Форонид - Phoronid
| Форониды | |
|---|---|
 | |
| Научная классификация | |
| Королевство: | Animalia |
| Подцарство: | Eumetazoa |
| Clade: | ParaHoxozoa |
| Clade: | Bilateria |
| Clade: | Нефрозоа |
| (без рейтинга): | Протостомия |
| (без рейтинга): | Спиралия |
| Суперфилум: | Лофотрохозоа |
| Clade: | Лофофората |
| Clade: | Брахиозоа |
| Тип: | Phoronida Hatschek, 1888 |
| Роды | |
| |
Форониды (научное название Phoronidaиногда называют подковообразные черви) маленькие филюм морских животных, которые фильтр-корм с лофофор («корона» из щупалец) и построить вертикальные трубки из хитин чтобы поддерживать и защищать свои мягкие тела. Они обитают в большинстве океанов и морей, включая Северный Ледовитый океан, но исключая Антарктический океан, и между приливной зона и около 400 метров вниз. Большинство взрослых форонид имеют длину 2 см и ширину около 1,5 мм, хотя самые крупные имеют длину 50 см.
Название группы происходит от ее типовой род: Форонис.[2][3]
Обзор
Нижний конец тела представляет собой ампулу (припухлость, похожую на колбу), которая закрепляет животное в трубке и позволяет ему очень быстро втягивать свое тело в случае угрозы. Когда лофофор расширяется в верхней части тела, реснички (маленькие волоски) по бокам щупалец притягивают частицы пищи ко рту, который находится внутри и немного сбоку от основания лофофора. Нежелательный материал можно исключить, закрыв крышку над ртом, или отбросить щупальца, реснички которых могут переключаться в обратном направлении. Затем пища опускается в желудок, который находится в ампуле. Твердые отходы перемещаются вверх по кишечнику и выводятся наружу через анус, которая находится снаружи и немного ниже лофофора.
Кровеносный сосуд ведет вверх по середине тела от желудка к круглому сосуду у основания лофофора, а оттуда по одному слепому сосуду поднимается вверх каждое щупальце. Пара кровеносных сосудов у стенки тела ведет вниз от кольца лофофора к желудку, а также к слепым ветвям по всему телу. Сердца нет, но основные сосуды могут волнообразно сокращаться, перемещая кровь. Форониды не вентилируют свои стволы насыщенной кислородом водой, а полагаются на дыхание через лофофор. В крови содержится гемоглобин, что необычно для таких мелких животных и кажется адаптацией к аноксический и гипоксический среды. Кровь Phoronis architea несет вдвое больше кислород как человек того же веса. Два метанефридия фильтровать жидкость организма, возвращая любые полезные продукты и сбрасывая оставшиеся растворимые отходы через пару пор рядом с анальным отверстием.
Один вид строит колонии подающий надежды или путем разделения на верхнюю и нижнюю части, и все форониды размножаться половым путем с весны до осени. Яйца большинства видов образуют свободно плавающих личинок актинотрохов, которые питаются планктоном. Актинотрох оседает на морское дно примерно через 20 дней, а затем претерпевает радикальные изменения за 30 минут: личиночные щупальца заменяются взрослыми лофофорами; анус перемещается снизу к краю лофофора; и это меняет кишечник с вертикального на U-образный, с животом внизу тела. Один вид образует личинку, похожую на слизняк, а личинки нескольких видов неизвестны. Живут форониды около года.
Некоторые виды живут отдельно, в вертикальных трубках, погруженных в мягкий осадок, в то время как другие образуют спутанные массы, погребенные в скалах и ракушках или покрытые коркой. В некоторых местообитаниях популяции форонид достигают десятков тысяч особей на квадратный метр. Личинки актинотроха известны среди планктон, а иногда и составляют значительную часть биомассы зоопланктона. К хищникам относятся рыбы, брюхоногие моллюски (улитки) и нематоды (крошечные круглые черви). Один вид форонид неприятен многим эпибентосный хищники. Различный паразиты поражают полости тела форонид, пищеварительный тракт и щупальца. Неизвестно, имеют ли форониды какое-либо значение для человека. В Международный союз охраны природы (МСОП) не внес в список исчезающих видов форонид.
По состоянию на 2010 г. бесспорных окаменелостей тел форонид нет.[4] Есть веские доказательства того, что форониды создали следы окаменелостей найдено в Силурийский, Девонский, Пермский период, Юрский период и Меловой периоды, и, возможно, в Ордовик и Триасовый. Форониды, брахиоподы и мшанки (эктопрокты) собирательно называются лофофораты, потому что все используют лофофоры для кормления. Примерно с 1940-х по 1990-е гг. родословные на основе эмбриологический и морфологический особенности помещают лофофораты среди или как сестринская группа к дейтеростомы, супер-тип, который включает хордовые и иглокожие. Хотя меньшинство придерживается этой точки зрения, большинство исследователей теперь рассматривают форониды как членов протостом супертип Лофотрохозоа.[5] Хотя аналитики, использующие молекулярная филогения уверены, что члены Lophotrochozoa более тесно связаны друг с другом, чем с нечленами, отношения между членами в основном неясны. Некоторые анализы рассматривают форонид и брахиопод как сестринские группы, в то время как другие относят форонид к подгруппе брахиопод.[6]
Сравнение похожих типов
| Особенность | Форониды[7] | Брахиоподы[8] | Мшанки[9] | Энтопрокты[10] |
|---|---|---|---|---|
| Щупальца полые | да | да | да | Нет |
| Защита и поддержка | Прямая трубка хитин | Корпус с двумя клапанами | Различные, в том числе хитин, минерализованный скелеты, растительные формы и масса студенистого материала | Никто |
| Поток кормления | Сверху вниз | Через стороны оболочки, через переднюю | Сверху вниз | Снизу вверх |
| Анус | Наружное кольцо щупалец | в мантия, или ничего и твердые отходы выбрасываются изо рта[11] | Наружное кольцо щупалец | Внутреннее кольцо щупалец |
| Колониальный | Один вид | Нет | Все, кроме одного рода | Большинство видов колониальных |
| Целом | да | да | да | Нет |
Описание
Структура тела
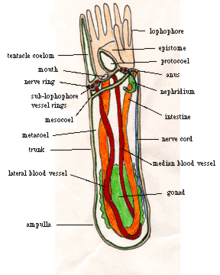
Большинство взрослых форонид имеют длину от 2 до 20 см и ширину около 1,5 мм.[7] хотя самые большие 50 см в длину.[12] В их шкурах нет кутикула но выделять жесткие трубы хитин,[7] похож на материал, используемый в членистоногие ' экзоскелеты,[13] и иногда подкрепляется осадок частицы и другой мусор.[1] Трубки у большинства видов прямостоячие, но у Phoronis vancouverensis горизонтальные и запутанные.[14] Форониды могут перемещаться внутри своих трубок, но никогда не покидают их.[7] Нижний конец корпуса представляет собой ампулу (набухание в виде колбы в трубчатой структуре.[15]),[7] который закрепляет животное в трубе и позволяет ему втягивать свое тело при угрозе,[12] уменьшение тела до 20 процентов от его максимальной длины.[1] Продольные мышцы втягивают тело очень быстро, а круговые мышцы медленно расширяют тело, сжимая внутреннюю жидкость.[12]
Для кормления и дыхание каждый форонид имеет на верхнем конце лофофор, «корона» из щупалец, с которой животное фильтры. У мелких видов «корона» представляет собой простой круг, у средних видов изогнута в виде круга. подкова с щупальцами на внешней и внутренней сторонах, а у самых крупных видов концы подковы закручиваются в сложные спирали. Эти более сложные формы увеличивают площадь, доступную для кормления и дыхание.[7] Щупальца полые, удерживаются в вертикальном положении за счет давления жидкости и могут двигаться индивидуально с помощью мускулов.[12]
Рот находится внутри основания макушки щупалец, но сбоку. Кишечник идет от рта к одной стороне желудок, в нижней части ампулы. Кишечник идет от желудка вверх по другой стороне тела и выходит через задний проход, снаружи и немного ниже макушки щупалец. Кишечник и кишечник поддерживаются двумя брыжейки (перегородки, которые проходят по длине тела) соединены со стенкой тела, а другая брыжейка соединяет кишечник с кишечником.[7]
Тело делится на coeloms,[7] отсеки выложены мезотелий.[16] Основная полость тела, расположенная под короной щупалец, называется Metacoelom, а щупальца и их основание разделяют мезоцелом.[7] Над ртом находится эпистом, полая крышка, которая может закрыть рот.[12] Полость в эпистоме иногда называют протокеломом, хотя другие авторы не согласны с тем, что это целом[17] а Рупперт, Фокс и Барнс думают, что это построено с помощью другого процесса.[7]
Трубка состоит из трехслойного внутреннего цилиндра из органического материала и внешнего агглютинированного слоя.[18]
Питание, кровообращение и выделение
Когда лофофор расширен, реснички (маленькие волоски) по бокам щупалец вытягивают воду между щупальцами к основанию лофофора. Более короткие реснички на внутренней стороне щупалец выталкивают частицы пищи в желобок по кругу под щупальцами и прямо внутри них, а реснички в канавке толкают частицы в рот.[12] Форониды направляют свои лофофоры в водное течение и быстро переориентируются, чтобы максимизировать зону ловли пищи при изменении течения. Их диета включает водоросли, диатомеи, жгутиконосцы, перидинийцы, мелкие личинки беспозвоночных и детрит.[1] Нежелательный материал можно исключить, закрыв эпистом (крышку над ртом), или отбросить щупальца, реснички которых могут переключаться в обратном направлении. Кишечник использует реснички и мышцы для перемещения пищи к желудку и выделения ферменты которые переваривают часть пищи, но желудок переваривает большую часть пищи.[12] Форониды также поглощают аминокислоты (строительные блоки белки[19]) через кожу, в основном летом.[1] Твердые отходы перемещаются вверх по кишечнику и выводятся наружу через анус, которая находится снаружи и немного ниже лофофора.[20]
Кровеносный сосуд[7] начинается с брюшина (в мембрана который свободно охватывает живот),[12] со слепым капилляры снабжая желудок.[7] Кровеносный сосуд ведет вверх по центру тела к круглому сосуду у основания лофофора, и оттуда по одному слепому сосуду поднимается вверх каждое щупальце. Пара кровеносных сосудов около стенки тела ведет вниз от кольца лофофоров, и у большинства видов они объединены в один, расположенный немного ниже кольца лофофора. Нисходящий сосуд (сосуды) ведет обратно к брюшине, а также к слепым ветвям по всему телу. Сердца нет, но мышцы в основных сосудах волнообразно сокращаются для перемещения крови.[12] В отличие от многих животных, которые живут в трубках, форониды не вентилируют свои хоботы насыщенной кислородом водой, а полагаются на дыхание лофофором, который простирается выше гипоксический отложения. В крови есть гемоциты, содержащие гемоглобин, что необычно для таких мелких животных и кажется адаптацией к аноксический и гипоксический среды. Кровь Phoronis architea переносит столько же кислорода на см3 как у большинства позвоночные; объем крови в см3 на грамм веса тела вдвое больше, чем у человека.[7]
Подоциты На стенках кровеносных сосудов проводят первую ступень фильтрации растворимых отходов в основную жидкость целом. Два метанефридия, каждый с воронкообразным забором, фильтруют жидкость второй раз,[7] возвращение любых полезных продуктов в coelom[21] и сброс оставшихся отходов через пару нефридиопоры рядом с анусом.[7]
Нервная система и движение
Между ртом и анусом находится нервный центр, а у основания лофофора - нервное кольцо.[1] Кольцо подводит нервы к щупальцам и непосредственно под кожей к мышцам стенки тела. Phoronis ovalis имеет два нервных ствола под кожей, тогда как у других видов есть один.[7] В багажнике (ах) есть гигантские аксоны (нервы, которые очень быстро передают сигналы), которые координируют втягивание тела при угрозе опасности.[12]
За исключением втягивания тела в трубку, форониды имеют ограниченное и медленное движение: частичный выход из трубки; сгибание тела при разгибании; и лофофор бросает еду в рот.[7]
Размножение и жизненный цикл
Только Phoronis ovalis естественным образом строит колонии подающий надежды или путем разделения на верхнюю и нижнюю части, которые затем превращаются в полные тела. В экспериментах другие виды успешно разделились, но только тогда, когда у обеих частей достаточно гонадный (репродуктивный[22]) ткань.[23] Все форониды размножаться половым путем с весны до осени. Некоторые виды гермафродитный (имеют как мужские, так и женские репродуктивные органы[24]) но перекрестное оплодотворение (оплодотворять яйца других членов[25]), а другие раздельнополый (иметь отдельные полы[26]).[1] В гаметы (сперма и яйцеклетка[27]) образуются в опухших гонадах вокруг желудка.[7] Гаметы плывут через метакоелому к метанефридиям.[12] Сперматозоиды выходят из нефридиопор, а некоторые из них захватываются лофофорами особей того же вида. Виды, откладывающие небольшие оплодотворенные яйца, выпускают их в воду в виде планктона,[1] в то время как виды с более крупными яйцами выводят их либо в трубке тела, либо приклеиваются клеем в центре лофофора.[12] Выведенные яйца выпускаются, чтобы питаться планктоном, когда они развиваются в личинок.[1]
Развитие яиц представляет собой смесь дейтеростом и протостом характеристики. Рано подразделения Яйца бывают холобластическими (клетки делятся полностью) и радиальными (постепенно образуют стопку кругов). Процесс носит регулирующий характер (судьба каждой клетки зависит от взаимодействия с другими клетками, а не от жесткой программы в каждой клетке), и эксперименты, в ходе которых делились ранние эмбрионы, дали полных личинок. Мезодерма сформирован из мезенхима происходящий из архентерон Целом образован шизокоэлия, а бластопор (вмятина на эмбрионе) становится ртом.[7]

Слизневая личинка Phoronis ovalis плавает около 4 дней, ползает по морскому дну от 3 до 4 дней, затем впивается в карбонатный дно.[28][29] О трех видах ничего не известно. Остальные виды развивают свободное плавание. актинотроч личинки, питающиеся планктоном. Актинотрох представляет собой вертикальный цилиндр с анусом внизу и окаймленным ресничками. Вверху находится мочка[1] или капюшон, под которым находятся: ганглий, соединенный с участком ресничек за пределами верхушки капюшона;[7] пара протонефридия (мельче и проще, чем метанефридии у взрослых);[1] рот; и кормящие щупальца, окружающие рот.[7] После 20-дневного плавания актинотрок оседает на морском дне и подвергается катастрофическая метаморфоза (радикальное изменение) через 30 минут: капюшон и щупальца личинки впитываются, и вокруг рта образуется взрослый лофофор, и оба теперь указывают вверх; кишечник имеет U-образный изгиб, так что анус находится прямо под лофофором и снаружи.[1] Наконец, взрослый форонид строит трубку.[7]
Форониды живут около года.[1]
Экология

Форониды обитают во всех океанах и морях, включая Арктический [30] и за исключением Антарктический океан,[1] и появляются между приливной зона и около 400 метров вниз. Некоторые встречаются отдельно, в вертикальных трубах, погруженных в мягкий осадок, такой как песок, грязь или мелкий гравий. Другие образуют спутанные массы из множества людей, погребенных в скалах и ракушках или покрытых коркой. В некоторых местообитаниях популяции форонид достигают десятков тысяч особей на квадратный метр. Личинки актинотроха известны среди планктон,[1] и иногда составляют значительную часть биомассы зоопланктона.[31]
Phoronis australis отверстия в стенке трубки цериантид анемон, Ceriantheomorphe brasiliensis, и использует это как основу для создания собственной трубы. В одном цериантиде может находиться до 100 форонид. В этом неравные отношения анемон не приносит ни значительных преимуществ, ни вреда, в то время как форонид получает пользу от: основы для трубки; корм (оба зверя - фильтраторы); и защита, так как цериантид уходит в свою трубку, когда возникает опасность, и это предупреждает форонид втягиваться в свою трубку.[32]
Хотя хищники форонид малоизвестны, к ним относятся рыбы, брюхоногие моллюски (улитки) и нематоды (крошечные круглые черви).[1] Phoronopsis viridis, который достигает плотности 26 500 на квадратный метр на приливных отмелях в Калифорния (США), многим не нравится эпибентосный хищники, включая рыбу и крабов. Неприятность наиболее сильна в верхней части, включая лофофор, который подвергается воздействию хищников при кормлении форонид. Когда лофофоры были удалены в эксперименте, форониды были более вкусными, но этот эффект уменьшился в течение 12 дней по мере регенерации лофофоров. Эти широко эффективные средства защиты, которые кажутся необычными среди беспозвоночных, обитающих в мягких отложениях, могут иметь важное значение для разрешения Phoronopsis viridis для достижения высокой плотности.[33] Немного паразиты заражают форониды: прогенетический метацеркарии и кисты из трематоды в форонидах целомический полости; неопознанный грегарины в пищеварительном тракте форонид; и анцистрокомид инфузория паразит, Heterocineta, в щупальцах.[1]
Неизвестно, имеют ли форониды какое-либо значение для человека. В Международный союз охраны природы (МСОП) не внес в список исчезающих видов форонид.[1]
Эволюционная история
Окаменелости

По состоянию на 2016 год бесспорных окаменелостей тел форонид нет.[4] Исследование нижнего Кембрийский Окаменелости Чэнцзян, в 1997 году Чен и Чжоу переводили Иотуба Chengjiangensis как форонид, поскольку у него были щупальца и U-образная кишка,[35] а в 2004 году Чен переводил Эофоронис как форонид.[36] Однако в 2006 г. Конвей Моррис рассматривается Иотуба и Эофоронис как синонимы одного и того же рода, который, по его мнению, выглядел как приапулид Луизелла.[37] В 2009 году Бальтазар и Баттерфилд нашли на западе Канады два образца новой окаменелости, датируемых 505 миллионами лет назад. Lingulosacculus nuda, который имел две оболочки, как у брахиоподы но нет минерализованный. По мнению авторов, П-образная кишка выходила за линию шарнира и выходила за пределы меньшей раковины. Это предотвратило бы прикрепление мышц, чтобы закрывать и открывать раковины, и 50% длины животного за линией шарнира потребовали бы продольных мышц, а также кутикула для защиты. Следовательно, они предполагают, что Lingulosacculus мог быть членом форонида стволовая группа в пределах язычковый брахиоподы.[38] Другая альтернатива - это Эксцентротека[39][40] лежит где-то в линии родословной форонид.
Есть веские доказательства того, что виды Форонис создал следы окаменелостей из ихноген Тальпина, которые были обнаружены в Девонский, Юрский период и Меловой периоды.[41] Животное Тальпина просверлено в известковый водоросли, кораллы, ехиноид тесты (раковины), раковины моллюсков и ростра из белемниты.[42] Хедереллиды или гедереллоиды - это окаменелые трубки, обычно изогнутые и шириной от 0,1 до 1,8 мм, найденные в Силурийский к Пермский период, и, возможно, в Ордовик и Триасовый. Их ветвящиеся колонии могли быть образованы форонидами.[34]
Семейное дерево
Форониды, брахиоподы и мшанки (эктопрокты) собирательно называются лофофораты, потому что все питаются с помощью лофофоров.[5] Примерно с 1940-х по 1990-е гг. родословные на основе эмбриологический и морфологический особенности помещают лофофораты среди или как сестринская группа к дейтеростомы,[6] супер-тип, который включает хордовые и иглокожие. На раннем этапе развития эмбрионов дейтеростомы образуют анус перед ртом, пока протостомы сначала сформируйте рот.[43]
Нильсен (2002) рассматривает форониды и брахиоподы как связанные с дейтеростомом. крылышки, которые также фильтруют питание щупальцами, потому что токопроводящие клетки лофофоров всех трех имеют один ресничка на клетку, а лофофоры мшанки, которые он считает протостомами, имеют несколько ресничек на клетку.[44] Helmkampf, Bruchhaus и Hausdorf (2008) суммируют эмбриологический и морфологический анализ нескольких авторов, которые сомневаются или не соглашаются с тем, что форониды и брахиоподы являются дейтеростомами:[5]
- В то время как у дейтеростомов три целомический полостей, лофофоратов, таких как форониды и брахиоподы, всего две.[17]
- Pterobranchs могут быть подгруппой энтеропнев («желудевые черви»). Это говорит о том, что предковый дейтеростом больше похож на подвижный червеобразный энтеропнев, чем на сидячий колониальный птеробранх. Тот факт, что и лофофораты, и птеробранхи используют щупальца для питания, вероятно, не является синапоморфией лофофоратов и дейтеростомов, а развились независимо как сходящийся адаптации к сидячему образу жизни.[5][45][46]
- Мезодерма не образуется энтерокоэли у форонид и мшанок, но есть у дейтеростомов, в то время как существуют разногласия по поводу того, образуют ли брахиоподы мезодерму путем энтерокоэлии.[5]
| Bilateria |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Начиная с 1988 г., анализ основан на молекулярная филогения, который сравнивает биохимический такие особенности, как сходство в ДНК, поместили форонид и брахиопод среди Лофотрохозоа, супер-тип протостома, который включает моллюски, кольчатые червя и плоские черви но исключает другой главный супертип протостомов Экдизозоа, в состав которой входят членистоногие.[5][6] Коэн писал: «Этот вывод, если он верен, подрывает практически все основанные на морфологии реконструкции филогении, сделанные в течение последнего столетия или более».[48]
Хотя анализ молекулярной филогении дает уверенность в том, что члены Lophotrochozoa более тесно связаны друг с другом, чем с нечленами, отношения между членами в основном неясны.[6][49] В Лофотрохозоа обычно делятся на: Лофофората (животные, у которых есть лофофоры), включая Phoronida и Brachiopoda; Трохозоа (животные, многие из которых трохофор личинки), в том числе моллюсков, кольчатые червя, эхиураны, сипункулы и немертины; и некоторые другие типы (например, Платигельминты, Гастротрича, Гнатостомулида, Микрогнатозоа, и Коловратка ).[6]
Молекулярная филогения указывает на то, что Phoronida тесно связаны с Brachiopoda, но Мшанки (Ectoprocta) не имеют близкого родства с этой группой, несмотря на использование аналогичных лофофоров для питания и дыхания.[49][50][51] Это означает, что традиционное определение «Lophophorata» не монофилетический. В последнее время термин «лофофората» применялся только к Phoronida и Brachiopoda, и Галаныч считает, что это изменение вызовет путаницу.[6] Некоторые анализы рассматривают Phoronida и Brachiopoda как сестринские группы, в то время как другие относят Phoronida к подгруппе внутри Brachiopoda,[6] подразумевая, что Brachiopoda парафилетический.[52] Анализ Коэна и Вейдмана (2005) заключает, что форониды представляют собой подгруппу нечленораздельные брахиоподы (те, у которых шарнир между двумя клапанами не имеет зубцов и гнезд[8]) и сестринская группа других нечленораздельных подгрупп. Авторы также предполагают, что предки моллюски и клады брахиопод + форонид разошлись между 900 и 560 млн лет, вероятнее всего, около 685 млн лет.[51]
Таксономия
| Взрослые виды[1] | Виды личинок[1] |
|---|---|
| Phoronis ovalis | (ползучая личинка) |
| Phoronis hippocrepia | Актинотроха гиппокрепия[29] |
| Phoronis ijimai, также называется Phoronis vancouverensis | Актинотроча ванкуверенсис |
| Phoronis australis | (неизвестно) |
| Phoronis muelleri | Актинотроча жаберная |
| Phoronis psammophila | Актинотроча сабатьери |
| Phoronis pallida | Актинотроча бледная |
| Phoronopsis albomaculata | (неизвестно) |
| Phoronopsis harmeri | Актинотроча гармерия |
| Форонопсис калифорнийский | (неизвестно) |
Тип имеет два роды, без класс или порядок имена. Зоологи дали личинок, обычно называемых актинотроч, отдельное название рода от взрослых.[1]
В 1999 году Темерева и Малахов описали Phoronis svetlanae.[53] В 2000 году Темерева описала новый вид, Phoronopsis malakhovi,[54] в то время как Эмиг считает это синонимом Phoronopsis harmeri.[1] Сантагата думает Phoronis architea это другой вид от обоих Phoronis psammophila и Phoronis muelleri и что «видовое разнообразие [форонид] в настоящее время недооценивается».[55] В 2009 году Темерева описала, какими могут быть личинки Phoronopsis albomaculata и Форонопсис калифорнийский. Она написала, что, хотя существует 12 бесспорных взрослых видов форонид, идентифицировано 25 морфологических типов личинок.[31]
Примечания
использованная литература
- ^ а б c d е ж г час я j k л м п о п q р s т ты v ш Икс у Эмиг, Кристиан К. (2003). "Тип: Phoronida" (PDF). В Бернхарде Гржимеке; Девра Г. Клейман; Майкл Хатчинс (ред.). Энциклопедия жизни животных Гржимека. 2: Протостомы (2-е изд.). Томпсон Гейл. С. 491–495. ISBN 978-0-7876-5362-0. Получено 2011-03-01.
- ^ «Форонида». Словарь Merriam-Webster. "Новая латынь, от Форонис + -Ида."
- ^ «Форонис». Словарь Merriam-Webster. "Новая латынь, вероятно, с латыни Форонис (Ио, мифическая жрица Аргос которого любил Зевс) ".
- ^ а б Тейлор, Пол Д .; Олев Винн; Марк А. Уилсон (2010). «Эволюция биоминерализации в лофофоратах.'". Специальные статьи по палеонтологии. 84: 317–333. Дои:10.1111 / j.1475-4983.2010.00985.x (неактивно 19.11.2020).CS1 maint: DOI неактивен по состоянию на ноябрь 2020 г. (ссылка на сайт)
- ^ а б c d е ж Гельмкампф, Мартин; Ирис Брухгауз; Бернхард Хаусдорф (август 2008 г.). «Филогеномный анализ лофофоратов (брахиопод, форонид и мшанок) подтверждает концепцию Lophotrochozoa». Труды Королевского общества B. 275 (1645): 1927–1933. Дои:10.1098 / rspb.2008.0372. ЧВК 2593926. PMID 18495619.
- ^ а б c d е ж г час Галаныч, К.М. (15 декабря 2004 г.). «Новый взгляд на филогению животных» (PDF). Ежегодный обзор экологии, эволюции и систематики. 35: 229–256. Дои:10.1146 / annurev.ecolsys.35.112202.130124. Получено 2011-03-09.
- ^ а б c d е ж г час я j k л м п о п q р s т ты v ш Икс Рупперт, Э.Е., Фокс, Р.С., и Барнс, Р.Д. (2004). "Lophophorata: Phoronida". Зоология беспозвоночных (7-е изд.). Брукс / Коул. стр.817–821. ISBN 978-0-03-025982-1.CS1 maint: несколько имен: список авторов (ссылка на сайт)
- ^ а б Рупперт, Э.Е., Фокс, Р.С., и Барнс, Р.Д. (2004). «Брахиоподы». Зоология беспозвоночных (7-е изд.). Брукс / Коул. стр.821–829. ISBN 978-0-03-025982-1.CS1 maint: несколько имен: список авторов (ссылка на сайт)
- ^ Ruppert, E.E; Fox, R.S .; Барнс, Р.Д. (2004). «Мшанки». Зоология беспозвоночных (7-е изд.). Брукс / Коул. стр.829–845. ISBN 978-0-03-025982-1.
- ^ Рупперт, Э.Е., Фокс, Р.С., и Барнс, Р.Д. (2004). «Камптозоа и циклиофора». Зоология беспозвоночных (7-е изд.). Брукс / Коул. стр.808–812. ISBN 978-0-03-025982-1.CS1 maint: несколько имен: список авторов (ссылка на сайт)
- ^ Доэрти, П.Дж. (2001). «Лофофораты». В Андерсоне, Д.Т. (ред.). Зоология беспозвоночных (2-е изд.). Издательство Оксфордского университета. С. 356–363. ISBN 978-0-19-551368-4.
- ^ а б c d е ж грамм час я j k л м Доэрти, П.Дж. (1998). «Лофофораты - Phoronida, Brachiopoda и Ectoprocta». В Д.Т. Андерсоне (ред.). Зоология беспозвоночных (1-е изд.). Издательство Оксфордского университета, Австралия. С. 346–349. ISBN 978-0-19-553941-7.
- ^ Рупперт, Э.Е., Фокс, Р.С., и Барнс, Р.Д. (2004). «Членистоногие». Зоология беспозвоночных (7-е изд.). Брукс / Коул. стр.518. ISBN 978-0-03-025982-1.CS1 maint: несколько имен: список авторов (ссылка на сайт)
- ^ Хинтон, Сэм (1987). Морская жизнь южной Калифорнии: знакомство с животным миром калифорнийских пляжей к югу от Санта-Барбары. Калифорнийский университет Press. ISBN 978-0-520-05924-5. Получено 2011-08-28.
- ^ Моррис, Кристофер Г. (1992). «Ампулла». Научно-технический словарь Academic Press. Gulf Professional Publishing. п. 102. ISBN 978-0-12-200400-1.
- ^ Рупперт, Э.Е., Фокс, Р.С., и Барнс, Р.Д. (2004). «Компартментализация». Зоология беспозвоночных (7-е изд.). Брукс / Коул. стр.205–206. ISBN 978-0-03-025982-1.CS1 maint: несколько имен: список авторов (ссылка на сайт)
- ^ а б Бартоломей, Т. (февраль 2001 г.). «Ультраструктура и формирование выстилки полости тела у Phoronis muelleri (Phoronida, Lophophorata)». Зооморфология. 120 (3): 135–148. Дои:10.1007 / s004350000030. S2CID 23516505.
- ^ Темерева, Елена; Щербакова, Татьяна; Цетлин, Александр (2020). «Первые данные о строении трубок, образованных форонидами». Зоология. 143: 125849. Дои:10.1016 / j.zool.2020.125849. PMID 33099239.
- ^ «Структуры жизни». Национальный институт общих медицинских наук. Получено 2011-03-03.
- ^ Маргулис, Линн; Карлин В. Шварц (1998). «Форонида». Пять королевств: иллюстрированный справочник по типам жизни на Земле (3-е изд.). Эльзевир. стр.340–341. ISBN 978-0-7167-3027-9. Получено 2011-03-27.
- ^ Рупперт, Э.Е., Фокс, Р.С., и Барнс, Р.Д. (2004). «Введение в Bilateria». Зоология беспозвоночных (7-е изд.). Брукс / Коул. стр.212–213. ISBN 978-0-03-025982-1.CS1 maint: несколько имен: список авторов (ссылка на сайт)
- ^ «Гонада - Определение». Бесплатный словарь Merriam-Webster. Merriam-Webster, Incorporated. Получено 2011-03-03.
- ^ Ринкевич, Барух; Валерия Матранга (2009). «Стволовые клетки при бесполом размножении морских беспозвоночных». Стволовые клетки морских организмов. Springer. п. 123. ISBN 978-90-481-2766-5. Получено 2011-03-29.
- ^ «Гермафродит - Определение». Бесплатный словарь Merriam-Webster. Merriam-Webster, Incorporated. Получено 2011-03-03.
- ^ «Перекрестное оплодотворение - определение». Бесплатный словарь Merriam-Webster. Merriam-Webster, Incorporated. Получено 2011-03-10.
- ^ «Двудомный - Определение». Бесплатный словарь Merriam-Webster. Merriam-Webster, Incorporated. Получено 2011-03-03.
- ^ «Гамета - Определение». Бесплатный словарь Merriam-Webster. Merriam-Webster, Incorporated. Получено 2011-03-03.
- ^ Бейли-Брок, Джули Х .; Кристиан К. Эмиг (2000). «Гавайские Phoronida (Lophophorata) и их распространение в Тихоокеанском регионе» (PDF). Тихоокеанская наука. 54 (2): 119–126. Получено 2011-03-11.
- ^ а б Эмиг, C.C. (1982). J.H.S. Blaxter (ред.). Достижения в морской биологии. Академическая пресса. С. 22–23. ISBN 978-0-12-026119-2. Получено 2011-03-12.
- ^ Темерева, Е.Н .; Малахов В.В .; Яковис Э.Л .; Фокин М.В. (Сентябрь – октябрь 2000 г.). "Phoronis ovalis (Phoronida, Lophophorata) в Белом море: первое открытие форонид в Арктическом бассейне ». Доклады биологических наук. 374: 523–525. PMID 11103334.
- ^ а б Темерева, Е. (2009). «Новые данные о распространении, морфологии и систематике личинок форонид (Lophophorata: Phoronida)» (PDF). Зоология беспозвоночных. 6 (1): 47–48. Дои:10.15298 / invertzool.06.1.05. Получено 2011-03-11.
- ^ Стампар, Серджио; Кристиан К. Эмиг; Андре К. Морандини; Гильерме Коджа; Ана Паула Бальбони; Фабио Ланг да Силвейра (2010). "Есть ли опасность в симбиотических видах, связанных с исчезающими видами? Случай, когда форонидный червь растет на Ceriantheomorphe трубка" (PDF). Да. Биол. Мар. 51: 205–211. Архивировано из оригинал (PDF) на 2010-11-12. Получено 2011-03-02.
- ^ Ларсон, Эми А .; Джон Дж. Стахович (2008). «Химическая защита мягкоосадочного форонида от местных эпибентосных хищников». Серия "Прогресс морской экологии". 374: 101–111. Дои:10.3354 / meps07767. ISSN 0171-8630.
- ^ а б Taylor, P.D .; Уилсон, М.А. (2008). "Морфология и родство гедереллоидных" мшанок."" (PDF). В Hageman, S.J .; Ки, М. Jr .; Уинстон, Дж. Э. (ред.). Bryozoan Studies 2007: Материалы 14-й Международной конференции по мшанкам. Музей естественной истории Вирджинии. С. 301–309. Архивировано из оригинал (PDF) on 2010-03-26. Получено 2011-03-26.
- ^ Chen, J.Y .; Г. Чжоу. «Биология фауны Чэнцзян». Бюллетень Национального музея естествознания, Тайбэй. 10: 11–105. - цитируется Эмигом (март 2010 г.) и Тейлор, Винн и Уилсон (2010 г.).
- ^ Чен, Дж. (2004). Рассвет животного мира (на китайском языке). Нанкин: Jiangsu Science and Technology Press. п. 366. - цитируется по Taylor, Vinn & Wilson (2010).
- ^ Конвей Моррис, С. (29 июня 2006 г.). "Дилемма Дарвина: реалии кембрийского взрыва"'". Философские труды Королевского общества B: биологические науки. 361 (1470): 1069–1083. Дои:10.1098 / rstb.2006.1846. ЧВК 1578734. PMID 16754615.
- ^ Бальтазар, Уве; Николас Дж. Баттерфилд (2009). «Раннекембрийские» мягкоствольные «брахиоподы как возможные форониды стеблевой группы» (PDF). Acta Palaeontologica Polonica. 54 (2): 307–314. Дои:10.4202 / приложение.2008.0042. S2CID 54072910. Получено 2011-03-08.
- ^ Skovsted, C.B .; Brock, G.A .; Paterson, J. R .; Holmer, L.E .; Бадд, Г. Э. (2008). "Склеритом Эксцентротека из нижнего кембрия Южной Австралии: сходство с лофофоратами и значение для филогении томмотиидов ». Геология. 36 (2): 171–174. Дои:10.1130 / g24385a.1.
- ^ Skovsted, C.B .; Брок; Topper, T. P .; Paterson, J. R .; Холмер, Л. Э. (2011). «Построение склеритом, биофации, биостратиграфия и систематика томмотиидов. Эксцентротека Helenia sp. ноя из раннего кембрия Южной Австралии ». Палеонтология. 54 (2): 253–286. Дои:10.1111 / j.1475-4983.2010.01031.x.
- ^ Бромли, Р. (2004). «Стратиграфия морской биоэрозии». В Д. Макилроя (ред.). Применение ихнологии к палеоэкологическому и стратиграфическому анализу. Геологическое общество. п. 461. ISBN 978-1-86239-154-3. Получено 2011-03-12.
- ^ Эмиг, Кристиан К. (март 2010 г.). "Ископаемые Phoronida и предполагаемые ихнотаксы" (PDF). Carnets de Géologie. Дои:10.4267/2042/35531. Получено 2011-03-04.
- ^ «Введение в Deuterostomia». Музей палеонтологии Калифорнийского университета. Получено 2010-03-08.
- ^ Нильсен, К. (июль 2002 г.). «Филогенетическое положение Entoprocta, Ectoprocta, Phoronida и Brachiopoda». Интегративная и сравнительная биология. 42 (3): 685–691. Дои:10.1093 / icb / 42.3.685. PMID 21708765. S2CID 15883626.
- ^ Cameron, C.B .; Garey, J.R .; Свалла, Б.Дж. (25 апреля 2000 г.). «Эволюция хордовой формы тела: новые выводы из филогенетического анализа типа deuterostome». Труды Национальной академии наук. 97 (9): 4469–4474. Дои:10.1073 / пнас.97.9.4469. ЧВК 18258. PMID 10781046.
- ^ Галаныч, Кеннет М. (февраль 1996 г.). «Конвергенция в аппаратах питания лофофоратов и птеробранховых полухордовых, выявленная 18S рДНК: интерпретация». Биологический бюллетень. 190 (1): 1–5. Дои:10.2307/1542669. JSTOR 1542669. PMID 29244547. S2CID 44165536.
- ^ Struck, T.H .; Schult, N .; Кусен, Т .; Hickman, E .; Блейдорн. C .; McHugh, D .; Галаныч, К. (Апрель 2007 г.). «Филогения аннелид и статус Sipuncula и Echiura». BMC Эволюционная биология. 7 (57): 57. Дои:10.1186/1471-2148-7-57. ЧВК 1855331. PMID 17411434.
- ^ Коэн, Бернард Л. (февраль 2000 г.). «Монофилия брахиопод и форонид: согласование молекулярных свидетельств с классификацией Линнея (подтип Phoroniformea nov.)». Труды Королевского общества B. 267 (1440): 225–231. Дои:10.1098 / rspb.2000.0991. ЧВК 1690528. PMID 10714876.
- ^ а б Гирибет, Гонсало (апрель 2008 г.). «Сборка лофотрохозойского (спиралевидного) древа жизни». Труды Королевского общества B. 363 (1496): 1513–1522. Дои:10.1098 / rstb.2007.2241. ЧВК 2614230. PMID 18192183.
- ^ Гарей, Джеймс Р. (июль 2002 г.). «Малоизвестные таксоны протостома: введение и дань уважения Роберту П. Хиггинсу». Интегративная и сравнительная биология. 42 (3): 611–618. Дои:10.1093 / icb / 42.3.611. PMID 21708757. S2CID 2117991.
- ^ а б Коэн, Бернард Л .; Агата Вейдман (1 декабря 2005 г.). «Молекулярное свидетельство того, что форониды являются субтаксоном брахиопод (Brachiopoda: Phoronata) и что генетическая дивергенция типов многоклеточных животных началась задолго до раннего кембрия» (PDF). Организмы, разнообразие и эволюция. 5 (4): 253–273. Дои:10.1016 / j.ode.2004.12.002. ISSN 1439-6092. Получено 2011-03-15.
- ^ де Роса, Рено (ноябрь – декабрь 2001 г.). «Молекулярные данные указывают на сходство с протостомами брахиопод». Систематическая биология. 50 (6): 848–859. Дои:10.1080/106351501753462830. PMID 12116636.
- ^ Темерева, Е.Н .; В.В. Малахов (1999). "Новый вид форонид, обитающих в скалах, Phoronis svetlanae (Lophophorata, Phoronida) из Японского моря ». Зоологический журнал (по-русски). 78 (5): 626–630. ISSN 0044-5134. Получено 2011-03-31.
- ^ Темерева, Е. (2000). "Новые виды форонид Phoronopsis malakhovi (Lophophorata, Phoronida) из Южно-Китайского моря ». Зоологический журнал (по-русски). 79 (9): 1088–1093.
- ^ Сантагата, Скотт (2009). «Форида Мексиканского залива». У Дэррила Л. Фелдера; Сильвия А. Эрл (ред.). Происхождение, воды и биота Мексиканского залива: биоразнообразие. Издательство Техасского университета A&M. С. 1133–1134. ISBN 978-1-60344-094-3. Получено 2011-03-29.