Теоретическая экология - Theoretical ecology
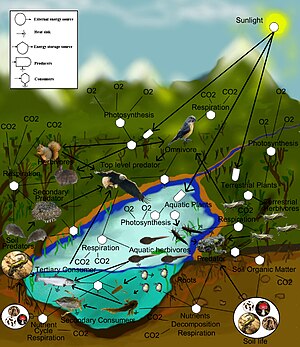

Теоретическая экология это научная дисциплина, посвященная изучению экологические системы используя теоретические методы, такие как простые концептуальные модели, математические модели, компьютерное моделирование, и продвинутый анализ данных. Эффективные модели улучшают понимание мира природы, показывая, как динамика популяций видов часто основана на фундаментальных биологических условиях и процессах. Кроме того, поле направлено на объединение разнообразных эмпирических наблюдений, предполагая, что общие механистические процессы порождают наблюдаемые явления у разных видов и экологической среды. Основываясь на биологически реалистичных предположениях, экологи-теоретики могут открывать новые, не интуитивные представления о естественных процессах. Теоретические результаты часто подтверждаются эмпирическими и наблюдательными исследованиями, раскрывая мощь теоретических методов как в предсказании, так и в понимании зашумленного, разнообразного биологического мира.
Эта область обширна и включает основы прикладной математики, информатики, биологии, статистической физики, генетики, химии, эволюции и биологии сохранения. Теоретическая экология направлена на объяснение разнообразных явлений в науках о жизни, таких как рост населения и динамика, рыболовство, конкуренция, эволюционная теория, эпидемиология, поведение животных и групповая динамика, пищевые полотна, экосистемы, пространственная экология и последствия изменения климата.
Теоретическая экология получила дальнейшие выгоды от появления быстрых вычислительных мощностей, позволяющих анализировать и визуализировать крупномасштабные компьютерные модели экологических явлений. Важно отметить, что эти современные инструменты обеспечивают количественные прогнозы о последствиях антропогенного изменения окружающей среды для разнообразных экологических явлений, таких как вторжение видов, изменение климата, влияние рыболовства и охоты на стабильность продовольственной сети и глобальные цикл углерода.
Подходы к моделированию
Как и в большинстве других наук, математические модели составляют основу современной экологической теории.
- Феноменологические модели: выделяют функциональные и распределительные формы из наблюдаемых закономерностей в данных, или исследователи выбирают функции и распределение, которые являются достаточно гибкими, чтобы соответствовать шаблонам, которые они или другие (полевые или экспериментальные экологи) обнаружили в полевых условиях или в результате экспериментов.[3]
- Механистические модели: моделируют основные процессы напрямую, с функциями и распределениями, которые основаны на теоретических рассуждениях об экологических процессах, представляющих интерес.[3]
Экологические модели могут быть детерминированный или же стохастический.[3]
- Детерминированные модели всегда развиваются одинаково, исходя из заданной отправной точки.[4] Они представляют собой среднее ожидаемое поведение системы, но отсутствуют случайное изменение. Много системная динамика модели детерминированы.
- Стохастические модели позволяют напрямую моделировать случайные возмущения, лежащие в основе экологических систем реального мира. Модели цепей Маркова стохастические.
Виды можно моделировать непрерывно или дискретное время.[5]
- Непрерывное время моделируется с помощью дифференциальные уравнения.
- Дискретное время моделируется с помощью разностные уравнения. Эти модели экологических процессов, которые можно описать как происходящие в течение дискретных временных шагов. Матричная алгебра часто используется для исследования эволюции популяций с возрастной или стадийной структурой. В Матрица Лесли, например, математически представляет дискретное изменение во времени населения с возрастной структурой.[6][7][8]
Для описания реальных процессов экологического воспроизводства одного или нескольких видов часто используются модели, которые можно смоделировать с использованием стохастических методов. ветвящиеся процессы. Примеры - динамика взаимодействующих популяций (хищничество конкуренция и мутуализм ), которые, в зависимости от представляющих интерес видов, лучше всего моделировать в непрерывном или дискретном времени. Другие примеры таких моделей можно найти в области математическая эпидемиология где моделируемые динамические отношения хозяин-патоген взаимодействия.[5]
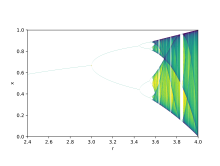
Теория бифуркации используется для иллюстрации того, как небольшие изменения в значениях параметров могут приводить к резко различающимся долгосрочным результатам, математический факт, который можно использовать для объяснения резких экологических различий, которые возникают в качественно очень похожих системах.[9] Логистические карты находятся полиномиальные отображения, и часто цитируются как архетипические примеры того, как хаотичное поведение может возникнуть из очень простых нелинейный динамические уравнения. Карты были популяризированы в оригинальной статье 1976 года теоретика-эколога. Роберт Мэй.[10] Уравнение разности предназначено для улавливания двух эффектов воспроизводства и голода.
В 1930 г. Р.А. Фишер опубликовал свой классический Генетическая теория естественного отбора, который представил идею о том, что частотно-зависимая приспособленность привносит стратегический аспект в эволюция, где выгодами для конкретного организма, возникающими в результате взаимодействия всех соответствующих организмов, является количество жизнеспособных потомков этого организма.[11] В 1961 г. Ричард Левонтин применил теорию игр к эволюционной биологии в своем Эволюция и теория игр,[12]за которым следует Джон Мейнард Смит, который в своей основополагающей статье 1972 года «Теория игр и эволюция борьбы»,[13] определил концепцию эволюционно устойчивая стратегия.
Поскольку экологические системы обычно нелинейный, они часто не могут быть решены аналитически, и для получения разумных результатов необходимо использовать нелинейные, стохастические и вычислительные методы. Один из классов вычислительных моделей, который становится все более популярным, - это агент-ориентированные модели. Эти модели могут моделировать действия и взаимодействия множества разнородных организмов там, где более традиционные аналитические методы неадекватны. Прикладная теоретическая экология дает результаты, которые используются в реальном мире. Например, теория оптимального сбора урожая опирается на методы оптимизации, разработанные в экономике, информатике и исследованиях операций, и широко используется в рыболовство.[14]
Экология населения
Экология населения является подполе экология что касается динамики разновидность население и как эти популяции взаимодействуют с среда.[15] Это исследование того, как размеры населения Виды, живущие вместе в группах, меняются во времени и пространстве, и это был один из первых аспектов экологии, который был изучен и смоделирован математически.
Экспоненциальный рост
Самый простой способ моделирования динамики популяции - это предположить, что темпы роста популяции зависят только от численности популяции в это время и скорости роста организма на душу населения. Другими словами, если количество особей в популяции в момент времени t равно N (t), то скорость роста популяции определяется по формуле:

где r - скорость роста на душу населения или собственная скорость роста организма. Его также можно описать как r = b-d, где b и d - неизменные во времени коэффициенты рождаемости и смертности на душу населения, соответственно. Этот первый заказ линейное дифференциальное уравнение можно решить, чтобы получить решение
- ,

траектория, известная как Мальтузианское развитие, после Томас Мальтус, который впервые описал его динамику в 1798 году. Популяция, в которой наблюдается мальтузианский рост, следует экспоненциальной кривой, где N (0) - начальный размер популяции. Популяция растет, когда r> 0, и уменьшается, когда r <0. Модель наиболее применима в случаях, когда несколько организмов начали колонию и быстро растут без каких-либо ограничений или ограничений, препятствующих их росту (например, бактерии, инокулированные в богатой среде) .
Логистический рост
Модель экспоненциального роста делает ряд предположений, многие из которых часто не выполняются. Например, многие факторы влияют на внутреннюю скорость роста и часто не зависят от времени. Простая модификация экспоненциального роста состоит в том, чтобы предположить, что собственная скорость роста зависит от размера популяции. Это разумно: чем больше численность населения, тем меньше доступных ресурсов, что может привести к снижению рождаемости и более высокой смертности. Следовательно, мы можем заменить неизменный во времени r на r '(t) = (b –a * N (t)) - (d + c * N (t)), где a и c - константы, которые модулируют рождение и смерть. ставки в зависимости от населения (например, внутривидовая конкуренция ). Как a, так и c будут зависеть от других факторов окружающей среды, которые мы можем на данный момент считать постоянными в этой приближенной модели. Дифференциальное уравнение теперь:[16]

Это можно переписать как:[16]

где r = b-d и K = (b-d) / (a + c).
Биологическое значение K становится очевидным при рассмотрении устойчивости состояний равновесия системы. Константа K - это грузоподъемность населения. Равновесия системы равны N = 0 и N = K. Если система линеаризована, можно видеть, что N = 0 является неустойчивым равновесием, а K - устойчивым равновесием.[16]
Структурированный рост населения
Другое предположение модели экспоненциального роста состоит в том, что все особи в популяции идентичны и имеют одинаковые шансы на выживание и воспроизводство. Это неверное предположение для видов со сложной историей жизни. Модель экспоненциального роста может быть изменена, чтобы учесть это, путем отслеживания количества особей в разных возрастных группах (например, одно-, двух- и трехлетние) или разных стадиях (подростки, суб-взрослые и взрослые). ) по отдельности и позволяя отдельным лицам в каждой группе иметь свои собственные коэффициенты выживания и воспроизводства. Общая форма этой модели такова:

куда Nт это вектор количества человек в каждом классе за раз т и L это матрица который содержит вероятность выживания и плодовитость для каждого класса. Матрица L называется Матрица Лесли за возрастной модели, и как матрица Лефковича для сценический модели.[17]
Если значения параметров в L оцениваются на основе демографических данных по конкретной популяции, затем можно использовать структурированную модель для прогнозирования того, будет ли эта популяция расти или сокращаться в долгосрочной перспективе, и какие ожидаемые возрастное распределение внутри населения будет. Это было сделано для ряда видов, включая логгерхед морские черепахи и правые киты.[18][19]
Экология сообщества
Экологическое сообщество - это группа трофически подобных, сочувствующий виды, которые фактически или потенциально конкурируют на определенной территории за одни и те же или аналогичные ресурсы.[20] Взаимодействие между этими видами - первые шаги в анализе более сложной динамики экосистем. Эти взаимодействия формируют распределение и динамику видов. Из этих взаимодействий хищничество является одним из наиболее распространенных видов деятельности популяций.[21]В самом общем смысле хищничество включает взаимодействия хищник-жертва, хозяин-патоген и хозяин-паразитоид.

Взаимодействие хищника и жертвы
Хищная добыча взаимодействия демонстрируют естественные колебания в популяциях как хищников, так и жертв.[21] В 1925 г. американский математик Альфред Дж. Лотка разработал простые уравнения для взаимодействий хищник-жертва в своей книге по биоматематике.[22] В следующем году итальянский математик Вито Вольтерра, провели статистический анализ уловов рыбы в Адриатике.[23] и независимо разработали те же уравнения.[24] Это одна из самых ранних и наиболее известных экологических моделей, известная как Модель Лотка-Вольтерра:


где N - жертва, а P - размер популяции хищников, r - скорость роста жертвы, принимаемая экспоненциальной в отсутствие хищников, α - коэффициент смертности жертвы от хищничества на душу населения (также называемый `` скоростью атаки '' ), c - эффективность превращения жертвы в хищника, а d - экспоненциальный коэффициент смертности хищников при отсутствии какой-либо добычи.
Вольтерра изначально использовал модель для объяснения колебаний численности рыб и акул после ловит рыбу был сокращен во время Первая мировая война. Однако впоследствии уравнения стали применяться более широко.[25] Другие примеры этих моделей включают модель Лотки-Вольтерры заяц на снегоступах и Канадская рысь в Северной Америке,[26] моделирование любого инфекционного заболевания, например недавняя вспышка ОРВИ[27]и биологический контроль Калифорнийская красная шкала введением своего паразитоид, Афитис мелинус.[28]
Надежной и простой альтернативой модели хищник – жертва Лотки-Вольтерры и их распространенным обобщениям, зависящим от жертвы, является соотношение зависимостей или Модель Ардити-Гинзбург.[29] Это две крайности спектра моделей вмешательства хищников. По мнению авторов альтернативной точки зрения, данные показывают, что истинные взаимодействия в природе настолько далеки от экстремума Лотки – Вольтерра на спектре интерференции, что модель можно просто отбросить как неверную. Они намного ближе к зависимому от отношения экстремуму, поэтому, если требуется простая модель, можно использовать модель Ардити – Гинзбурга в качестве первого приближения.[30]
Взаимодействие хозяин-патоген
Второе взаимодействие, взаимодействие хоста и возбудитель, отличается от взаимодействий хищник-жертва тем, что патогены намного меньше по размеру, имеют гораздо более быстрое время генерации и требуют для воспроизводства хозяина. Таким образом, в моделях «хозяин – патоген» отслеживается только популяция хозяина. Обычно используются компартментные модели, которые разделяют популяцию хозяев на группы, такие как восприимчивые, инфицированные и выздоровевшие (SIR).[31]
Взаимодействие хозяин-паразитоид
Третье взаимодействие, взаимодействие хоста и паразитоид, можно проанализировать Модель Николсона – Бейли, который отличается от моделей Lotka-Volterra и SIR дискретностью по времени. Эта модель, как и модель Лотки-Вольтерры, явно отслеживает обе популяции. Обычно в общем виде он гласит:
![{displaystyle N_ {t + 1} = лямбда N_ {t} [1-f (N_ {t}, P_ {t})]}](https://wikimedia.org/api/rest_v1/media/math/render/svg/70346762c7b4aebcd36578c549923bd804870171)

где f (Nт, Пт) описывает вероятность заражения (обычно распределение Пуассона ), λ - скорость роста хозяев на душу населения в отсутствие паразитоидов, а c - эффективность преобразования, как в модели Лотки-Вольтерра.[21]
Конкуренция и мутуализм
При изучении популяций двух видов система уравнений Лотки-Вольтерры широко использовалась для описания динамики поведения между двумя видами, N1 и н2. Примеры включают отношения между D. discoiderum и Кишечная палочка,[32]а также теоретический анализ поведения системы.[33]


Коэффициенты r определяют «базовую» скорость роста каждого вида, а коэффициенты K соответствуют несущей способности. Что действительно может изменить динамику системы, так это члены α. Они описывают характер отношений между двумя видами. Когда α12 отрицательно, это означает, что N2 отрицательно влияет на N1, соревнуясь с ним, охотясь на него или любыми другими возможностями. Когда α12 положительна, однако это означает, что N2 положительно влияет на N1через какое-то мутуалистическое взаимодействие между ними.12 и α21 отрицательны, отношения описываются как конкурентный. В этом случае каждый вид отвлекает от другого, потенциально из-за конкуренции за ограниченные ресурсы.12 и α21 положительны, отношения становятся одним из мутуализм. В этом случае каждый вид приносит пользу другому, так что присутствие одного способствует росту популяции другого.
- Видеть Конкурентные уравнения Лотки – Вольтерра. для дальнейших расширений этой модели.
Нейтральная теория
Единая нейтральная теория это гипотеза, предложенная Стивен Хаббелл в 2001.[20] Гипотеза направлена на объяснение разнообразия и относительной численности видов в экологических сообществах, хотя, как и другие нейтральные теории В экологии гипотеза Хаббелла предполагает, что различия между членами экологического сообщества трофически сходных видов «нейтральны» или не имеют отношения к их успеху. Нейтралитет означает, что при данном трофический уровень в пищевой сети, виды эквивалентны по коэффициентам рождаемости, смертности, скорости распространения и скорости видообразования при измерении на душу населения.[34] Это означает, что биоразнообразие возникает случайно, поскольку каждый вид следует случайная прогулка.[35] Это можно считать нулевая гипотеза к теория ниши. Гипотеза вызвала споры, и некоторые авторы считают ее более сложной версией других нулевых моделей, которая лучше соответствует данным.
Согласно единой нейтральной теории, сложные экологические взаимодействия разрешены между индивидами экологическое сообщество (например, конкуренция и сотрудничество), при условии, что все люди подчиняются одним и тем же правилам. Асимметричные явления, такие как паразитизм и хищничество исключены техническим заданием; но совместные стратегии, такие как роение, и негативное взаимодействие, такое как борьба за ограниченную пищу или свет, разрешено, если все люди ведут себя одинаково. Теория делает прогнозы, которые имеют значение для управления биоразнообразие, особенно управление редкими видами. Он предсказывает существование фундаментальной константы биоразнообразия, обычно записываемой θ, который, по-видимому, управляет видовым богатством в самых разных пространственных и временных масштабах.
Хаббелл основывался на более ранних нейтральных концепциях, включая Макартур & Уилсон теория островная биогеография[20] и Гулд концепции симметрии и нулевых моделей.[34]
Пространственная экология
Биогеография
Биогеография это изучение распределения видов в пространстве и времени. Его цель - выявить, где обитают организмы, в каком количестве и почему они (или не встречаются) в определенной географической области.
Биогеография наиболее пристально наблюдается на островах, что привело к развитию субдисциплины островная биогеография. Эти среды обитания часто являются более управляемыми областями изучения, поскольку они более компактны, чем более крупные экосистемы на материке. В 1967 г. Роберт Макартур и E.O. Уилсон опубликовано Теория островной биогеографии. Это показало, что видовое богатство в районе можно предсказать с точки зрения таких факторов, как площадь ареала, скорость иммиграции и скорость исчезновения.[36] Теория считается одной из основ экологической теории.[37] Применение теории островной биогеографии к фрагменты среды обитания стимулировали развитие областей биология сохранения и ландшафтная экология.[38]
теория r / K-отбора
Концепция популяционной экологии - это теория отбора r / K, одна из первых прогнозных моделей в экологии, используемых для объяснения эволюция жизненного цикла. Предпосылка модели выбора r / K состоит в том, что давление естественного отбора изменяется в зависимости от плотность населения. Например, когда остров впервые колонизируется, плотность особей низкая. Первоначальное увеличение численности популяции не ограничивается конкуренцией, в результате чего остается множество доступных Ресурсы для быстрого роста населения. Эти ранние фазы рост населения опыт независимый от плотности силы естественного отбора, который называется р-выбор. По мере того, как население становится более густонаселенным, оно приближается к пропускной способности острова, что вынуждает людей более активно конкурировать за меньшее количество доступных ресурсов. В условиях скученности население испытывает зависящие от плотности силы естественного отбора, называемые K-выбор.[39][40]

Теория ниши
Метапопуляции
Пространственный анализ экологических систем часто показывает, что допущения, которые действительны для пространственно однородных популяций - и действительно интуитивно понятны - могут больше не действовать, когда рассматриваются мигрирующие субпопуляции, перемещающиеся с одного участка на другой.[42] В простом составе, состоящем из одного вида, субпопуляция может занимать участок, перемещаться с одного участка на другой пустой участок или вымереть, оставив после себя пустой участок. В таком случае пропорция занятых участков может быть представлена как

где m - скорость колонизация, e - скорость вымирание.[43] В этой модели, если e


В этом случае, если e слишком велико, p1 и р2 будет нулевым в устойчивом состоянии. Однако, когда скорость вымирания умеренная, p1 и р2 могут стабильно сосуществовать. Установившееся значение p2 дан кем-то

(п*1 может быть выведено из симметрии). Если e равно нулю, динамика системы благоприятствует видам, которые лучше колонизируют (т. е. имеют более высокое значение m). Это приводит к очень важному результату в теоретической экологии, известному как Гипотеза о промежуточном нарушении, где биоразнообразие (количество видов, которые сосуществуют в популяции) максимизируется, когда нарушение (в данном случае е является прокси) не слишком велико или слишком мало, а находится на промежуточных уровнях.[44]
Форма дифференциальных уравнений, используемых в этом упрощенном подходе к моделированию, может быть изменена. Например:
- Колонизация может зависеть от p линейно (m * (1-p)) в отличие от нелинейного режима m * p * (1-p), описанного выше. Этот способ размножения вида называется «дождем пропагул», когда в каждом поколении в популяцию попадает множество новых особей. В таком сценарии устойчивое состояние, при котором популяция равна нулю, обычно нестабильно.[45]
- Вымирание может нелинейно зависеть от p (e * p * (1-p)) в отличие от линейного (e * p) режима, описанного выше. Это называется «спасательный эффект », И при этом режиме опять же труднее добиться исчезновения популяции.[45]
Модель также может быть расширена до комбинаций четырех возможных линейных или нелинейных зависимостей колонизации и вымирания от p, более подробно описанных в.[46]
Экология экосистемы
Представляем новые элементы, будь то биотический или же абиотический, в экосистемы может быть разрушительным. В некоторых случаях это приводит к экологический коллапс, трофические каскады и гибель многих видов в экосистеме. Абстрактное понятие экологическое здоровье попытки измерить устойчивость и способность восстановления экосистемы; т.е.насколько далеко экосистема от своего устойчивого состояния. Однако часто экосистемы отскакивают от разрушительного агента. Разница между коллапсом и отскоком зависит от токсичность введенного элемента и устойчивость исходной экосистемы.
Если экосистемы регулируются в первую очередь стохастический процессы, посредством которых его последующее состояние будет определяться как предсказуемыми, так и случайными действиями, они могут быть более устойчивыми к внезапным изменениям, чем каждый вид в отдельности. В отсутствие баланс природы видовой состав экосистем будет претерпевать сдвиги, которые будут зависеть от характера изменения, но полный экологический коллапс, вероятно, будет нечастым явлением. В 1997 г. Роберт Уланович использовал теория информации инструменты для описания структуры экосистем с акцентом на взаимная информация (корреляции) в исследуемых системах. Опираясь на эту методологию и предыдущие наблюдения за сложными экосистемами, Уланович описывает подходы к определению уровней стресса в экосистемах и прогнозированию реакции системы на определенные типы изменений в их условиях (например, увеличение или уменьшение потока энергии и эвтрофикация.[47]
Экопат это бесплатный программный пакет для моделирования экосистем, первоначально разработанный NOAA, и широко используется в управлении рыболовством в качестве инструмента для моделирования и визуализации сложных взаимосвязей, существующих в реальных морских экосистемах.
Пищевые полотна
Пищевые полотна обеспечивают основу, в рамках которой может быть организована сложная сеть взаимодействий хищник – жертва. Модель пищевой сети - это сеть пищевые цепи. Каждая пищевая цепочка начинается с основной производитель или же автотроф, организм, такой как растение, который может производить себе пищу. Следующим в цепочке идет организм, который питается первичным продуцентом, и эта цепочка продолжается в виде цепочки последовательных хищников. Организмы в каждой цепи сгруппированы в трофические уровни в зависимости от того, сколько ссылок они удалены от основных производителей. Длина цепочки, или трофический уровень, является мерой количества видов, встречающихся по мере того, как энергия или питательные вещества перемещаются от растений к высшим хищникам.[48] Пищевая энергия течет от одного организма к другому, к следующему и так далее, при этом на каждом уровне теряется некоторая энергия. На данном трофическом уровне может существовать один вид или группа видов с одними и теми же хищниками и жертвами.[49]
В 1927 г. Чарльз Элтон опубликовали влиятельный синтез по использованию пищевых сетей, в результате которого они стали центральной концепцией в экологии.[50] В 1966 году интерес к пищевым сетям возрос после Роберт Пейн экспериментальное и описательное исследование приливных берегов, предполагающее, что сложность трофической сети является ключом к поддержанию видового разнообразия и экологической стабильности.[51] Многие теоретики-экологи, в том числе Сэр Роберт Мэй и Стюарт Пимм, были побуждены этим и другими открытием исследовать математические свойства пищевых сетей. Согласно их анализу, сложные пищевые сети должны быть менее стабильными, чем простые пищевые сети.[1]:75–77[2]:64 Очевидный парадокс между сложностью пищевых сетей, наблюдаемых в природе, и математической хрупкостью моделей пищевых сетей в настоящее время является областью интенсивных исследований и дискуссий. Парадокс может быть частично вызван концептуальными различиями между устойчивостью пищевой сети и равновесием. стабильность пищевой сети.[1][2]
Системная экология
Системная экология можно рассматривать как применение общая теория систем экологии. Требуется целостный и междисциплинарный подход к изучению экологических систем, и в особенности экосистем. Системная экология особенно озабочена тем, как человеческое вмешательство может повлиять на функционирование экосистем. Как и другие области теоретической экологии, он использует и расширяет концепции термодинамика и разрабатывает другие макроскопические описания сложных систем. Также учитывается потоки энергии через различные трофические уровни в экологических сетях. В системной экологии принципы потоков энергии экосистемы формально считаются аналогичными принципам. принципы энергетики. Системная экология также учитывает внешнее влияние экологическая экономика, который в экосистемной экологии обычно не рассматривается.[52] По большей части системная экология - это подраздел экологии экосистемы.
Экофизиология
Это исследование того, как "окружающая среда, как физическая, так и биологическая, взаимодействует с физиологией организма. Оно включает влияние климата и питательных веществ на физиологические процессы как у растений, так и у животных, и особое внимание уделяется масштабам физиологических процессов. с размером организма ».[53][54]
Поведенческая экология
Роевое поведение

Роевое поведение это коллективное поведение демонстрируются животными одинакового размера, которые собираются вместе, возможно, размалывая примерно одно и то же место, или, возможно, миграция в каком-то направлении. Роевое поведение обычно проявляют насекомые, но оно также встречается в стекаться птиц, учеба в школе рыбы и стадное поведение четвероногих. Это сложный возникающий поведение, которое происходит, когда отдельные агенты соблюдайте простые правила поведения.
Недавно был открыт ряд математических моделей, объясняющих многие аспекты эмерджентного поведения. Алгоритмы Swarm следуют Лагранжиан подход или Эйлеров подход.[56] Эйлеров подход рассматривает рой как поле, работая с плотностью роя и определяя свойства среднего поля. Это гидродинамический подход, который может быть полезен для моделирования общей динамики больших скоплений.[57][58][59] Однако большинство моделей работают с лагранжевым подходом, который агент-ориентированная модель следуя за отдельными агентами (точками или частицами), составляющими рой. Модели отдельных частиц могут следовать информации о направлении и расстоянии, которая теряется в подходе Эйлера.[56][60] Примеры включают оптимизация колонии муравьев, самоходные частицы и оптимизация роя частиц
Эволюционная экология
Британский биолог Альфред Рассел Уоллес наиболее известен тем, что независимо предложил теорию эволюция из-за естественный отбор это побудило Чарльз Дарвин опубликовать свою теорию. В своей знаменитой статье 1858 года Уоллес предложил естественный отбор как своего рода механизм обратной связи, который позволяет видам и разновидностям адаптироваться к окружающей среде.[61]
Действие этого принципа точно такое же, как у центробежный регулятор паровой машины, которая проверяет и исправляет любые нарушения почти до того, как они станут очевидными; и точно так же никакой неуравновешенный дефицит в животном царстве никогда не может достигнуть сколько-нибудь заметного размера, потому что он проявится уже на самом первом этапе, затрудняя существование и почти наверняка последующее вымирание.[62]
В кибернетик и антрополог Грегори Бейтсон В 1970-х годах заметил, что, хотя Уоллес писал это только в качестве примера, «вероятно, он сказал самое сильное из того, что было сказано в 19 веке».[63] Следовательно, связь между естественным отбором и теория систем стала областью активных исследований.[61]
Другие теории
В отличие от предыдущих экологических теорий, которые рассматривали наводнения быть катастрофическими событиями, река концепция импульса наводнения утверждает, что годовой пульс паводка является наиболее важным аспектом и наиболее биологически продуктивной особенностью экосистема реки.[64][65]
История
Теоретическая экология опирается на новаторские работы, выполненные Дж. Эвелин Хатчинсон и его ученики. Братья H.T. Odum и E.P. Odum общепризнаны как основоположники современной теоретической экологии. Роберт Макартур принес теорию общественная экология. Дэниел Симберлофф был студентом E.O. Уилсон, с которым МакАртур сотрудничал Теория островной биогеографии, плодотворный труд в развитии теоретической экологии.[66]
Симберлофф добавил статистической строгости к экспериментальной экологии и был ключевой фигурой в SLOSS дебаты о том, что предпочтительнее защищать: один большой или несколько мелких.[67] Это привело к тому, что сторонники Джаред Даймонд правила собрания сообщества, защищающие свои идеи с помощью анализа нейтральных моделей.[67] Симберлофф также сыграл ключевую роль в (все еще продолжающихся) дебатах о полезности коридоров для соединения изолированных резервов.
Стивен Хаббелл и Майкл Розенцвейг объединил теоретические и практические элементы в работы, которые расширили теорию биогеографии Макартура и острова Вильсона - Хаббелл с его Единая нейтральная теория биоразнообразия и биогеографии и Розенцвейг с его «Разнообразием видов в пространстве и времени».
Теоретические и математические экологи
Предварительно можно провести различие между математиками-экологами, экологами, которые применяют математику к экологическим проблемам, и математиками, которые развивают саму математику, которая возникает из экологических проблем.
В этих категориях можно найти некоторых известных теоретиков-экологов:
- Категория: Математические экологи
- Категория: Биологи-теоретики
Журналы
Смотрите также
- Эффект бабочки
- Сложная системная биология
- Теория экологических систем
- Модель экосистемы
- Уравнение интегроразличия - широко используется для моделирования рассредоточение и рост населения
- Ограничение сходства
- Математическая биология
- Динамика населения
- Моделирование населения
- Количественная экология
- Закон Тейлора
- Теоретическая биология
Рекомендации
- ^ а б c Май РМ (2001) Стабильность и сложность модельных экосистем Princeton University Press, перепечатка издания 1973 года с новым предисловием. ISBN 978-0-691-08861-7.
- ^ а б c Pimm SL (2002) Пищевые полотна University of Chicago Press, перепечатка издания 1982 года с новым предисловием. ISBN 978-0-226-66832-1.
- ^ а б c Болкер БМ (2008) Экологические модели и данные в R Princeton University Press, страницы 6–9. ISBN 978-0-691-12522-0.
- ^ Сугихара Г., Май Р. (1990). «Нелинейное прогнозирование как способ отличить хаос от ошибки измерения во временных рядах» (PDF). Природа. 344 (6268): 734–41. Bibcode:1990Натура.344..734S. Дои:10.1038 / 344734a0. PMID 2330029. Архивировано из оригинал (PDF) на 2011-08-14. Получено 2011-05-13.
- ^ а б Soetaert K и Герман PMJ (2009) Практическое руководство по экологическому моделированию Springer. ISBN 978-1-4020-8623-6.
- ^ Грант В.Е. (1986) Системный анализ и моделирование в науках о дикой природе и рыболовстве. Wiley, Университет Миннесоты, стр. 223. ISBN 978-0-471-89236-6.
- ^ Джопп Ф (2011) Моделирование сложной экологической динамики Springer, стр.122. ISBN 978-3-642-05028-2.
- ^ Burk AR (2005) Новые тенденции в исследованиях экологии Nova Publishers, стр. 136. ISBN 978-1-59454-379-1.
- ^ Ма Т. и Ван С. (2005) Теория бифуркаций и приложения World Scientific. ISBN 978-981-256-287-6.
- ^ Мэй, Роберт (1976). Теоретическая экология: принципы и приложения. Blackwell Scientific Publishers. ISBN 978-0-632-00768-4.
- ^ Фишер, Р. А. (1930). Генетическая теория естественного отбора. Оксфорд: пресса Clarendon.
- ^ Р. С. Левонтин (1961). «Эволюция и теория игр». Журнал теоретической биологии. 1 (3): 382–403. Дои:10.1016/0022-5193(61)90038-8. PMID 13761767.
- ^ Джон Мейнард Смит (1974). «Теория игр и эволюция конфликтов животных» (PDF). Журнал теоретической биологии. 47 (1): 209–21. Дои:10.1016/0022-5193(74)90110-6. PMID 4459582.
- ^ Суприатна АК (1998) Теория оптимальной добычи для метапопуляций хищник – жертва Университет Аделаиды, факультет прикладной математики.
- ^ Одум, Евгений П. (1959). Основы экологии (Второе изд.). Филадельфия и Лондон: W. B. Saunders Co., стр.546 с.. ISBN 9780721669410. OCLC 554879.
- ^ а б c Мосс Р., Уотсон А. и Олласон Дж. (1982) Динамика поголовья животных Springer, стр. 52–54. ISBN 978-0-412-22240-5.
- ^ Хэл Касвелл (2001). Матричные модели популяций: построение, анализ и интерпретация. Синауэр.
- ^ Д.Т. Кроуз, Л. Краудер, Х. Касвелл (1987). «Поэтапная модель популяции головастых морских черепах и ее значение для сохранения». Экология. 68 (5): 1412–1423. Дои:10.2307/1939225. JSTOR 1939225.CS1 maint: несколько имен: список авторов (связь)
- ^ М. Фудзивара; Х. Касвелл (2001). «Демография находящегося под угрозой исчезновения североатлантического кита». Природа. 414 (6863): 537–541. Bibcode:2001Натура.414..537F. Дои:10.1038/35107054. PMID 11734852.
- ^ а б c Хаббелл, СП (2001). «Единая нейтральная теория биоразнообразия и биогеографии (МПБ-32)». Архивировано из оригинал на 2011-07-18. Получено 2010-12-16.
- ^ а б c Бонсолл, Майкл Б.; Хасселл, Майкл П. (2007). «Взаимодействие хищника и жертвы». В мае Роберт; Маклин, Анджела (ред.). Теоретическая экология: принципы и приложения (3-е изд.). Издательство Оксфордского университета. С. 46–61.
- ^ Лотка, А.Дж., Элементы физической биологии, Уильямс и Уилкинс, (1925)
- ^ Гоэль, Н. и др., "О модели Вольтерры и других нелинейных моделях взаимодействующих популяций", Academic Press Inc., (1971)
- ^ Вольтерра, V (1926). "Variazioni e fluttuazioni del numero d'individui in specie animali conviventi". Mem. Акад. Линчеи Рома. 2: 31–113.
- ^ Бегон, М .; Harper, J. L .; Таунсенд, К. Р. (1988). Экология: отдельные лица, популяции и сообщества. Blackwell Scientific Publications Inc., Оксфорд, Великобритания.
- ^ К.С. Элтон (1924). «Периодические колебания численности животных - их причины и следствия». Журнал экспериментальной биологии. 2 (1): 119–163.
- ^ Липсич М., Коэн Т., Купер Б., Робинс Дж. М., Ма С., Джеймс Л., Гопалакришна Г., Чу С. К., Тан СС, Самор М. Х., Фисман Д., Мюррей М. (2003). «Динамика передачи и контроль тяжелого острого респираторного синдрома». Наука. 300 (5627): 1966–70. Bibcode:2003Sci ... 300.1966L. Дои:10.1126 / science.1086616. ЧВК 2760158. PMID 12766207.
- ^ Джон Д. Рив; Уильям В. Мердок (1986). «Биологический контроль со стороны паразитоидов Aphytis melinus и стабильность популяции калифорнийской красной чешуи». Журнал экологии животных. 55 (3): 1069–1082. Дои:10.2307/4434. JSTOR 4434.
- ^ Arditi, R .; Гинзбург, Л. (1989). «Взаимодействие в динамике хищник – жертва: зависимость отношения» (PDF). Журнал теоретической биологии. 139 (3): 311–326. Дои:10.1016 / с0022-5193 (89) 80211-5.
- ^ Ардити, Р., Гинзбург, Л. (2012) Как виды взаимодействуют: изменение стандартного взгляда на трофическую экологию Издательство Оксфордского университета. ISBN 9780199913831.
- ^ Гренфелл, Брайан; Килинг, Мэтью (2007). «Динамика инфекционной болезни». В мае Роберт; Маклин, Анджела (ред.). Теоретическая экология: принципы и приложения (3-е изд.). Издательство Оксфордского университета. С. 132–147.
- ^ Х. М. Цучия; Дж. Ф. Дрейк; Дж. Л. Йост и А. Г. Фредриксон (1972). "Взаимодействие хищника и жертвы Dictyostelium discoideum и кишечная палочка в непрерывной культуре1 ". Журнал бактериологии. 110 (3): 1147–53. Дои:10.1128 / JB.110.3.1147-1153.1972. ЧВК 247538. PMID 4555407.
- ^ Такеучи, Ю. (1989). «Кооперативная теория систем и глобальная устойчивость диффузионных моделей». Acta Applicandae Mathematicae. 14 (1–2): 49–57. Дои:10.1007 / BF00046673.
- ^ а б Хаббелл, С. П. (2005). «Нейтральная теория биоразнообразия и биогеографии и Стивен Джей Гулд». Палеобиология. 31: 122–123. Дои:10.1666 / 0094-8373 (2005) 031 [0122: TNTOBA] 2.0.CO; 2.
- ^ Макгилл, Б. Дж. (2003). «Тест единой нейтральной теории биоразнообразия». Природа. 422 (6934): 881–885. Bibcode:2003Натура.422..881М. Дои:10.1038 / природа01583. PMID 12692564.
- ^ Макартур Р. Х. и Уилсон Е. О. (1967) Теория островной биогеографии
- ^ Wiens, J. J .; Донохью, М. Дж. (2004). «Историческая биогеография, экология и видовое богатство» (PDF). Тенденции в экологии и эволюции. 19 (12): 639–644. Дои:10.1016 / j.tree.2004.09.011. PMID 16701326.CS1 maint: ref = harv (связь)
- ^ Это касается британских и американских ученых; ландшафтная экология имеет ярко выраженный генезис среди европейских ученых.
- ^ Впервые представленный в книге MacArthur & Wilson (1967), получившей заметное упоминание в истории и теоретической науке об экологии, Теория островной биогеографии
- ^ Reznick, D .; Bryant, M. J .; Башей, Ф. (2002). «Новый взгляд на r- и K-отбор: роль регулирования популяции в эволюции жизненного цикла» (PDF). Экология. 83 (6): 1509–1520. Дои:10.1890 / 0012-9658 (2002) 083 [1509: RAKSRT] 2.0.CO; 2. ISSN 0012-9658. Архивировано из оригинал (PDF) на 2010-12-30. Получено 2015-01-05.
- ^ Гевин V (2006). «За пределами нейтралитета - экология находит свою нишу». ПЛОС Биол. 4 (8): 1306–1310. Дои:10.1371 / journal.pbio.0040278. ЧВК 1543692. PMID 16895443.
- ^ Ганский I (1999) Экология метапопуляции Издательство Оксфордского университета. ISBN 978-0-19-854065-6.
- ^ Ханский I, Гилпин М (1991). «Динамика метапопуляции: краткая история и концептуальная область» (PDF). Биологический журнал Линнеевского общества. 42 (1–2): 3–16. Дои:10.1111 / j.1095-8312.1991.tb00548.x.
- ^ Кокс CB и Мур PD (2010) Биогеография: экологический и эволюционный подход Джон Уайли и сыновья, стр.146. ISBN 978-0-470-63794-4.
- ^ а б Вандермейер Дж. Х. и Голдберг Д. Е. (2003) Популяционная экология: основные принципы Издательство Принстонского университета, стр. 175–176. ISBN 978-0-691-11441-5.
- ^ Илкка Ханский (1982). «Динамика регионального распространения: гипотеза основных и спутниковых видов». Ойкос. 38 (2): 210–221. Дои:10.2307/3544021. JSTOR 3544021.
- ^ Роберт Уланович (). Экология, восходящая перспектива. Columbia Univ. Нажмите. ISBN 0-231-10828-1.
- ^ Пост, Д. М. (1993). «Длинная и короткая длина пищевой цепи». Тенденции в экологии и эволюции. 17 (6): 269–277. Дои:10.1016 / S0169-5347 (02) 02455-2.
- ^ Джерри Боброу, доктор философии; Стивен Фишер (2009). CliffsNotes CSET: несколько тем (2-е изд.). Джон Уайли и сыновья. п. 283. ISBN 978-0-470-45546-3.
- ^ Элтон С.С. (1927) Экология животных. Переиздано в 2001 году. University of Chicago Press.
- ^ Пейн RT (1966). «Сложность пищевой сети и видовое разнообразие». Американский натуралист. 100 (910): 65–75. Дои:10.1086/282400.
- ^ Р. Л. Китчинг, Системная экология, University of Queensland Press, 1983, стр.9.
- ^ Экофизиология, Природа. Дата обращения: 9 августа 2017 г.
- ^ Петерс, Р. Х. (1986) Экологические последствия размера тела Издательство Кембриджского университета. ISBN 9780521288866
- ^ Бхаттачарья К. и Вичек Т. (2010) «Коллективное принятие решений сплоченными стаями»
- ^ а б Li YX; Lukeman R; Эдельштейн-Кешет Л (2007). «Минимальные механизмы образования школы в самоходных частицах» (PDF). Physica D: нелинейные явления. 237 (5): 699–720. Bibcode:2008PhyD..237..699L. Дои:10.1016 / j.physd.2007.10.009. Архивировано из оригинал (PDF) на 2011-10-01.
- ^ Toner J и Tu Y (1995) "Дальний порядок в двухмерной xy-модели: как птицы летают вместе" Письма ревю, 75 (23)(1995), 4326–4329.
- ^ Топаз C, Бертоцци A (2004). «Модели роения в двухмерной кинематической модели для биологических групп». SIAM J Appl Math. 65 (1): 152–174. Bibcode:2004APS..MAR.t9004T. CiteSeerX 10.1.1.88.3071. Дои:10.1137 / S0036139903437424.
- ^ Топаз С, Бертоцци А, Льюис М (2006). «Нелокальная модель континуума для биологической агрегации». Бык математика биол. 68 (7): 1601–1623. arXiv:q-bio / 0504001. Дои:10.1007 / s11538-006-9088-6. PMID 16858662.
- ^ Каррильо, Дж; Форнасье, М; Тоскани, G (2010). Частичная, кинетическая и гидродинамическая модели роения (PDF). Моделирование и имитация в науке, технике и технологиях. 3. С. 297–336. CiteSeerX 10.1.1.193.5047. Дои:10.1007/978-0-8176-4946-3_12. ISBN 978-0-8176-4945-6.
- ^ а б Смит, Чарльз Х. "Незаконченное дело Уоллеса". Сложность (издательство Wiley Periodicals, Inc.) Том 10, № 2, 2004 г.. Получено 2007-05-11.
- ^ Уоллес, Альфред. "О склонности разновидностей к неограниченному отходу от первоначального типа". Альфред Рассел Уоллес Пейдж, организованный Университет Западного Кентукки. Получено 2007-04-22.
- ^ Бренд, Стюарт. "Ради бога, Маргарет". CoEvolutionary Quarterly, июнь 1976 г.. Получено 2007-04-04.
- ^ Торп, Дж. Х. и Делонг, М. Д. (1994). Модель продуктивности реки: эвристический взгляд на источники углерода и переработку органических веществ в крупных речных экосистемах. Ойкос, 305-308
- ^ Бенке, А.С., Чауби, И., Уорд, Г.М., и Данн, Е.Л. (2000). Динамика пульсаций наводнений в нерегулируемой пойме реки на юго-востоке прибрежной равнины США. Экология, 2730-2741.
- ^ Каддингтон К. и Бейснер Б. Е. (2005) Утраченные экологические парадигмы: пути изменения теории Академическая пресса. ISBN 978-0-12-088459-9.
- ^ а б Soulé ME, Simberloff D (1986). «Что генетика и экология говорят нам о дизайне заповедников?» (PDF). Биологическое сохранение. 35 (1): 19–40. Дои:10.1016 / 0006-3207 (86) 90025-Х. HDL:2027.42/26318.
дальнейшее чтение
- Классический текст Теоретическая экология: принципы и приложения, к Анджела МакЛин и Роберт Мэй. Издание 2007 года опубликовано Oxford University Press. ISBN 978-0-19-920998-9.
- Болкер БМ (2008) Экологические модели и данные в R Издательство Принстонского университета. ISBN 978-0-691-12522-0.
- Дело TJ (2000) Иллюстрированное руководство по теоретической экологии Издательство Оксфордского университета. ISBN 978-0-19-508512-9.
- Касуэлл Х (2000) Матричные модели популяций: построение, анализ и интерпретация, Синауэр, 2-е изд. ISBN 978-0-87893-096-8.
- Эдельштейн-Кешет Л (2005) Математические модели в биологии Общество промышленной и прикладной математики. ISBN 978-0-89871-554-5.
- Готелли, штат Нью-Джерси (2008) Букварь по экологии Sinauer Associates, 4-е изд. ISBN 978-0-87893-318-1.
- Готелли, штат Нью-Джерси, и Эллисон (2005) Учебник по экологической статистике Издательство Sinauer Associates. ISBN 978-0-87893-269-6.
- Гастингс А (1996) Популяционная биология: концепции и модели Springer. ISBN 978-0-387-94853-9.
- Хилборн R&M Кларк (1997) Экологический детектив: модели с данными Издательство Принстонского университета.
- Кокко Х (2007) Моделирование для полевых биологов и других интересных людей Издательство Кембриджского университета. ISBN 978-0-521-83132-1.
- Кот М (2001) Элементы математической экологии Издательство Кембриджского университета. ISBN 978-0-521-00150-2.
- Лоутон Дж. Х. (1999). «Есть ли общие законы в экологии?» (PDF). Ойкос. 84 (2): 177–192. CiteSeerX 10.1.1.331.1173. Дои:10.2307/3546712. JSTOR 3546712. Архивировано из оригинал (PDF) на 2010-06-11.
- Мюррей JD (2002) Математическая биология, Том 1 Springer, 3-е изд. ISBN 978-0-387-95223-9.
- Мюррей JD (2003) Математическая биология, Том 2 Springer, 3-е изд. ISBN 978-0-387-95228-4.
- Пастор Дж (2008) Математическая экология популяций и экосистем Вили-Блэквелл. ISBN 978-1-4051-8811-1.
- Roughgarden J (1998) Учебник по экологической теории Прентис Холл. ISBN 978-0-13-442062-2.
- Уланович Р (1997) Экология: восходящая перспектива Издательство Колумбийского университета.