Экосистема субтропического круговорота северной части Тихого океана - Ecosystem of the North Pacific Subtropical Gyre
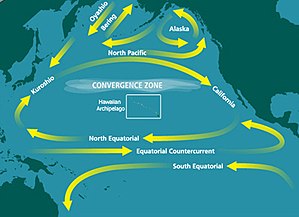
В Субтропический круговорот северной части Тихого океана (NPSG) - крупнейшая непрерывная экосистема на Земле. В океанография, а субтропический круговорот кольцевая система Океанские течения вращаясь по часовой стрелке в Северное полушарие и против часовой стрелки в Южное полушарие вызвано Эффект Кориолиса. Как правило, они образуются на больших открытых участках океана, которые лежат между массивами суши.
NPSG - крупнейший из круговоротов, а также самая большая экосистема на нашей планете. Как и у других субтропических круговоротов, в его центре находится зона высокого давления. Циркуляция вокруг центра происходит по часовой стрелке вокруг этой зоны высокого давления. Субтропические круговороты составляют 40% поверхности Земли и играют важную роль в фиксация углерода и круговорот питательных веществ.[1] Этот круговорот охватывает большую часть Тихого океана и включает четыре преобладающих океанских течения: Северо-Тихоокеанское течение на север, Калифорнийское течение на восток Северное экваториальное течение на юг, а Куросио Текущий на запад. Его большие размеры и удаленность от берега привели к тому, что пробы NPSG были плохо отобраны и, следовательно, плохо изучены.[2]
Жизненные процессы в экосистемах открытого океана раковина для атмосфера увеличивается CO
2. Круговороты составляют значительную часть, примерно 75% того, что мы называем открытым океаном, или областью океана, не состоящей из прибрежных областей. Их считают олиготрофными или бедными питательными веществами, потому что они далеки от наземного стока.[3] Когда-то эти регионы считались однородными и статичными средами обитания. Однако появляется все больше свидетельств того, что NPSG демонстрирует существенную физическую, химическую и биологическую изменчивость в различных временных масштабах.[2] В частности, NPSG демонстрирует сезонные и межгодовые изменения в первичная продуктивность (определяется просто как производство нового растительного материала), что важно для усвоения CO
2.
NPSG - это не только приемник для CO
2 в атмосфере, а также другие загрязнители. Как прямой результат этого кругового рисунка, круговороты действуют как гигантские водовороты и становятся ловушками для антропогенных загрязнителей, таких как морской мусор. NPSG получил признание за большое количество пластикового мусора, плавающего прямо под поверхностью в центре круговорота. Эта область в последнее время привлекла много внимания средств массовой информации и обычно называется Большой тихоокеанский мусорный полигон.
История открытия
NPSG не часто отбирается из-за удаленности от побережья и нехватки морская жизнь. Эти огромные и глубокий океан Воды, вдали от влияния суши, исторически считались океаническим эквивалентом наземных пустынь с низкими запасами биомассы и низкими темпами производства. Эта точка зрения вытекает из недостаточного количества всесторонних исследований местообитаний центральных круговоротов. За последние два десятилетия эти взгляды были оспорены новым пониманием динамики NPSG.[4]

В первые дни морских исследований HMSПретендент (1872–1876 гг.), На пути от Иокогамы до Гонолулу, собрал образцы растений и животных, а также многочисленные образцы морской воды.[2] Целью этой экспедиции было определение химический состав морской воды и органическая материя во взвешенном состоянии и для изучения распространения и численности различных сообществ организмов. Мотивация к изучению экосистем открытого океана со временем изменилась, в то время как сегодня более современные исследования сосредоточены на биоразнообразии и влиянии климата на динамику экосистем. Сегодня программа «Временные ряды Гавайских океанов» (HOT) собрала крупнейшие и наиболее полные экологические набор данных для NPSG и планируется продолжить следующее тысячелетие.[2] Такие программы, как HOT, опровергли гипотезу о том, что эта экосистема статична и однородна, обнаружив, что NPSG демонстрирует динамические сезонные закономерности, отделяющие ее от других систем открытого океана.
Физические характеристики
NPSG является крупнейшей из сред обитания в открытом океане и считается крупнейшей смежной территорией Земли. биом.[5] Эта великая антициклоническая циркуляция простирается от 15 ° до 35 ° северной широты и от 135 ° до 135 ° западной долготы. Его площадь поверхности составляет примерно 2 x 107 км2. Его западная часть, к западу от 180 ° долготы, имеет большую физическую изменчивость, чем восточная часть. Эта изменчивость, когда разные погодные условия по-разному влияют на субрегионы, объясняется большими размерами этого круговорота.[2]
Эта большая изменчивость вызвана дискретными вихрями, почти инерционными движениями и внутренние приливы.[2] Климатические модели, такие как круговорот северной части Тихого океана (NPGO), Эль-Ниньо / южное колебание (ENSO) и Тихоокеанская декадная осцилляция (PDO), влияют на межгодовую изменчивость первичной продуктивности в NPSG.[3] DiLorenzo et al., 2008 Эти условия могут иметь глубокое влияние на биологические процессы в этой среде обитания,[2] у них есть способность менять температура поверхности моря (SST), структура хлорофилла, структура питательных веществ, концентрация кислорода, смешанный слой глубины, и, следовательно, грузоподъемность (количество жизни, которое может вместить эта среда обитания) NPSG.
Круговорот питательных веществ
Низкие концентрации питательных веществ и, следовательно, низкая плотность живые организмы характеризуют поверхностные воды НПС. Низкий биомасса приводит к чистой воде, позволяя фотосинтез происходить на значительную глубину. NPSG классически описывается как двухуровневая система. Верхний слой с ограниченным содержанием питательных веществ составляет большую часть основное производство, поддерживаемые в основном переработанными питательными веществами. Нижний слой содержит более доступные питательные вещества, но фотосинтез ограничен светом.[4]
В системах открытого океана биологическое производство зависит от интенсивной рециркуляции питательных веществ в эвфотический (солнечная) зона, лишь небольшая часть которой поддерживается поступлением «новых» питательных веществ.[6] Раньше считалось, что NPSG - это морская пустыня и что «новые» питательные вещества обычно не добавлялись в эту систему. Мнение изменилось, поскольку ученые начали лучше понимать эту среду обитания.[2] Хотя довольно высокие темпы первичной продукции поддерживаются за счет быстрой переработки питательных веществ,[4] физические процессы, такие как внутренние волны и приливы, циклонические мезомасштабные водовороты, ветровая накачка Экмана и атмосферные штормы могут нести новые питательные вещества.[7]
Питательные вещества, которые не попадают на поверхность, в конечном итоге оседают и питают среду обитания на морском дне. Глубоко бентические среды обитания Считалось, что океанические круговороты обычно состоят из некоторых из самых бедных с пищей регионов на планете.[8] Одним из источников питательных веществ для этой глубоководной среды обитания океана является морской снег. Морской снег состоит из детрита, мертвого органического вещества, которое выпадает из поверхностных вод, где продуктивность наиболее высока, и переносит углерод и азот из поверхностного смешанного слоя в глубину океана. Данные об обилии морского снега до глубины дно океана отсутствует в этой большой экосистеме.[9] Однако Pilskaln et al. обнаружили, что в NPSG количество морского снега было больше, чем ожидалось, и было удивительно сопоставимо с системой глубокого прибрежного апвеллинга.
Более высокая питательная ценность может быть связана с матами Rhizosolenia, которые также играют важную роль в формировании морского снега в субтропических круговоротах. Обычно это многовидовые ассоциации видов диатомовых Rhizosolenia. Этот более крупный фитопланктон может достигать в размерах до 10 сантиметров.[9] Этих матов особенно много в НПС. Их изобилие в этой экосистеме предполагает более высокий приток питательных веществ в NPSG, чем предсказывали классические теории.

Хотя этот механизм переносит азот на большую глубину, поверхностные воды потенциально отрезаны от этого источника. Азот должен быть доступен для жизни на поверхности. Чтобы объяснить недостаток азота на поверхности, существуют организмы, способные к азотфиксация в НПС. Триходесмий - один из видов, способных фиксировать азот, который встречается во многих цветениях поверхностного планктона.[7] Фиксация азота - это процесс, в котором инертный N2 берется из атмосферы и превращается в азотное соединение, доступное для использования организмами. Во многих олиготрофных морские экосистемы азотфиксация - это общий источник азота.
Вертикально мигрирующий зоопланктон также может активно переносить питательные вещества в разные зоны столб воды. Зоопланктон ночью питаются в поверхностных водах, а днем выпускают фекальные гранулы в средние слои воды, которые могут переносить C, N и P в более глубокие воды. В NPSG сообщество зоопланктона не статично, а колеблется в зависимости от сезона, и в нем преобладают веслоногие рачки, эвфаузииды и хетогнаты.[6]
Недавно классические теории о нехватке питательных веществ в NPSG были опровергнуты, и новые теории предполагают, что экосистема на самом деле динамична и характеризуется сильной сезонной, межгодовой и даже десятилетней изменчивостью.[9] Он также считается очень чувствительным к изменение климата, ученые наблюдали увеличение стратификации водной толщи и снижение доступности неорганических питательных веществ. Эти изменения предлагаются в качестве движущих механизмов, которые изменяют текущую тенденцию в структуре сообщества фитопланктона с эукариотических популяций на прокариотические, поскольку эти более простые организмы могут выдерживать меньшее поступление питательных веществ.[9] Зоопланктон и фитопланктон составляют менее 10% живых организмов в этом регионе, и в настоящее время хорошо задокументировано, что NPSG является «микробной экосистемой».[2]
Микробное сообщество
Микробный организмы составляют большинство первичные производители в НПС. Они есть автотрофный, что означает, что они собирают свою «пищу» из солнечного света и химикатов, в том числе CO
2. Эти организмы составляют основу пищевая цепочка, и поэтому их присутствие в экосистеме является фундаментальным. В NPSG первичная продуктивность часто описывается как низкая.
До 1978 года ученые предположили, что диатомовые водоросли преобладали в популяциях планктона в NPSG. Предполагалось, что первичными потребителями будет относительно крупный мезозоопланктон.[2] Сейчас хорошо известно, что большинство водорослей в NPSG на самом деле являются бактериями (одноклеточными организмами), в которых преобладают цианобактерии, или сине-зеленые водоросли. Эти простые организмы составляют большую часть постоянного запаса фотосинтезирующих морских обитателей в этой экосистеме. Ученые также недавно открыли археи (также одноклеточные микроорганизм, но больше похожи на эукариот, чем на бактерии) гены в NPSG, предполагая, что в этой среде обитания существует дополнительное разнообразие. В этом круговороте может существовать множество микроорганизмов, потому что маленький размер тела имеет конкурентное преимущество в океане для получения ресурсов (света и питательных веществ).[2] С современной точки зрения NPSG, микробная пищевая сеть присутствует всегда, в то время как более крупная пищевая цепь эукариот-травоядная носит сезонный и эфемерный характер.[2]
Сообщество эукариотического планктона
Эукариотический Планктон в круговороте зависит от «новых» питательных веществ, поступающих в результате физических погодных условий. В классической двухслойной модели, рассмотренной в предыдущих разделах, верхний слой рассматривается как «вращающееся колесо» с небольшим экспортом питательных веществ, поскольку они постоянно перерабатываются. Эта модель не позволяет вводить новые питательные вещества, что проблематично, поскольку это сделало бы невозможным быстрое увеличение или цветение фитопланктона. Несмотря на постоянное ограничение по питательным веществам в верхней части, биомасса планктона и скорость первичной продукции имеют значительную временную изменчивость и действительно приводят к цветению в NPSG.[3]
Эта межгодовая изменчивость объясняется изменениями в снабжении питательными веществами верхних слоев океана, вызванными физическими изменениями, вызванными ЭНСО и ЗОП.[3] Основываясь на новых данных, теперь выясняется, что нынешние темпы первичной продукции в этих регионах с низким содержанием питательных веществ намного выше, чем считалось ранее, и могут значительно варьироваться во временных масштабах от суточных до междесятилетних.[2] Весной иногда наблюдается быстрое увеличение поверхностного фитопланктона в связи с циклоническими мезомасштабными вихрями или интенсивными атмосферными возмущениями - оба физических процесса, которые приносят новые питательные вещества.[4] Летом цветение наблюдается более регулярно и, как правило, преобладают диатомовые водоросли и цианобактерии. Это регулярное летнее цветение может быть вызвано вариациями PDO.[3] Летнее цветение в этих водах наблюдается с тех пор, как их посещают исследовательские суда. Все эти цветения наблюдались в восточной части NSPG, и ни о каком цветении не сообщалось к западу от 160 ° з.[4] Гипотезы, объясняющие это явление, заключаются в том, что круговорот характеризуется низким содержанием фосфатов, но что цветение восточной NPSG имеет значительно более высокие концентрации фосфатов, чем западные.[4]
Вариации первичной продукции в NPSG могут существенно повлиять на круговорот питательных веществ, пищевой сети динамика и глобальные потоки элементов.[3] Гранулометрический состав пелагических первичных продуцентов определяет как состав, так и количество выносимых питательных веществ в более глубокие воды.[2] Это, в свою очередь, влияет на сообщества, которые живут в более глубоких водах этой системы.
Мезопелагическое сообщество
В мезопелагическая зона иногда называют сумеречной зоной; он простирается от 200 м до 1000 м. В более глубоких слоях NPSG виды, находящиеся выше в пищевой цепи, будут мигрировать вертикально или горизонтально в круговороте, в него и из него. Согласно анализу сообщества зоопланктона, центральная часть северной части Тихого океана имеет высокий видовое разнообразие (или большое количество видов) и высокая равноправность (имеется в виду относительно равное количество каждого вида). Также существует низкая степень сезонной изменчивости плотности зоопланктона.[2]
Исследования мезопелагических рыб центральных субтропических вод немногочисленны. Те немногие исследования, которые существуют, показали, что мезопелагические виды рыб не являются равномерно распределены по всей субтропической части Тихого океана. Их географические ареалы соответствуют моделям, показанным зоопланктоном. Некоторые из найденных видов ограничиваются этими низкопродуктивными центральными круговоротами. Некоторые из широко представленных семейств рыб - это Mytophids, Gonostomatids, Photichthyids, Sternoptychids и Melamphaids.[10] Наше понимание мезопелагического сообщества NPSG страдает от нехватки данных из-за сложности доступа к более глубоким зонам этой системы.
Бентическое сообщество
Самое глубокое сообщество в NPSG - это бентосный сообщество. В глубине круговорота лежит морское дно мелкозернистый глинистые отложения. Этот осадок является домом для сообщества организмов, которые обычно получают свои питательные вещества в виде «дождя» продуктивности, опускающегося сверху. На глубине под круговоротом находится одна из самых бедных с пищей областей на планете, поэтому здесь очень низкая плотность и биомасса донной инфауны или животных, обитающих в отложениях.[8] В самих отложениях содержание питательных веществ, включая углерод, хлорофилл и азот, обычно уменьшается с глубиной. Плотность придонной инфауны соответствует этой структуре питательных веществ. Инфауны обычно встречаются в более мелких слоях отложений, где граница раздела отложений и воды лежит и обычно уменьшается в количестве с увеличением глубины в осадке.[11] Бактерии в отложениях демонстрируют эту закономерность, а также макрофауна (инфаунальные организмы> 0,5 мм), в которых преобладают агглютинирующие (мягкотелые) фораминиферы и нематоды. Другая известная макрофауна, обнаруженная в отложениях, - это известковые фораминиферы, веслоногие рачки, полихеты и двустворчатые моллюски.[11] Эти бентосные организмы во многом зависят от поступления питательных веществ, которые оседают на морском дне. Любое изменение первичной продукции на поверхности может создать серьезную угрозу для этих организмов, а также вызвать другие потенциальные негативные последствия для других частей NPSG.
Будущее и значение NPSG

До недавнего времени NPSG считалась статичной частью обширной глобальной морской пустыни. Недавние открытия доказали, что эта система динамична и содержит физическую, химическую и биологическую изменчивость в различных временных масштабах. В связи с текущим изменением климата закономерности в атмосфере меняются и вызывают изменения в первичной продукции в NPSG. Изменения в первичной продуктивности могут повлиять на океан цикл углерода и потенциально атмосферный CO
2 и климат, потому что такие изменения могут изменить количество углерода, хранящегося в приповерхностных слоях океанов.[12] Поскольку NPSG - это крупнейший непрерывный биом на Земле, он важен не только для сообщества организмов, но и для всей планеты.
NPSG привлекла к себе пристальное внимание из-за другой проблемы, с которой она сейчас сталкивается. Вихревые эффекты круговорота служат для удержания загрязняющих веществ в его центре. Если загрязняющее вещество попадает в ловушку в потоке, который движется к круговороту, он будет оставаться там бесконечно или до тех пор, пока не истечет срок жизни загрязняющего вещества. Одним из таких стойких загрязнителей, часто встречающихся в НПС, является пластиковый мусор. NPSG забрасывает обломки в центральную зону. Это явление недавно дало этому круговороту прозвище «Тихоокеанский мусорный участок». Среднее количество и вес пластиковых частей в этом районе в настоящее время являются самыми большими, наблюдаемыми в Тихом океане.[13] Ходят слухи, что этот пластиковый «суп» имеет размер от Техаса до США. С ростом интереса к загрязнению и изменению климата NPSG привлекает все больше внимания. Важно, чтобы наши знания об этой системе продолжали развиваться по этим причинам, а также исключительно для понимания самой большой экосистемы в мире.
Смотрите также
Рекомендации
- ^ Порецкий, 2009 г.
- ^ а б c d е ж грамм час я j k л м п о Карл, Д. 1999
- ^ а б c d е ж Корно и др., 2007 г.
- ^ а б c d е ж Дор и др., 2008 г.
- ^ (Карл и др., 2002)
- ^ а б Hannides et al., 2009 г.
- ^ а б Николсон и др., 2008 г.
- ^ а б (Шуленбергер и Хесслер, 1974)
- ^ а б c d Pilskaln et al., 2005
- ^ Барнетт, 1984
- ^ а б Смит-младший и др., 2002 г.
- ^ Brix et al., 2006
- ^ Мур и др., 2001
Источники
- Барнетт, М.А. (1984). «Зоогеография мезопелагических рыб в центральной тропической и субтропической частях Тихого океана: видовой состав и структура в репрезентативных местах в трех экосистемах». Морская биология. 82 (2): 199–208. Дои:10.1007 / BF00394103.
- Брикс, Х., Грубер, Н., Карл, Д., и Н. Бейтс. 2006. О взаимосвязи между первичным, сетевым сообществом и экспортной продукцией в субтропических круговоротах. Глубокое море Часть II исследования. (53) 698-717.
- Корно, Г., Карл, Д., Черч, М., Летелье, Р., Лукас, Р., Бидигар, Р. и М. Эбботт. 2007. Влияние климатическое воздействие по экосистемным процессам в субтропическом круговороте северной части Тихого океана. Журнал геофизических исследований. (112) 1-14.
- ДиЛоренцо Э., Шнайдер Н., Кобб К., Фрэнкс П., Чхак К., Миллер А., МакВильямс Дж., Боград С., Аранго Х., Курчицер Э., Пауэлл , Т. и П. Ривьер. 2008. North Pacific Gyre Oscillations связывает климат океана и изменение экосистемы. Письма о геофизических исследованиях. (35) 1-6.
- Dore, J.E .; Letelier, R.M .; Черч, М. Дж .; Лукас, Р .; Карл, Д. М. (2008). «Летнее цветение фитопланктона в олиготрофном субтропическом круговороте северной части Тихого океана: историческая перспектива и недавние наблюдения». Прогресс в океанографии. 76: 2–38. Дои:10.1016 / j.pocean.2007.10.002.
- Hannides, C.C.S .; Landry, M. R .; Бенитес-Нельсон, К.; Стили, Р. Е. М .; Montoya, J.P .; Карл, Д. М. (2009). «Экспортная стехиометрия и опосредованный мигрантами поток фосфора в субтропическом круговороте северной части Тихого океана». Deep-Sea Research Part I: Oceanographic Research Papers. 56: 73–88. Дои:10.1016 / j.dsr.2008.08.003.
- Карл, Д. М. (1999). «Мини-обзоры: Море перемен: биогеохимическая изменчивость в субтропическом круговороте северной части Тихого океана» (PDF). Экосистемы. 2 (3): 181–214. Дои:10.1007 / с100219900068. JSTOR 3658829.
- Карл, Д. М .; Лукас, Р. (1996). «Программа временного ряда Гавайских океанов (HOT): предыстория, обоснование и практическая реализация». Deep-Sea Research Part II: Актуальные исследования в океанографии. 43 (2–3): 129. Дои:10.1016/0967-0645(96)00005-7.
- Карл, Д. М .; Bidigare, R. R .; Летелье Р. М. (2002). «Устойчивая и апериодическая изменчивость в производстве органических веществ и структура сообщества фототрофных микробов в субтропическом круговороте северной части Тихого океана». Продуктивность фитопланктона. п. 222. Дои:10.1002 / 9780470995204.ch9. ISBN 9780470995204.
- Мур, К., Мур, С., Ликастер, М., и С. Вейсберг. 2001. Сравнение пластика и планктона в центральном круговороте северной части Тихого океана. Загрязнение морской среды Бюллетень. (42) номера страниц.
- Николсон, Дэвид; Эмерсон, Стивен; Эриксен, Чарльз К. (2008). «Сетевое производство сообществ в глубокой эвфотической зоне субтропического круговорота северной части Тихого океана по данным планерных исследований» (PDF). Лимнология и океанография. 53 (5 часть 2): 2226–2236. Дои:10.4319 / lo.2008.53.5_part_2.2226. Получено 11 ноября 2017.
- Pilskaln, C.H .; Villareal, T. A .; Dennett, M .; Darkangelo-Wood, C .; Медоуз, Г. (2005). «Высокая концентрация морского снега и матов диатомовых водорослей в субтропическом круговороте северной части Тихого океана: влияние на циклы углерода и азота в олиготрофном океане». Deep-Sea Research Part I: Oceanographic Research Papers. 52 (12): 2315. Дои:10.1016 / j.dsr.2005.08.004. HDL:1912/404.
- Порецкий, Р. С .; Hewson, I .; Sun, S .; Allen, A.E .; Zehr, J. P .; Моран, М. А. (2009). «Сравнительный метатранскриптомический анализ день / ночь микробных сообществ в субтропическом круговороте северной части Тихого океана». Экологическая микробиология. 11 (6): 1358–75. Дои:10.1111 / j.1462-2920.2008.01863.x. PMID 19207571.
- Shulenberger, E .; Хесслер, Р. Р. (1974). «Поедающие глубинные бентосные амфиподы, пойманные в ловушку олиготрофных центральных вод северной части Тихого океана». Морская биология. 28 (3): 185. Дои:10.1007 / BF00387296.
- Smith, K. L .; Болдуин, Р. Дж .; Карл, Д. М .; Боэтиус, А. (2002). «Реакция бентосных сообществ на зернобобовые в пелагических пищевых продуктах: субтропический круговорот в северной части Тихого океана». Deep-Sea Research Part I: Oceanographic Research Papers. 49 (6): 971. Дои:10.1016 / S0967-0637 (02) 00006-7.

