Экология устья Сан-Франциско - Ecology of the San Francisco Estuary
В Устье Сан-Франциско вместе с Сакраменто - Дельта реки Сан-Хоакин представляет собой сильно измененный экосистема. Регион был в значительной степени реконструирован для удовлетворения потребностей водоснабжения, судоходства, сельского хозяйства и, в последнее время, развития пригородов. Эти потребности вызвали прямые изменения в движении воды и характере ландшафта, а также косвенные изменения в результате внедрения неместные виды. Новые виды изменили архитектуру пищевой сети так же верно, как дамбы изменили ландшафт островов и каналов, которые образуют сложную систему, известную как Дельта.[1]
В этой статье особенно рассматривается экология низкого соленость зона (LSZ) лимана. Реконструкция исторического пищевой сети для LSZ сложно по ряду причин. Во-первых, нет четких сведений о видах, исторически населявших эстуарий. Во-вторых, устье и дельта Сан-Франциско находились в геологическом и гидрологическом переходе на протяжении большей части своей 10 000-летней истории, и поэтому описание «естественного» состояния устья во многом похоже на «попадание в движущуюся цель».[1] Изменение климата, гидрологическая инженерия, меняющиеся потребности в воде и новые интродуцированные виды продолжит изменять конфигурацию трофической сети устья. Эта модель обеспечивает моментальный снимок текущего состояния с примечаниями о недавних изменениях или интродукциях видов, которые изменили конфигурацию пищевой сети. Понимание динамики существующей пищевой сети может оказаться полезным для усилий по восстановлению, направленных на улучшение функционирования и видового разнообразия эстуария.
Физическая география
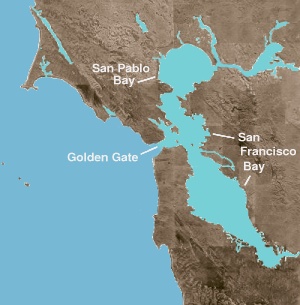
В Залив Сан-Франциско это оба залив и устье. Первый термин относится к любому заливу или бухте, обеспечивающему физическое убежище от открытого океана. Устье - это любой физико-географический объект, где пресная вода встречается с океаном или морем. Северная часть залива представляет собой солоноватое устье, состоящее из ряда физических заливов, в которых преобладают потоки морской и пресной воды. Эти географические объекты, перемещающиеся от соленой воды к пресной (или с запада на восток): Сан-Пабло Бэй, непосредственно к северу от Центрального залива; то Каркинес пролив, узкий, глубокий канал, ведущий к Suisun Bay; и Дельта из Сакраменто и Сан-Хоакин реки.
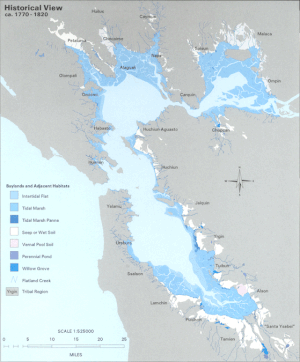
Вплоть до XX века ЛСЗ лимана окаймляли Tule - преобладающая пресная вода водно-болотные угодья. От 80 до 95% этих исторических водно-болотных угодий были заполнены для облегчения землепользования и развития в районе залива.[2] Утрата среды обитания на окраинах пелагическая зона считается, что приводит к потере родного пелагическая рыба видов, увеличивая уязвимость перед хищниками.
В приливной и бентосный эстуарий в настоящее время преобладают илистые равнины которые в значительной степени являются результатом седиментации, происходящей из золотодобыча в Сьерра-Невада в конце 19 века. Тенденция к высокому содержанию наносов изменилась в 1950-х годах с появлением Проект Центральной долины, блокируя большую часть наносов за дамбами, что приводит к ежегодной чистой потере наносов из устья.[3] Таким образом, кажется, что илы медленно отступают, хотя мутность остается чрезвычайно высокой. Высокая мутность воды является причиной уникальных условий, существующих в устье Сан-Франциско, где высокая доступность питательных веществ не приводит к высокому производству фитопланктона. Вместо этого большинство фотосинтезирующих водорослей организмов имеют ограниченный свет.[4]
Дельта также претерпела серьезные изменения. Начиная с 19 века, естественные дамбы были укреплены на постоянство, чтобы защитить сельскохозяйственные угодья от регулярных наводнений. Многие из этих ферм были созданы на торфяных островах, расположенных в центре водных путей Дельты. Интенсивное земледелие окислили высокое содержание углерода в почве, что привело к значительной потере массы почвы. Как следствие, эти острова опустились или погрузились почти на 6 метров ниже уровня моря.[5] Сегодня Дельта состоит из сильно изрезанных водных путей, перемежающихся островами, которые выглядят как «плавучие чаши» с бассейнами, расположенными далеко ниже поверхности воды.[6] Эти острова подвержены высокому риску затопления из-за обрушения дамбы. Ожидается, что последующий сдвиг солености на восток резко изменит экологию всей LSZ эстуария Сан-Франциско.[7]
Гидродинамика

LSZ сосредотачивается около 2 psu (единицы практической солености, измерение солености) и колеблется от 6 до 0,5 пс. Основные поступления пресной воды в эстуарий происходят из стока региональных осадков, рек Сакраменто и Сан-Хоакин.[8]
Приток реки в значительной степени контролируется попусками из водохранилищ выше по течению. Значительная часть этого притока экспортируется из Дельты федеральными властями. Проект Центральной долины и Государственный водный проект на юг Калифорния для сельскохозяйственного и городского использования. Эти изменения устранили большую часть изменений в оттоке через эстуарий (то есть пресной воде, которая делает его Золотые ворота ), создавая более низкий отток зимой и более высокий отток летом, чем исторически наблюдалось в устье реки. В среднем потоки пресной воды в эстуарий составляют 50% исторических потоков.[9] Фитопланктон, зоопланктон, а личинки и взрослые особи рыбы могут быть захвачены насосами для вывоза, оказывая потенциально значительное, но неизвестное влияние на численность этих организмов. Это может быть особенно верно в отношении находящихся под угрозой исчезновения дельта корюшка, небольшая эндемичная рыба; без исключений, за исключением того, что он был описан как чрезвычайно богатый в исторических отчетах.[10] Считается, что дельтовая корюшка мигрирует и нерестится вверх по течению в Дельте в начале лета, подвергая ее яйца и личинки высокому риску уноса.[11] Управление корювой в настоящее время является источником разногласий, поскольку его экология приводит к столкновению несопоставимых потребностей в воде для сохранения, развития и сельского хозяйства в Калифорнии.
Движение воды из устья является сложным и зависит от ряда факторов. Приливные циклы заставлять воду двигаться к Золотым воротам и от них четыре раза за 24 часа.
Используя 2 psu в качестве маркера для зоны низкой солености, направление и величину колебаний можно отслеживать как расстояние в километрах от Золотых ворот или X2. Поскольку положение X2 зависит от ряда физических параметров, включая приток, экспорт и приливы, его положение смещается на многие километры в зависимости от суточного и сезонного цикла; в течение года он может колебаться от залива Сан-Пабло в периоды высокого стока до дельты реки во время летней засухи. Положение X2 тщательно отслеживается и поддерживается путем выпуска воды из резервуаров выше по течению в ожидании экспортного спроса. Это предусмотрено Решением Совета штата по водным ресурсам 1641 и требует, чтобы закачка на уровне штата и на федеральном уровне была сокращена, если X2 смещается к востоку от острова Чиппс (75 километров реки вверх по течению от моста Золотые Ворота) в период с февраля по май или к востоку от Коллинсвилля ( 81 километр реки вверх по течению от моста Золотые Ворота) в январе, июне, июле и августе. (D-1641, стр.150)
Гравитационный Циркуляция заставляет стратифицированную воду с высокой соленостью на глубине течь к суше, а вода с низкой соленостью наверху течет в сторону моря.[12] Эффект гравитационной циркуляции может быть наиболее выражен в периоды высокого потока пресной воды, обеспечивая отрицательную обратную связь для поддержания солевого поля и распределения пелагических организмов в устье.
Перемешивание важно на краю гравитационной циркуляции, обращенном к суше, часто около точки X2, где слой воды становится менее стратифицированным.[13] Зона фиксированного перемешивания возникает на «выступе Бенисия» на восточной оконечности пролива Каркинес, где глубоководный канал резко мелеет, когда он входит в залив Суисун.[14] Перемешивание имеет решающее значение для поддержания солености, поэтому требуются чрезвычайно большие поступления пресной воды для перемещения X2 на небольшое расстояние на запад. Смешивание также помогает пелагическим организмам сохранять свое положение в устье реки.[1] замедление адвекции первичной и вторичной продукции из системы.
Пелагическая зона
Пелагические организмы проводят всю или часть своей жизни на открытой воде, где среда обитания определяется не краями, а физиологической толерантностью к солености и температуре. Зона низкой солености (LSZ) Устье Сан-Франциско представляет собой среду обитания для набора организмов, которые специализируются на выживании в этом уникальном слиянии земной, пресная вода, и морской влияет. Хотя есть много мест обитания с четко выраженными экология которые являются частью устье (включая морские, пресноводные, приливное болото и бентосный ил систем), каждая из которых связана с ЗЗЗ экспортом и импортом пресной воды, питательных веществ, углерода и организмов.[1]
Распространение и численность организмов в ЗЗЗ зависит как от абиотических, так и от биотических факторов. К абиотическим факторам относятся Физическая география и гидрология эстуария, в том числе питательное вещество входы, осадок нагрузка, мутность, экологическая стохастичность, климат и антропогенные воздействия.[8] Абиотические факторы имеют тенденцию стимулировать производство в эстуарной среде и опосредованы биотическими факторами.
Биотические факторы включают потребление питательных веществ и основное производство, вторичное производство из зоопланктон, пищевой сети и трофическая динамика, передача энергии, адвекция и распространение в системе и из нее, выживание и смертность, хищничество, и конкуренция из интродуцированные виды.
Пищевой сети

Исторический пищевой сети устья Сан-Франциско из-за драматических изменений в географии, гидрологии и видовом составе, произошедших в прошлом веке. Однако мониторинг, начатый в 1970-х годах, дает некоторую информацию об исторической динамике пищевой сети. До 1980-х годов в ЗСЗ доминировала трофическая сеть, управляемая фитопланктоном, стабильная популяция мезопланктона, в которой преобладали копепода Eurytemora affinis, и крупный макрозоопланктон, типичный для залива Сан-Франциско. креветка и мизид креветки.[15] Они обеспечивали питание и энергию местных фильтраторов, таких как северный анчоусы (Engraulis mordax), и планктоядные, такие как дельта корюшка и несовершеннолетний лосось.
Изменение пищевой сети исторически было вызвано повышенной мутностью, а в последнее время - интродуцированными видами, как описано в разделах, посвященных первичному и вторичному производству.
Примечательно, что высокий клиренс введенных Амурский моллюск Potamocorbula amurensis население привело к десятикратному снижению плотности планктона, что привело к образованию углеродной ловушки в бентос и предполагаемое увеличение образования обломочных отходов.[16] Предполагается, что эти отходы подпитывают микробную петлю, что приводит к увеличению микрозоопланктона, такого как веслоногие рачки. Лимноитона тетраспина, которые используют коловратки и инфузории.
Эти изменения являются одной из причин сокращения рыбных запасов. Например, северный анчоус, Engraulis mordaxдо 1980-х гг. в Зоне низкой солености было довольно много, пока его ареал в устье не стал ограничиваться Центральным и Южным заливами.[17] Вероятно, это связано с поведенческой реакцией после вселения реки Амур (Potamocorbula amurensis) моллюсков и последующее снижение доступности планктона.
Совсем недавно было описано общее сокращение пелагических организмов (POD), и это стало источником серьезной озабоченности в научных, управленческих и политических кругах.[18]
Несколько ключевых видов, в том числе дельта корюшка, длинноперый корюшка, полосатый окунь, и шэд были объявлены "представляющими интерес видами" из-за постепенного снижения численности, начавшегося в 2001 г.[19] Это сопровождалось аналогичным снижением вторичной продуктивности и в настоящее время является источником большого количества исследований. Для объяснения POD был предложен ряд гипотез, включая сокращение пищевой сети, экспорт воды из Дельты и токсичные вещества из городских, промышленных или сельскохозяйственных источников.
Продюсеры
Первичная продукция и потребление питательных веществ
Основное производство фитопланктоном фиксирует энергию и ключевые питательные вещества в биологически доступную форму (то есть в пищу) посредством фотосинтез. Продукция фитопланктона во многом определяется физическими параметрами: доступностью питательных веществ, солнечным светом, мутностью и температурой.
В устье Сан-Франциско есть многочисленные источники питательных веществ, которые могут быть использованы для первичного производства, в основном из очистных сооружений, сельскохозяйственных и городских сточных вод, а также океана.[20][21] Несмотря на это, эстуарий уникален тем, что имеет тенденцию к относительно низкой скорости первичной продукции.[22] Вероятно, это связано с двумя факторами: большим поступлением азота в виде аммоний, который подавляет поглощение нитратов фитопланктоном, который предпочитает метаболически более дешевый NH4+ и высокая мутность, которая ограничивает свет, необходимый для фотосинтеза, до верхних нескольких сантиметров водяного столба.[21] Эта мутность - наследие гидравлической добычи золота в Сьерра-Невада в 1850-х гг.[23]
Большое время пребывания воды в устье имеет тенденцию к накоплению биомассы фитопланктона, увеличивая плотность, в то время как малое время пребывания удаляет фитопланктон из устья.[1] Последнее типично для основных каналов эстуария в периоды сильного стока, когда поверхностные воды имеют тенденцию смывать частицы и планктон вниз по течению.
Травоядные также удаляет фитопланктон из водной толщи. В то время как пелагическая трофическая сеть основана на продукции фитопланктона, большая часть этой продукции перенаправляется в бентос в результате хищничества интродуцированных моллюсков реки Амур (Corbula amurensis ). Уровни биомассы фитопланктона снизились на порядок после повсеместной интродукции C. amurensis в середине 1980-х годов и не восстановились.[24]
Фотосинтетическая продукция
Основной источник фотосинтетически производная энергия фитопланктон. Вообще говоря, диатомеи и микрофлагелляты производят большую часть биодоступный углерод в устье.[25] Другие типы, особенно динофлагелляты, может производить вредоносное цветение водорослей, или же красные приливы, которые менее доступны для ассимиляции в пищевой сети.[26]
Основное производство от фитопланктона зависит от двух различных факторов: скорости роста и накопления (рис. 1). Хотя LSZ является приемником высоких концентраций питательные вещества из городских и сельскохозяйственных источников темпы производства фитопланктона довольно низкие.[27] Нитрат оптимально используется фитопланктоном для роста, но аммоний (в основном происходит из сбросов сточных вод) оказывает подавляющее влияние на скорость роста. Таким образом, фитопланктон, хотя и не ограничен питательными веществами, имеет тенденцию к более медленному росту из-за присутствующих видов азота.[28] Еще одним фактором, сдерживающим рост, является высокий мутность эстуария, что ограничивает возможность фотосинтетически активная радиация (PAR), чтобы проникать за пределы верхних нескольких сантиметров водяного столба. Это ограничивает фотосинтез фитопланктона относительно мелкими участками. световая зона. Таким образом, когда столб воды является стратифицированный, мутность высокая и присутствует аммоний, скорость роста фитопланктона обычно подавляется.[29][30]
Накопление фитопланктона в первую очередь является результатом Время жительства.[27] Северная Дельта и Suisun Bay имеют относительно низкое время пребывания из-за большого объема воды, проходящей через область для нисходящего потока и для экспорт в южную Калифорнию. Поскольку вода движется через эту часть системы быстрее, скорость накопления уменьшается по мере того, как производительность выносится из системы. Напротив, части южной дельты имеют более высокое время пребывания из-за небольшого объема воды, проходящей через систему; фактически вода иногда течет в обратном направлении из-за отсутствия притока из реки Сан-Хоакин и откачки на экспорт. Летом плотность фитопланктона здесь может быть на порядок выше, чем в других частях эстуария.[31]
Вредное цветение водорослей (ВЦВ) динофлагеллят или цианобактерии производят токсичные побочные продукты метаболизма, которые делают их вредными для многих организмов. Благодаря сочетанию высоких концентраций питательных веществ и температур, ВЦВ оказывают двойное негативное влияние на пищевую сеть, исключая на конкурентной основе диатомовые водоросли и микрофлагелляты, что еще больше снижает первичную биодоступную продукцию. Хотя определенные беспозвоночные Такие как двустворчатые моллюски не могут быть затронуты напрямую, они могут распространять токсины по пищевой цепочке, вызывая тошноту или смерть хищники. Не совсем понятно, как копеподы под действием. Инвазивный водоросли Microcystis aeruginosa в настоящее время обычное дело в Дельте в летние месяцы и может снизить продуктивность копепод (помимо того, что потенциально канцерогенный для человека).[32]
Детритовая добыча
Огромное количество осадок и детрит поток через LSZ. По большей части это органический мусор в виде растворенных частиц и частиц. органическая материя (DOM и POM соответственно). Помимо источников, расположенных выше по течению, органическое вещество может накапливаться в результате гибели местных организмов и образования отходов.[33][34]
Детритофаги извлекать выгоду из этого источника энергии, создавая альтернативную и параллельную пищевую сеть потенциально большого значения. Это потому что фиксация углерода попадание в детритную пищевую сеть не ограничивается стратификацией, мутностью или продолжительностью дня, которые ограничивают фотосинтез. Добыча обломков происходит непрерывно, ограничиваясь только поступлениями и адвекцией из системы Дельта.[25][35]
Бактерии - главные агенты преобразования РОВ и ПОМ в биодоступный углерод через микробная петля. Этот механизм особенно важен при ограничении питательных веществ. морской системы, где бактерии высвобождают питательные вещества из тонущего детрита, позволяя вернуть его обратно в фотическую зону. В отношении функции микробной петли в устье Сан-Франциско было проведено мало исследований, но, возможно, роль бактерий не является критической для повторного использования питательных веществ в эвтрофный система. Скорее, они могут обеспечить альтернативную пищевую цепочку за счет прямого выпаса скота жгутиконосцы, коловратки и инфузории.[36]
Высокая численность циклопоид копепода Лимноитона тетраспина может быть связано с тем, что он полагается на инфузории, а не на фитопланктон в качестве основного источника пищи.[37] Основные виды каланоид Копеподы могут также использовать инфузорий в качестве дополнительного или даже основного источника пищи, но в какой степени неизвестно.[38][39]
Вторичное производство
Вторичное производство относится к организмам, которые питаются первичным производством и передают энергию более высоким трофические уровни эстуарной пищевой сети. Исторически сложилось так, что вторичное производство в устье Сан-Франциско определялось креветка-мизид производство.[40][41] Однако родной mysid Неомизис мерседис был в значительной степени заменен введенным Acanthomysis Bowmani, который сохраняется при более низких плотностях. Представленный амфипода Гаммурус дайбери возможно, занял часть этой ниши, но в основном это пресная вода.
Сегодня основным источником вторичного производства является копеподы. Натурализованный уроженец каланоид копепода Eurytemora affinis Считается, что он был завезен в конце 19 века.[42] Он доминировал в зоопланктоне зоны низкой солености до 1980-х годов, когда он был в значительной степени заменен другим завезенным каланоидным копеподом, Pseudodiaptomus forbesi.[16][43] P. forbesi сохраняется за счет сохранения исходной популяции в пресноводных, густонаселенных районах устья, особенно в дельте, за пределами допустимого диапазона солености моллюска реки Амур.[44] Поскольку некогда доминирующий E. affinis не имеет ареала вверх по течению, он более уязвим для хищников со стороны моллюсков и страдает от явной конкуренции с P. forbesi.
Другими каланоидными веслоногими рачками, которые могут иметь значение, являются недавно введенные Sinocalanus doerri и Acartiella sinensis. Мало что известно об истории жизни этих организмов, хотя, исходя из их морфологии, они могут охотиться на других веслоногих ракообразных. Они появляются в нерегулярных циклах численности, во время которых могут доминировать в зоопланктоне.[45]
Еще один инвазивный веслоногий рачок, очень маленький циклопоид Лимноитона тетраспина, появился в зоне низкой солености в 1990-х гг. С тех пор L. tetraspina стала численно доминирующей копеподой, достигнув плотности порядка 10 000 / м3. Он полагается на микробная петля в качестве источника пищи, питаясь бактериями, инфузориями и коловратками.[37] Кроме того, он кажется неуязвимым для хищников амурского моллюска по неизвестным причинам. Из-за своего небольшого размера L. tetraspina обычно недоступен для употребления более крупными хищниками, особенно рыбами, что делает его энергетическим тупиком.
Потребители
Основные потребители
Основные потребители полагаться на основное производство как основной источник пищи. Наиболее важными потребителями пелагической сети LSZ являются веслоногие рачки, а также упомянутые выше коловратки, жгутиконосцы и инфузории. Все виды каланоидных веслоногих моллюсков сократились под высоким давлением хищников недавно завезенного моллюска реки Амур (Corbula amurensis).[16] Из-за этого, а также из-за того, что веслоногие ракообразные полагаются как на фотосинтетические, так и на детритные источники пищи, веслоногие рачки в LSZ имеют ограниченную обратную связь по первичной продукции, в отличие от морских и лентичных систем, где веслоногие рачки могут пастись на цветках за считанные дни.
Псевдодиаптомус форбези является доминирующим каланоидным копеподом LSZ по биомассе. Он имеет достаточно широкий диапазон солености, поэтому может сохраняться как при низкой солености, так и в пресной воде. Такое широкое распространение помогает популяции сохранять убежище от хищников вверх по течению, в отличие от других видов с более узкими допусками к засолению.[44]
Лимноитона тетраспина стал численно доминирующим циклопоидным веслоногим с момента его появления в 1993 году. Он питается в основном инфузориями и микрофлагеллятами, но в отличие от P. forbesi, он относительно невосприимчив к хищникам со стороны моллюсков или рыб, отсюда и его многочисленность. Энергетически, L. tetraspina может оказаться тупиком для пищевой сети; эти веслоногие рачки либо выводятся из системы приливами и течениями, либо умирают и падают в бентос, где они могут быть доступны микробной петле или детритивным животным.[46]
Хищные веслоногие рачки
По всей Дельте существует ряд хищных веслоногих рачков, о которых известно относительно немного. Sinocalanus doerri, Acartiella sinensis, и Tortanus dextrilobatus все они морфологически способны нападать на других веслоногих рачков. Каждый был введен в устье, вероятно, через балластная вода биржа с 1980-х гг. Как правило, их недостаточно, чтобы отрицательно повлиять на потребителей веслоногих рачков; однако периодические цветения С. Доерри и A. sinensis происходят, которые не были хорошо изучены.[47]
Макробеспозвоночные

Хотя способен фильтрующий, мизиды (крошечные креветочные существа) в основном плотоядны, питаясь взрослыми веслоногими рачками. Они обеспечивали энергетический канал между планктоном и планктоноядными рыбами, включая молодь рыб, осетр, Чавычи лосось, и Американский шэд. Когда-то мизиды были в изобилии, пока Неомизис мерседис был заменен в середине 1980-х годов инвазивным Acanthomysis Bowmani, который меньше и менее многочисленен. Снижение мизид было связано с последующим сокращением численности ряда видов рыб в эстуарии в 1980-х и 90-х годах.[48][49]
Креветка хищники широкого профиля, которые охотятся в основном на мизидов и амфиподы. Crangon franciscorum представляет собой один из двух оставшихся коммерческих промыслов в устье. Хотя они больше не используются для приготовления коктейлей из креветок в заливе Сан-Франциско, их собирают для наживки. Другие хищники включают полосатый окунь и Чавычи лосось взрослые и смолты.[50]
Рыбы

Потому что рыба - это таксономически и морфологически разнообразная группа, виды различаются по своему трофический экология. В целом, рыбу можно разделить на четыре основные категории кормления: питатели-фильтры, планктоядные, рыбоядные и бентосный кормушки.
Фильтр-питатели без разбора нагружают толщу воды мелкой добычей, как правило, фито- и зоопланктоном. В эту категорию рыб входят шэд (Доросома петененсе), Американский шэд (Алоза сапидиссима), внутренние серебряные берега (Менидия бериллина), и анчоусы (Engraulis mordax). Некоторые данные свидетельствуют о том, что некоторые из этих видов ограничены в пище из-за пониженного уровня планктона после интродукции моллюска из реки Амур. Анчоусы покинули LSZ в пользу более продуктивных регионов устья в Сан-Пабло и Центральных заливах.[17]
Планкоядные выборочно охотятся на отдельный зоопланктон, такой как веслоногие рачки, мизиды и гаммариды. В эту группу входит большинство личинок рыб, дельта корюшка (Гипомез транспацифический ) и длинноперый корюшка (Spirinchus thaleichthys ), тюльский окунь (Hysterocarpus traski), и лосось смолты. Дельта корюшка представляет особый интерес из-за своего статус под угрозой исчезновения. Это может быть ограниченное питание, но доказательства несколько противоречивы. Другие факторы, такие как унос икры и личинок при выкачивании пресной воды из дельты на экспорт, также могут объяснить снижение.[11]

Основное животное LSZ - это полосатый окунь (Morone saxatilis ), который был завезен в 1890-х годах и сильно охотится на местных рыб. Полосатый окунь является важным спортивным промыслом в устье Сан-Франциско и, как таковой, представляет собой незначительное изъятие биомассы из устья.[51]
Бентические или донные рыбы включают: белый осетр (Acipenser transmontanus), белый сом (Ameiurus catus), и звездная камбала (Platichthys stellatus). Из-за их среда обитания Ориентация, они питаются в основном эпибентосными организмами, такими как амфиподы, заливные креветки и двустворчатые моллюски. Известно, что эти рыбы, по крайней мере, иногда кормятся моллюском реки Амур, который представляет собой один из немногих каналов для потока энергии от этого вида, за исключением производства обломков.[52]
Единственным коммерческое рыболовство в LSZ для креветок наживки. Есть множество спортивное рыболовство которые представляют собой незначительный поток углерода, но значительные потоки капитала в местные экономики вокруг устья. Большинство рекреационных промыслов занимаются полосатым окунем, осетровыми и введены пресноводные бассейны в пресноводной дельте. Такая нехватка рыбных ресурсов делает устье Сан-Франциско уникальным. Почти все эстуарии во всем мире поддерживают по крайней мере остатки значительного рыболовства.[53] Устье Сан-Франциско одно время поддерживало крупный промысел лосося, анчоусов и Крабы Dungeness до 1950-х гг. Исчезновение этих промыслов, вероятно, было связано с потеря среды обитания чем чрезмерный сбор.[2]
Птицы
Устье Сан-Франциско - главная остановка на Тихоокеанский пролетный путь для перелета водоплавающих птиц. Однако мало что известно о потоке углерода в устье или из устья через птицы. Миллионы водоплавающая птица ежегодно используют отмели залива как убежище. Большинство птиц веселящиеся утки которые питаются подводной водной растительностью. Ныряющие утки (например, scaups ) питаются эпибентосными организмами, такими как C. amurensis, представляя возможный поток углерода из этого тупика.[54][55] Два вымирающие виды птиц здесь водятся: Калифорнийская наименее крачка и Калифорнийская задвижка. Незащищенный заливные грязи обеспечить важные зоны кормления для кулики, но нижележащие слои ила залива представляют геологическую опасность для структур, расположенных вблизи многих частей периметра залива. Рыбоядные птицы, такие как бакланы и пеликаны также обитают в эстуарии, но их трофическое влияние остается малоизученным.
В январе 2015 года ученые работали над выявлением серого, густого, липкого покрытия без запаха на птицах вдоль береговой линии залива Сан-Франциско. Сотни птиц погибли, и еще сотни были покрыты веществом. Ученых беспокоят другие дикие животные, которым это вещество может угрожать.[56]
Млекопитающие


До 1825 года испанцы, французы, англичане, русские и американцы были привлечены в Район залива для сбора огромного количества урожая. бобр (Castor canadensis), речная выдра, куница, рыболов, норка, лисица, ласка, гавань и морские котики (Каллоринус урсинус) и морская выдра (Enhydra lutris). Эта ранняя торговля мехом, известная как Калифорнийская меховая гонка, более чем какой-либо другой фактор, ответственный за открытие Запада и Область залива Сан-Франциско, в частности, мировой торговле.[57] К 1817 году каланы в этом районе были практически истреблены.[58] Русские сохранили станцию по опечатыванию поблизости Фараллонские острова с 1812 по 1840 год, ежегодно забирая от 1200 до 1500 морских котиков, хотя американские корабли уже эксплуатировали острова.[59] К 1818 году количество тюленей быстро уменьшилось, и в год можно было вылавливать только около 500 особей, а в течение следующих нескольких лет морской котик был истреблен с островов, пока в 1996 году они не начали переселять острова.[60] Хотя натуралисты 20-го века скептически относились к тому, что бобры исторически существовали в прибрежных ручьях или в самом заливе,[61] более ранние записи показывают, что калифорнийский золотой бобер (Castor canadensis ssp. субауратус) был одним из самых ценных из взятых животных и, по-видимому, был найден в большом количестве.[57] Томас Маккей сообщил, что за один год компания Гудзонова залива забрала 4000 бобровых шкур на берегах залива Сан-Франциско. Недавно бобры заселили солоноватую Напа Сонома Марш на севере Сан-Пабло Бэй и это Sonoma Creek и Река Напа притоки. Также недавно реколонизируя залив и его притоки, Североамериканская речная выдра (Lontra canadensis) впервые было сообщено в Redwood Creek в Muir Beach в 1996 г.[62] и недавно в Corte Madera Creek, а в Южном заливе на Койот Крик,[63] а также в 2010 году в самом заливе Сан-Франциско на Ричмонд Марина.
Медуза
Медуза до недавнего времени не были распространены в устье. В устьях Восточного побережья, таких как Chesapeake залив, они часто хищники высшего уровня, без разбора питаясь как рыбой, так и зоопланктоном. Несколько небольших инвазивных таксонов были идентифицированы в LSZ и пресноводных регионах. Эти виды стробилируют летом, но сохраняют полипы в бентос весь год.
Их влияние на планктон неизвестно, но в настоящее время проводятся исследования для его количественной оценки. При достаточной плотности желе могут играть дополнительную роль по отношению к C. amurensis в подавлении зоопланктона, населяя районы с низкой соленостью за пределами ареала моллюсков, где планктонные виды имели убежище без хищников.
Бентические потребители
В бентосный Сообщество взяло на себя непропорционально большую роль в экологии пищевой сети эстуария из-за основных вторжений двустворчатые моллюски.[16] Использование этих моллюсков, моллюск реки Амур (Corbula amurensis) имеет широкий допуск по солености, который распространяется на зону с низкой соленостью, но не в пресную воду. Фильтр питается фитопланктоном и мелким зоопланктоном, например каланоидными копеподами. науплии. У моллюсков мало хищников в устье Сан-Франциско, и это позволяет им расти до высокой плотности (порядка десятков тысяч / м2). Благодаря высокой скорости очистки, он способен очистить всю толщу воды от частей эстуария за несколько дней, что приводит к резкому истощению популяций планктона.[64] Считается, что это основная причина снижения продуктивности экосистем после нашествия моллюсков в середине 1980-х годов.[16][65][66]
Это снижение продуктивности в основном связано с перенаправлением пелагической сети на бентическую цепь этим одним видом.[16] Поскольку моллюск реки Амур питается первичными производителями, потребителями и хищниками, он оказывает влияние на несколько трофические уровни. Следовательно, почти весь планктон проявляет признаки явная конкуренция, поскольку продукция на одном трофическом уровне влияет на все остальные, увеличивая численность моллюсков. Это приводит к отрицательному Обратная связь: C. amurensis ограничивает биомассу планктона, что, в свою очередь, ограничивает C. amurensis. Однако поступление извне системы из-за приливной адвекции или источников вверх по течению может увеличиваться. C. amurensis биомасса, дальнейшее ограничение планктона. Эта обратная связь еще больше усиливается, поскольку моллюск может сохраняться более одного или двух лет, что оказывает дополнительное давление на популяции планктона во время циклов низкой продуктивности.
Перенаправление углерода C. amurensis в бентос создало ограниченную цепочку, в результате чего пелагическая сеть стала разряженной. Производство детрита в результате экскреции и гибели моллюсков может стимулировать производство бактерий, которые могут циркулировать в детритной пищевой сети или микробной петле.
В то время как переработанные питательные вещества могут поддерживать некоторый рост фитопланктона, в конечном итоге они подпитываются увеличением популяций C. amurensis. Недавнее успешное вторжение Limnoithona tetraspina можно понять в терминах этого феномена. Он питается инфузории и микрофлагелляты, которые слишком малы, чтобы их мог поедать моллюск, что позволяет избежать конкуренции. Кроме того, L. tetraspina кажется невосприимчивым к хищничеству моллюсков или (почти) чего-либо еще. Рост микробной пищевой сети и вторжение L. tetraspina capitalize являются результатом неиспользованного альтернативного пути потока энергии в пищевой сети, которому способствует C. amurensis. Последующие модели вторжения могут отражать аналогичную модель.
Интродуцированные виды
Интродукция видов увеличивалась, по крайней мере, с 19 века как функция увеличения торговли и движения. Интродукции включают многочисленные таксоны, в том числе веслоногие рачки, креветка, амфиподы, двустворчатые моллюски, рыба, а также корневые и плавающие растения. Многие пелагические виды были интродуцированы совсем недавно через балластная вода выпуски с больших кораблей прямо в лиман.[67] В результате многие из этих интродуцированных видов происходят из устьев Тихоокеанского побережья, особенно веслоногие рачки, такие как P. forbesi и L. tetraspina. Моллюски реки Амур происходят из Азии и привели к значительным и радикальным изменениям в экологии ЗЗЗ, в первую очередь за счет перенаправления пелагической пищи в бентос и в ускоренную микробную петлю.[68]
Виды также были завезены посредством прикрепления к спортивным лодкам, которые курсируют между регионами.[67] Это вероятный источник ряда растений с низкой соленостью, таких как Эгерия денса и водяной гиацинт (Эйхорния крассипес ). Эти растения вызвали глубокие изменения в Дельте, нарушив поток воды, затеняя фитопланктон и создавая среду обитания для рыбоядный рыба как полосатый окунь, Morone saxatilis, сама намеренно введенная в конце 1800-х годов из Chesapeake залив.[51][67] Пресная вода квагга мидия, родом из Европы, ожидается, что яхтсмены будут завезены сюда в течение следующих двух-десяти лет, несмотря на принятые меры предосторожности.[69] Более того, молодой возраст и изолированная среда устья Сан-Франциско привели к успеху инвазивных беспозвоночных из-за отсутствия биоразнообразия.[53]
Будущие вторжения
Современная трофическая сеть образовалась в результате серии вторжений и трофических замен.
Ожидается, что этот процесс будет продолжаться по мере поступления новых организмов в результате случайной или преднамеренной интродукции. Менее ясно то, в какой степени предыдущие внедрения прокладывают путь для будущих вторжений. Это может произойти одним из трех способов.
- Ранний захватчик может предоставить ресурс, который не используется в новой системе до тех пор, пока не появится новый хищник (L. tetraspina and the microbial loop, as described above).
- Early invaders may facilitate new ones by altering habitat and making it suitable for subsequent invasions (jelly polyps using Amur River clam shells for substrate).
- Apparent competition between old and new residents may increase the possibilities for invasion and settlement of new organisms that can capitalize on unexploited resources (the subsidization of the Amur River clam by upstream populations of the introduced copepod P. forbesi, creating pressure on native copepods).
Резюме
The LSZ food web of the San Francisco Estuary operates in two parallel and asymmetrical directions. The bulk of carbon is assimilated into the benthic and microbial loops, which represent energetic dead ends. A smaller fraction is delivered to higher pelagic trophic levels which may support copepods, fish, birds and fisheries. This redirection of the food web into these two narrow loops may be responsible for the decline in macroinvertebrates and fishes in the estuary, which operate outside of these chains. Restoration of the estuary to a higher degree of function relies upon the probability of delivering increased benefits to the pelagic web without subsidizing the benthic.
Future ecology
The ecology of the Low Salinity Zone of the San Francisco Estuary is difficult to characterize because it is the result of a complex synergy of both abiotic and biotic factors. In addition, it continues to undergo rapid change resulting from newly introduced species, direct anthropogenic influences and изменение климата. Future ecological changes will be driven on an ecosystem wide scale, particularly as повышение уровня моря, тектонический instability and infrastructure decline cause levee failure in the Delta.[70] The resulting back-surge in water flow is expected to force X2 into the Delta, jeopardizing spatially oriented habitat (like freshwater marshes), channelizing the low salinity zone, and threatening southern California's water supply, with unknown and unforeseeable consequences for the natural and human ecology of the West coast's largest estuary.
Смотрите также
- Список водотоков в районе залива Сан-Франциско
- Гидрография района залива Сан-Франциско
- Проект водораздела
- Наука об устье и водоразделе Сан-Франциско
Примечания
- ^ а б c d е Kimmerer 2004
- ^ а б Conomos 1979
- ^ Vayssieres 2006
- ^ Jassby 2000
- ^ Deverel 1998
- ^ Philip 2007
- ^ Lund 2007
- ^ а б Kimmerer 2002
- ^ Gunther, Andrew (2011). "The State of the San Francisco Bay 2011, San Francisco Estuary Partnership" (PDF). The State of the San Francisco Bay 2011. Получено 6 октября 2017.
- ^ Moyle 1992
- ^ а б Bennett 2006
- ^ Monismith 1996
- ^ Burau 1998
- ^ Schoellhamer 2001
- ^ Orsi 1986
- ^ а б c d е ж Kimmerer 1996
- ^ а б Kimmerer 2006
- ^ Davis, Jay (September 2006). "The Pulse of the Estuary 2006" (PDF). sfei.org. Институт устья Сан-Франциско. Получено 2 ноября 2016.
- ^ Taugher 2005
- ^ Смит 2002
- ^ а б Dugdale 2003
- ^ Jassby 2002
- ^ Nichols 1986
- ^ Kimmerer 1994
- ^ а б Jassby and Cloern 2000
- ^ Anderson et al. 2002 г.
- ^ а б Jassby et al. 2002 г.
- ^ Dugdale et al. 2003 г.
- ^ Cloern 1987
- ^ Cole and Cloern 1987
- ^ Ball and Arthur 1979
- ^ Lehman and Waller 2003
- ^ Murrell et al. 1999 г.
- ^ Murrell and Hollibaugh 2000
- ^ Jassby et al. 1993 г.
- ^ Hollibaugh and Wong 1996
- ^ а б Bouley 2006
- ^ Rollwagen-Bollens and Penry 2003
- ^ Rollwagen-Bollens et al. 2006 г.
- ^ Modlin 1997
- ^ Kimmerer 1998
- ^ Lee 1999
- ^ Orsi 1991
- ^ а б Durand2006
- ^ Orsi 1999
- ^ Bouley and Kimmerer 2006
- ^ Orsi and Ohtsuka 1999
- ^ Orsi and Knutson 1979
- ^ Modlin and Orsi 1997
- ^ Sitts and Knight 1979
- ^ а б Radovich 1963
- ^ Peterson 1997
- ^ а б Nichols et al. 1986 г.
- ^ Poulton et al. 2002 г.
- ^ Richman and Lovvorn 2004
- ^ Scientists race to identify goop on birds along San Francisco Bay shorelines (Январь 2015), Christian Science Monitor
- ^ а б Skinner, John E. (1962). An Historical Review of the Fish and Wildlife Resources of the San Francisco Bay Area (The Mammalian Resources). California Department of Fish and Game, Water Projects Branch Report no. 1. Sacramento, California: California Department of Fish and Game. Архивировано из оригинал на 2011-07-26.
- ^ Suzanne Stewart and Adrian Praetzellis (November 2003). Archeological Research Issues for the Point Reyes National Seashore - Golden Gate National Recreation Area (PDF) (Отчет). Anthropological Studies Center, Sonoma State University. п. 335. Получено 10 января, 2010.
- ^ Thompson, R. A. (1896). The Russian Settlement in California Known as Fort Ross, Founded 1812...Abandoned 1841: Why They Came and Why They Left. Santa Rosa, California: Sonoma Democrat Publishing Company. п. 7. ISBN 0-559-89342-6.
- ^ White, Peter (1995). The Farallon Islands: Sentinels of the Golden Gate. San Francisco, California: Scottwall Associates. ISBN 0-942087-10-0.
- ^ Таппе, Дональд Т. (1942). «Статус бобров в Калифорнии». Игровой бюллетень № 3. Калифорнийский департамент рыбы и дичи.
- ^ "Get Outside!". Хроники Сан-Франциско. Апрель 1966 г.. Получено 2011-03-06.
- ^ "Blue Oak Ranch Reserve". Калифорнийский университет. Архивировано из оригинал на 2011-05-12. Получено 2011-03-06.
- ^ Werner and Hollibaugh 1993
- ^ Carlton et al. 1990 г.
- ^ Nichols et al. 1990 г.
- ^ а б c Carlton 1996
- ^ Коул 1992
- ^ Dey 2007
- ^ Epstein 2006
Рекомендации
- Anderson, D. M., P. M. Glibert, and J. M. Burkholder. 2002. Harmful Algal Blooms and Eutrophication: Nutrient Sources, Composition, and Consequences. Estuaries 25:704-726.
- Ball, M. D., and J. F. Arthur. 1979. Planktonic chlorophyll dynamics in the northern San Francisco Bay and Delta. Pages 265–286 in T. J. Conomos, editor. San Francisco Bay: The urbanized estuary. American Association for the Advancement of Science, Pacific Division, San Francisco, CA.
- Bennett, W. A. (2006). Delta Smelt Growth and Survival During the Recent Pelagic Organism Decline: What Causes Them Summer Time Blues? CALFED Science Conference. Сакраменто, Калифорния.
- Bouley P., W. J. K. (2006). "Ecology of a highly abundant, introduced cyclopoid copepod in a temperate estuary. " Marine Ecology Progress Series 324: 219–228.
- Burau, J. R. (1998). "Results from the hydrodynamic element of the 1994 entrapment zone study in Suisun Bay, Sacramento, CA. " Interagency Ecological Program for the San Francisco Bay-Delta Estuary Technical Report 56: 13–62.
- Carlton, J. T., J. K. Thompson, L. E. Schemel, and F. H. Nichols. 1990. Remarkable invasion of San Francisco Bay (California, USA) by the Asian clam Potamocorbula amurensis. 1. Introduction and dispersal. Серия «Прогресс морской экологии». 66:81-94.
- Carlton, J. (1996). "Biological Invasions and Cryptogenic Species. " Ecology 77(6).
- Cole, B. E., and J. E. Cloern. 1987. An empirical model for estimating phytoplankton productivity in estuaries. Серия «Прогресс морской экологии». 36:299-305.
- Cole, B. E., J. K. Thompson, and J. E. Cloern (1992). "Measurement of filtration rates by infaunal bivalves in a recirculating flume. " Marine Biology 113: 219–225.
- Cloern, J. E. 1987. Turbidity as a control on phytoplankton biomass and productivity in estuaries. Continental Shelf Research 7:1367-1381.
- Conomos, T. J., Ed. (1979). San Francisco Bay: The urbanized estuary. San Francisco, CA, Pacific Division, American Association for the Advancement of Science.
- Deverel, S. (1998). Subsidence mitigation in the Sacramento-San Joaquin Delta., CALFED Bay-Delta Program.
- Dey, R. (2007, January 16, 2007). "Invasive Mussel Update from Lake Mead National Recreation Area. " 100th Meridian Initiative, from http://100thmeridian[постоянная мертвая ссылка ]. org/mead. asp.
- Dugdale, R. C., V. Hogue, A. Marchi, A. Lassiter, and F. Wilkerson. 2003. Effects of anthropogenic ammonium input and flow on primary production in the San Francisco Bay. CALFED Science Program, Sacramento, CA.
- Durand, J. (2006). Determinants of calanoid copepod recruitment failure in the San Francisco Estuary Calfed Science Program. Сакраменто, Калифорния.
- Epstein, E. (2006). Creaky levees ripe for disaster. Хроники Сан-Франциско. Сан-Франциско.
- Hollibaugh, J. T., and P. S. Wong. 1996. Distribution and activity of bacterioplankton in San Francisco Bay. Pages 263–288 in J. T. Hollibaugh, editor. San Francisco Bay: The ecosystem. American Association for the Advancement of Science, San Francisco, CA.
- Jassby, A. D., J. E. Cloern, and B. E. Cole. 2002. Annual primary production: Patterns and mechanisms of change in a nutrient-rich tidal ecosystem. Limnology and Oceanography 47:698-712.
- Jassby, A. D., J. E. Cloern, and T. M. Powell. 1993. Organic carbon sources and sinks in San Francisco Bay - variability induced by river flow. Серия «Прогресс морской экологии». 95.
- Jassby, A. D., and J. E. Cloern. 2000. Organic matter sources and rehabilitation of the Sacramento-San Joaquin Delta (California, USA). Aquatic Conservation: Marine and Freshwater Ecosystems 10:323-352.
- Jassby, A. D., J. E. Cloern and B. E. Cole. (2002). "Annual primary production: Patterns and mechanisms of change in a nutrient-rich tidal ecosystem. " Limnology and Oceanography 47(3): 698–712.
- Kimmerer, W. J., E. Gartside, and J. J. Orsi. (1994). "Predation by an introduced clam as the likely cause of substantial declines in zooplankton of San Francisco Bay. " Mar. Ecol. Прог. Сер. 113: 81–93.
- Kimmerer, W. J., J. J. Orsi. 1996. Changes in the zooplankton of the San Francisco Bay estuary since the introduction of the clam Potamocorbula amurensis. Pages 403–425 in J. T. Hollibaugh, editor. San Francisco Bay: The Ecosystem. American Association for the Advancement of Science, San Francisco, CA.
- Kimmerer, W. J., J. R. Burau, and W. A. Bennett. (1998). "Tidally oriented vertical migration and position maintenance of zooplankton in a temperate estuary. " Limnol. Oceanogr. 43(7): 1697–1709.
- Kimmerer, W. J. (2002). "Physical, biological, and management responses to variable freshwater flow into the San Francisco Estuary. " Estuaries 25: 1275–1290.
- Kimmerer, W. (2004). "Open water process of the San Francisco Estuary: from physical forcing to biological responses. " San Francisco Estuary and Watershed Science 2(1).
- Kimmerer, W. (2006). "Response of anchovies dampens effects invasive bivalve Corbula amurensis on San Francisco Estuary foodweb. " Marine Ecology Progress Series 324: 207–218.
- Lee, C. E. (1999). "Rapid and repeated invasions of fresh water by the copepod Eurytermora affinis. " Evolution 53(5): 1423–1434.
- Lehman, P. W., and S. Waller. 2003. Microcystis blooms in the delta. IEP Newsletter 16:18-19.
- Lund, J., E. Hanak, W. Fleenor, R. Howitt, J. Mount, P. Moyle (2007). Envisioning Futures for the Sacramento-San Joaquin Delta. San Francisco, Public Policy Institute.
- Modlin, R. F., and J. J. Orsi (1997). "Acanthomysis bowmani, a new species, and A. asperali, Mysidacea newly reported from the Sacramento-San Joaquin Estuary, California (Crustacea: Mysidae). " Proceedings of the Biological Society of Washington 110(3): 439–446.
- Monismith, S. G., J. R. Burau, and M. Stacey (1996). Stratification dynamics and gravitational circulation on northern San Francisco Bay. San Francisco Bay: The ecosystem. J. T. Hollibaugh. San Francisco, CA, American Association for the Advancement of Science: 123–153.
- Moyle, P. B., B. Herbold, D. E. Stevens, and L. W. Miller. (1992). "Life history and status of delta smelt in the Sacramento-San Joaquin Estuary, California. " Trans. Являюсь. Рыбы. Soc. 121: 67–77.
- Murrell, M. C., J. T. Hollibaugh, M. W. Silver, and P. S. Wong. 1999. Bacterioplankton dynamics in northern San Francisco Bay: Role of particle association and seasonal freshwater flow. Limnology and Oceanography. 44:295-308.
- Murrell, M. C., and J. T. Hollibaugh. 2000. Distribution and composition of dissolved and particulate organic carbon in northern San Francisco Bay during low flow conditions. Estuarine, Coastal and Shelf Science 51:75-90.
- Nichols, F. H., J. E. Cloern, S. Luoma, and D. H. Peterson. 1986. The modification of an estuary. Science 231:567-573.
- Nichols, F. H., J. K. Thompson, and L. E. Schemel. 1990. Remarkable invasion of San Francisco Bay (California, USA) by the Asian clam Potamocorbula amurensis. 2. Displacement of a former community. Серия «Прогресс морской экологии». 66:95-101.
- Orsi, J. J., and A. C. Knutson. 1979. The role of mysid shrimp in the Sacramento-San Joaquin Estuary and factors affecting their abundance and distribution. Pages 401–408 in T. J. Conomos, editor. San Francisco Bay: the urbanized estuary. American Association for the Advancement of Science, Pacific Division, San Francisco.
- Orsi, J. J., and W. L. Mecum. (1986). "Zooplankton distribution and abundance in the Sacramento-San Joaquin Delta in relation to certain environmental factors. " Estuaries 9(4 B): 326–339.
- Orsi, J. J., and T. C. Walter. (1991). "Pseudodiaptomus forbesi and P. marinus (Copepoda: Calanoida), the latest copepod immigrants to California's Sacramento-San Joaquin Estuary. " Bull. Plankton Soc. Japan Spec. Vol. (Proceedings of the Fourth International Conference on Copepoda): 553–562.
- Orsi, J. J., and S. Ohtsuka. (1999). "Introduction of the Asian copepods Acartiella sinensis, Tortanus dextrilobatus (Copepoda: Calanoida), and Limnoithona tetraspina (Copepoda: Cyclopoida) to the San Francisco Estuary, California, USA. " Plank. Bio. Ecol. 46: 128–131.
- Peterson, H. 1997. Clam-stuffed sturgeon. IEP Newsletter 9:19-19.
- Philip, T. (2007). A time to deliver water solutions. Сакраменто пчела. Сакраменто, Калифорния.
- Poulton, V. K., J. R. Lovvorn, and J. Y. Takekawa. 2002. Clam density and scaup feeding behavior in San Pablo Bay, California. Кондор. 104:518-527.
- Radovich, J. (1963). "Effects of ocean temperature on the seaward movement of striped bass, Roccus saxatilis, on the Pacific coast. " California Department of Fish and Game 49: 191–205.
- Richman, S. E., and J. R. Lovvorn. 2004. Relative foraging value to lesser scaup ducks of native and exotic clams from San Francisco Bay. Ecological Applications 14:1217-1231.
- Rollwagen-Bollens, G. C., and D. L. Penry. 2003. Feeding dynamics of Acartia sp. in a large, temperate estuary (San Francisco Bay, CA). Marine Ecology Progress Series 257:139-158.
- Rollwagen-Bollens, G., S. Gifford, A. Slaughter, and S. M. Bollens. 2006. Protists in a Temperate Estuary: Diversity, Grazing and Consumption by Metazoans. Eos, Transactions of the American Geophysical Union, Ocean Sciences Meeting Supplement. 87:36 OS11D-05.
- Schoellhamer, D. H. (2001). Influence of salinity, bottom topography, and tides on locations of estuarine turbidity maxima in northern San Francisco Bay. Coastal and Estuarine Fine SedimentProcesses. W. H. McAnally and A. J. Mehta. Amsterdam, Netherlands, Elsevier: 343–356.
- Sitts R. M., A. W. Knight. 1979. Predation by the estuarine shrimps Crangon franciscorum Stimpson and Palaemon macrodactylus Rathbun. Biological Bulletin 156:356-368.
- Smith, S. V., and J. T. Hollibaugh (2002). "Water, salt, and nutrient exchanges in the San Francisco Bay. " Interagency Ecological Program for the San Francisco Bay-Delta Estuary TechnicalReport 66.
- Taugher, Mike (2005). Environmental sirens in Delta are screaming. Contra Costa Times.
- Trott, J. S. S., Warren J. Ferguson, and Andrew J. Kleinfeld, (2006). Central Delta Water Agency v. Bureau of Reclamation. Апелляционный суд США девятого округа. Сан-Франциско, Калифорния.
- Vayssieres, M. P. (2006). Trends in the benthos of the upper San Francisco Esutary. CALFED Science Conference. Сакраменто, Калифорния.
- Werner, I., and J. T. Hollibaugh. 1993. Potamocorbula amurensis (Mollusca, Pelecyopoda): Comparison of clearance rates and assimilation efficiencies for phytoplankton and bacterioplankton. Limnology and Oceanography 38:949-964.


