прокариотической виросфере». Обзоры микробиологии и молекулярной биологии. 75 (4): 610–35. Дои:10.1128 / MMBR.00011-11. ЧВК 3232739. PMID 22126996.
- ^ Чжао Икс, Чен С, Шен В, Хуанг Дж, Ле С, Лу С, Ли М, Чжао Ю, Ван Дж, Рао Х, Ли Дж, Шен М, Го К, Ян Ю, Тан Й, Ху Ф (2016) . «Глобальный транскриптомный анализ взаимодействий между Синегнойная палочка и бактериофаг PaP3 ". Научный представитель. 6: 19237. Bibcode:2016НатСР ... 619237Z. Дои:10.1038 / srep19237. ЧВК 4707531. PMID 26750429.
- ^ Blasche S, Wuchty S, Rajagopala SV, Uetz P (2013). «Сеть взаимодействия белков бактериофага лямбда с его хозяином, Escherichia coli». Дж. Вирол. 87 (23): 12745–55. Дои:10.1128 / JVI.02495-13. ЧВК 3838138. PMID 24049175.
- ^ Breitbart M, Саламон П., Андресен Б., Махаффи Дж. М., Сегал А. М., Мид Д., Азам Ф., Ровер Ф. (октябрь 2002 г.). «Геномный анализ некультивируемых морских вирусных сообществ». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 99 (22): 14250–5. Bibcode:2002PNAS ... 9914250B. Дои:10.1073 / pnas.202488399. ЧВК 137870. PMID 12384570.
- ^ Мартин, К. (1988). «Применение методов отслеживания бактериофагов в юго-западной воде». Журнал "Вода и окружающая среда". 2 (6): 638–642. Дои:10.1111 / j.1747-6593.1988.tb01352.x.
- ^ Берг, О. (1989). «Большое количество вирусов, обнаруженных в водной среде». Природа. 340 (6233): 467–468. Bibcode:1989Натура.340..467Б. Дои:10.1038 / 340467a0. PMID 2755508. S2CID 4271861.
- ^ Кин, Эрик С .; Блисковский, Валерий В .; Малагон, Франциско; Бейкер, Джеймс Д .; Принц, Джеффри С .; Клаус, Джеймс С .; Адхья, Sankar L .; Гройсман, Эдуардо А. (2017). "Роман" Суперпредседатель "Бактериофаги способствуют горизонтальному переносу генов путем трансформации". мБио. 8 (1): e02115–16. Дои:10,1128 / мБио.02115-16. ЧВК 5241400. PMID 28096488.
- ^ Лекунберри, Итциар; Субиратс, Джессика; Borrego, Carles M .; Бальказар, Хосе Л. (2017). «Изучение вклада бактериофагов в устойчивость к антибиотикам». Загрязнение окружающей среды. 220 (Pt B): 981–984. Дои:10.1016 / j.envpol.2016.11.059. HDL:10256/14115. PMID 27890586.
- ^ Штраус, Джеймс Х .; Зиншеймер, Роберт Л. (июль 1963 г.). «Очистка и свойства бактериофага MS2 и его рибонуклеиновой кислоты». Журнал молекулярной биологии. 7 (1): 43–54. Дои:10.1016 / S0022-2836 (63) 80017-0. PMID 13978804.
- ^ Миллер, ES; Куттер, Э; Мосиг, G; Arisaka, F; Кунисава, Т; Рюгер, W (март 2003 г.). «Геном бактериофага Т4». Обзоры микробиологии и молекулярной биологии. 67 (1): 86–156, содержание. Дои:10.1128 / MMBR.67.1.86-156.2003. ЧВК 150520. PMID 12626685.
- ^ Ackermann, H.-W .; Криш, Х. М. (6 апреля 2014 г.). «Каталог бактериофагов Т4-типа». Архив вирусологии. 142 (12): 2329–2345. Дои:10.1007 / s007050050246. PMID 9672598. S2CID 39369249.
Библиография
- Хаузер, АР (2016). «Помимо антибиотиков: новые терапевтические подходы к бактериальным инфекциям». Клинические инфекционные болезни. 63 (1): 89–95. Дои:10.1093 / cid / ciw200. ЧВК 4901866. PMID 27025826.
- Strathdee, Штеффани; Паттерсон, Том (2019). Идеальный хищник. Книги Hachette. ISBN 978-0316418089.
внешняя ссылка
- Хойслер, Т. (2006) «Вирусы против супербактерий», Macmillan
- Анимация бактериофага, нацеленного на бактерии E. coli
- Общая информация о бактериофагах на Phage.org
- бактериофаги иллюстрации и геномика
- Бактериофаги захватывают добычу
- Подкаст NPR Science Friday, «Использование фаговых вирусов для борьбы с инфекцией», апрель 2008 г.
- Анимация научно правильного бактериофага Т4, нацеленного на бактерии E. coli
- Анимация Гибридная анимация Медицина для нацеливания на бактериофаг Т4 Кишечная палочка бактерии
- Бактериофаги: что это такое. Презентация профессора Грэма Хатфулла, Питтсбургский университет на YouTube
| Составные части |  | |
|---|---|---|
| Жизненный цикл вирусов | ||
| Генетика | ||
| Хозяин | ||
| Другой | ||
| ||