Антифриз протеин - Antifreeze protein
| Протеин антифриза насекомых, Тенебрио-тип | |||||||||
|---|---|---|---|---|---|---|---|---|---|
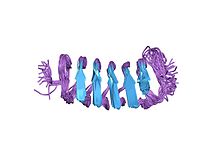 Структура Тенебрио Молитор бета-спиральный антифризный белок[1] | |||||||||
| Идентификаторы | |||||||||
| Символ | AFP | ||||||||
| Pfam | PF02420 | ||||||||
| ИнтерПро | IPR003460 | ||||||||
| SCOP2 | 1ezg / Объем / СУПФАМ | ||||||||
| |||||||||
| Антифризный белок насекомых (CfAFP) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Структура Choristoneura fumiferana (червь еловая) бета-спиральный антифризный белок[2] | |||||||||
| Идентификаторы | |||||||||
| Символ | CfAFP | ||||||||
| Pfam | PF05264 | ||||||||
| ИнтерПро | IPR007928 | ||||||||
| SCOP2 | 1м8н / Объем / СУПФАМ | ||||||||
| |||||||||
| Рыбный антифриз протеин, тип I | |
|---|---|
| Идентификаторы | |
| Символ | ? |
| ИнтерПро | IPR000104 |
| SCOP2 | 1wfb / Объем / СУПФАМ |
| Рыбный антифриз протеин, тип II | |
|---|---|
| Идентификаторы | |
| Символ | ? |
| ИнтерПро | IPR002353 |
| CATH | 2py2 |
| SCOP2 | 2afp / Объем / СУПФАМ |
| Рыбный антифриз протеин III типа | |
|---|---|
| Идентификаторы | |
| Символ | ? |
| ИнтерПро | IPR006013 |
| SCOP2 | 1hg7 / Объем / СУПФАМ |
| Также домен SAF (ИнтерПро: IPR013974 ). | |
| Связующий со льдом протеиноподобный (организм морского льда) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||
| Символ | DUF3494 | ||||||||
| Pfam | PF11999 | ||||||||
| ИнтерПро | IPR021884 | ||||||||
| |||||||||
Антифризы протеины (AFP) или же белки, структурирующие лед (Интернет-провайдеры) относятся к классу полипептиды произведено некоторыми животные, растения, грибы и бактерии которые позволяют им выжить при температурах ниже точки замерзания воды. AFP связываются с небольшими кристаллы льда подавлять рост и перекристаллизация льда, который иначе был бы фатальным.[3] Также появляется все больше доказательств того, что AFP взаимодействуют с мембранами клеток млекопитающих, чтобы защитить их от повреждения холодом. Эта работа предполагает участие ОВП при простуде. акклиматизация.[4]
Неколлигативные свойства
В отличие от широко применяемого автомобильного антифриза, этиленгликоль, AFP не понижают точку замерзания пропорционально концентрации.[нужна цитата ] Скорее, они работают безколлигативный манера. Это явление позволяет им действовать как антифриз при концентрациях от 1/300 до 1/500 от концентраций других растворенных веществ. Их низкая концентрация сводит к минимуму их влияние на осмотическое давление.[4] Необычные свойства AFP объясняются их избирательным сродством к определенным кристаллическим формам льда и, как следствие, блокадой процесса образования льда.[5]
Температурный гистерезис
AFP создают разницу между температурой плавления и замерзания (температура разрушения кристаллов льда, связанных AFP), известную как тепловой гистерезис. Добавление AFP на границе раздела между твердым льдом и жидкой водой подавляет термодинамически благоприятный рост кристаллов льда. Рост льда кинетически ингибируется AFP, покрывающими доступные для воды поверхности льда.[5]
Температурный гистерезис легко измерить в лаборатории с помощью нанолитровый осмометр. Организмы различаются по величине теплового гистерезиса. Максимальный уровень теплового гистерезиса, показанный AFP рыб, составляет приблизительно -3,5 ° C (Sheikh Mahatabuddin et al., SciRep) (29,3 ° F). Однако белки-антифризы из насекомых в 10–30 раз активнее рыбных белков. Эта разница, вероятно, отражает более низкие температуры, с которыми сталкиваются насекомые на суше. Напротив, водные организмы подвергаются воздействию только от -1 до -2 ° C ниже нуля. В экстремальные зимние месяцы еловый почковый червь устойчив к замерзанию при температуре около −30 ° C.[4] Аляскинский жук Upis ceramboides могут выжить при температуре -60 ° C, используя антифризы, не являющиеся белками.[6]
Скорость охлаждения может влиять на значение теплового гистерезиса AFP. Быстрое охлаждение может существенно снизить температуру неравновесного замерзания и, следовательно, величину теплового гистерезиса. Следовательно, организмы не обязательно могут адаптироваться к своей отрицательной среде, если температура резко падает.[4]
Устойчивость к замерзанию или предотвращение замерзания
Виды, содержащие AFP, могут быть классифицированы как
Избегающий замораживания: Эти виды способны полностью предотвратить замерзание жидкостей своего тела. Как правило, функция AFP может быть преодолена при очень низких температурах, что приводит к быстрому росту льда и смерти.
Морозоустойчивый: Эти виды способны пережить замерзание жидкости организма. Считается, что некоторые устойчивые к замораживанию виды используют AFP в качестве криопротекторов, чтобы предотвратить повреждение замораживания, но не замораживания в целом. Точный механизм пока неизвестен. Однако считается, что AFP могут ингибировать перекристаллизацию и стабилизировать клеточные мембраны, чтобы предотвратить повреждение льдом.[7] Они могут работать вместе с нуклеаторы белкового льда (PIN) для контроля скорости распространения льда после замерзания.[7]
Разнообразие
Есть много известных негомологичный типы ОВП.
Рыба AFPs

Антифриз гликопротеины или AFGP находятся в Антарктика нототениоиды и северная треска. Они составляют 2,6–3,3 кДа.[8] AFGP развивались отдельно у нототениоидов и северной трески. У нототениоидов ген AFGP произошел от предкового гена трипсиноген-подобной сериновой протеазы.[9]
- AFP типа I встречается в зимняя камбала, длиннорогий бычок и шорторог. Это лучше всего документированный AFP, потому что он был первым, в котором была определена его трехмерная структура.[10] АФП I типа состоит из одной длинной амфипатической альфа-спирали размером около 3,3-4,5 кДа. У трехмерной структуры есть три грани: гидрофобная, гидрофильная и грань Thr-Asx.[10]
- Тип I-hyp АФП (где hyp означает гиперактивность) обнаруживается у нескольких камбал с правым глазом. Это приблизительно 32 кДа (две димерные молекулы 17 кДа). Белок выделен из плазмы крови озимой камбалы. Он значительно лучше выдерживает низкие температуры замерзания, чем большинство AFP для рыбы.[11] Эта способность частично проистекает из его многочисленных повторов сайта связывания льда типа I.[12]
- ОВП типа II (например, P05140) находятся в морской ворон, нюхать и сельдь. Это богатые цистеином глобулярные белки, содержащие пять дисульфидные связи.[13] AFP типа II, вероятно, произошли от кальций-зависимых (c-типа) лектинов.[14] Морские вороны, корюшка и сельдь - довольно разные линии происхождения костистость. Если ген AFP присутствовал в самом последнем общем предке этих линий, то необычно, что ген разбросан по этим линиям, присутствует в одних порядках и отсутствует в других. Было высказано предположение, что латеральный перенос гена может быть объяснен этим несоответствием, так что корюшка приобрела ген AFP типа II от сельди.[15]
- ОВП типа III обнаружены в Антарктике. бельдюга. Они обладают такой же общей гидрофобностью на поверхностях, связывающих лед, что и AFP типа I. Они имеют размер примерно 6 кДа.[8] АФП типа III, вероятно, произошли от гена синтазы сиаловой кислоты (SAS), присутствующего у антарктического бельдюга. В результате дупликации гена этот ген, который, как было показано, проявляет некоторую собственную активность по связыванию льда, превратился в эффективный ген AFP за счет потери N-концевой части.[16]
- ОВП типа IV (P80961) встречаются у длиннорогих бычков. Это альфа-спиральные белки, богатые глутаматом и глутамином.[17] Этот белок имеет размер примерно 12 кДа и состоит из 4-спирального пучка.[17] Его единственная посттрансляционная модификация - это пироглутамат остаток, циклизованный глутамин остаток на его N-конец.[17]
Завод AFP
Классификация АФП стала более сложной, когда были открыты антифризы из растений.[18] Растительные AFP значительно отличаются от других AFP по следующим аспектам:
- По сравнению с другими AFP, они обладают гораздо более слабой активностью теплового гистерезиса.[19]
- Их физиологическая функция, вероятно, заключается в подавлении перекристаллизации льда, а не в предотвращении образования льда.[19]
- Большинство из них эволюционировали патогенез -связанные белки, иногда сохраняющие противогрибковый характеристики.[19]
AFP насекомых
У насекомых обнаружен ряд AFP, в том числе из Дендроиды, Тенебрио и Рагиум жуки, еловые черви и бледные мотыльки, а также мошки (в том же порядке, что и мухи). AFP насекомых имеют определенное сходство, при этом большинство из них обладают более высокой активностью (то есть большим значением теплового гистерезиса, называемым гиперактивным) и повторяющейся структурой с плоской поверхностью, связывающей лед. Те из близкородственных Тенебрио и Дендроиды Жуки гомологичны, и каждый 12–13-аминокислотный повтор стабилизирован внутренней дисульфидной связью. Изоформы имеют от 6 до 10 таких повторов, которые образуют катушку или бета-соленоид. Одна сторона соленоида имеет плоскую поверхность, связывающую лед, которая состоит из двойного ряда остатков треонина.[7] [20] Другие жуки (род Рагиум) имеют более длинные повторы без внутренних дисульфидных связей, которые образуют сжатый бета-соленоид (бета-сэндвич) с четырьмя рядами остатков треонина,[21] и этот AFP структурно аналогичен моделированному негомологичному AFP из бледной красавицы.[22] Напротив, AFP еловой почковой совки представляет собой соленоид, внешне напоминающий Тенебрио белок, с аналогичной поверхностью связывания со льдом, но имеет треугольное поперечное сечение с более длинными повторами, в которых отсутствуют внутренние дисульфидные связи. АФП от мошек структурно сходен с таковыми от мошек. Тенебрио и Дендроиды, но бета-соленоид с дисульфидной связкой образован из более коротких 10 аминокислотных повторов, и вместо треонина связывающая лед поверхность состоит из одного ряда остатков тирозина.[23] Коллемболы (Collembola) не насекомые, но, как и насекомые, это шестиногие членистоногие. Вид, обитающий в Канаде, которого часто называют «снежной блохой», производит гиперактивные AFP.[24] Хотя они также повторяются и имеют плоскую ледяную поверхность, на этом сходство заканчивается. Около 50% остатков представляют собой глицин (Gly) с повторами Gly-Gly-X или Gly-X-X, где X - любая аминокислота. Каждый 3-аминокислотный повтор образует один виток спирали полипролина II типа. Затем спирали складываются вместе, образуя пучок толщиной в две спирали с поверхностью, связывающей лед, в которой преобладают небольшие гидрофобные остатки, такие как аланин, а не треонин.[25] Другие насекомые, такие как аляскинский жук, производят гиперактивные антифризы, которые даже менее похожи, поскольку они представляют собой полимеры сахаров (ксиломаннан), а не полимеры аминокислот (белков).[26] Взятые вместе, это говорит о том, что большинство AFP и антифризов возникло после того, как линии, давшие начало этим различным насекомым, разошлись. Сходства, которые они разделяют, являются результатом конвергентной эволюции.
ОВП организмов морского льда
Многие микроорганизмы, живущие в морской лед обладают AFP, которые принадлежат одной семье. В диатомеи Fragilariopsis cylindrus и F. curta играют ключевую роль в сообществах полярного морского льда, доминируя в сообществах как пластинчатого слоя, так и пакового льда. AFP широко распространены у этих видов, и наличие AFP гены как мультигенное семейство указывает на важность этой группы для рода Фрагилариопсис.[27] ОВП, выявленные в F. cylindrus принадлежат к семейству AFP, которое представлено различными таксонами и может быть обнаружено у других организмов, связанных с морским льдом (Колвеллия виды, Navicula glaciei, Chaetoceros neogracile и Stephos longipes и Leucosporidium antarcticum)[28][29] и антарктические внутренние ледяные бактерии (Флавобактерии ),[30][31] а также у морозоустойчивых грибов (Typhula ishikariensis, Lentinula edodes и Flammulina populicola ).[32][33]
Решено несколько структур для AFP морского льда. Это семейство белков сворачивается в бета-спираль которые образуют плоскую ледяную поверхность.[34] В отличие от других AFP, для сайта связывания со льдом нет единого мотива последовательности.[35]
AFP, обнаруженный из метагенома инфузория Euplotes focardii а психрофильные бактерии обладают эффективной способностью ингибировать перекристаллизацию льда.[36] 1 мкМ Euplotes focardii белок консорциума, связывающий лед (EFCIBP) достаточно для полного подавления перекристаллизации льда при температуре –7,4 ° C. Эта способность ингибировать перекристаллизацию льда помогает бактериям переносить лед, а не предотвращает образование льда. EFCIBP также создает зазор теплового гистерезиса, но эта способность не так эффективна, как способность ингибирования рекристаллизации льда. EFCIBP помогает защитить как очищенные белки, так и целые бактериальные клетки при отрицательных температурах. Зеленый флуоресцентный белок работает после нескольких циклов замораживания и плавления при инкубации с EFCIBP. кишечная палочка выдерживает более длительные периоды при температуре 0 ° C, когда efcIBP ген был вставлен в Кишечная палочка геном.[36] EFCIBP имеет типичную структуру AFP, состоящую из нескольких бета-листы и альфа-спираль. Кроме того, все полярные остатки, связывающиеся со льдом, находятся в одном и том же сайте белка. [36]
Эволюция
Замечательное разнообразие и распределение AFP предполагают, что различные типы AFP эволюционировали недавно в ответ на изменение уровня моря. оледенение произошедшие 1-2 миллиона лет назад в Северном полушарии и 10-30 миллионов лет назад в Антарктиде. Это независимое развитие подобных приспособлений называется конвергентная эволюция.[4] Есть две причины, по которым многие типы AFP могут выполнять одну и ту же функцию, несмотря на их разнообразие:
- Хотя лед однородно состоит из молекул воды, он имеет множество различных поверхностей, подверженных связыванию. Различные типы AFP могут взаимодействовать с разными поверхностями.
- Хотя пять типов AFP различаются по своему первичная структура аминокислот, когда каждая сворачивается в функционирующий белок, они могут иметь общие черты в их трехмерном или третичная структура что обеспечивает такое же взаимодействие со льдом.[4][37]
Механизмы действия
Считается, что АФП подавляют рост адсорбция –Механизм торможения.[38] Они адсорбируются на небазальные плоскости льда, подавляя термодинамически благоприятный рост льда.[39] Наличие плоской жесткой поверхности в некоторых AFP, по-видимому, способствует его взаимодействию со льдом через Сила Ван-дер-Ваальса комплементарность поверхности.[40]
Привязка ко льду
Обычно кристаллы льда, выращенные в растворе, имеют только базальную (0001) и призматическую (1010) грани и выглядят как круглые и плоские диски.[5] Однако, похоже, присутствие AFP открывает другие лица. Теперь выясняется, что поверхность льда 2021 года является предпочтительной связывающей поверхностью, по крайней мере, для AFP типа I.[41] Первоначально предполагалось, что в ходе исследований AFP типа I лед и AFP взаимодействуют посредством водородных связей (Raymond and DeVries, 1977). Однако, когда части белка, которые, как считается, способствуют образованию водородных связей, были мутированы, предполагаемого снижения активности антифриза не наблюдалось. Последние данные предполагают, что основной причиной могут быть гидрофобные взаимодействия.[42] Трудно определить точный механизм связывания из-за сложной границы раздела вода-лед. В настоящее время попытки раскрыть точный механизм делаются с помощью молекулярное моделирование программы (молекулярная динамика или Метод Монте-Карло ).[3][5]
Механизм связывания и функция антифриза
Согласно исследованию структуры и функций антифриза из Pseudopleuronectes americanus,[43] Было показано, что механизм антифриза молекулы AFP I типа обусловлен связыванием со структурой зародышеобразования льда по типу застежки-молнии посредством водородных связей гидроксильные группы из четырех Thr остатки кислорода вдоль направление в решетке льда, впоследствии останавливая или замедляя рост ледяных пирамидальных плоскостей, чтобы снизить точку замерзания.[43]
![[01 { overline {1}} 2]](https://wikimedia.org/api/rest_v1/media/math/render/svg/25dd6fa8ecf7c3d8135f0b3bda61dde8032d89ce)
Вышеупомянутый механизм может быть использован для выяснения взаимосвязи структура-функция других белков-антифризов со следующими двумя общими особенностями:
- повторение Thr остаток (или любой другой полярный аминокислотный остаток, боковая цепь которого может образовывать водородную связь с водой) в периоде из 11 аминокислот вдоль рассматриваемой последовательности, и
- высокий процент Ала остаточный компонент в нем.[43]
История
В 1950-х годах норвежский ученый Шоландер попытался объяснить, как арктические рыбы могут выжить в воде, температура которой ниже точки замерзания их крови. Его эксперименты привели его к мысли, что в крови арктических рыб есть «антифриз».[3] Затем, в конце 1960-х годов, биолог-зоотехник Артур ДеВрис смог выделить антифризный белок в ходе исследования антарктических рыб.[44] Эти белки позже были названы антифризовыми гликопротеинами (AFGP) или антифризовыми гликопептидами, чтобы отличить их от недавно открытых негликопротеиновых биологических антифризов (AFP). Де Врис работал с Робертом Фини (1970), чтобы охарактеризовать химические и физические свойства антифризовых белков.[45] В 1992 году Гриффит и другие. задокументировали открытие АФП в листьях озимой ржи.[18] Примерно в то же время Уррутия, Думан и Найт (1992) зарегистрировали белок теплового гистерезиса у покрытосеменных.[46] В следующем году Думан и Олсен отметили, что AFP были обнаружены у более чем 23 видов животных. покрытосеменные, в том числе съеденные людьми.[47] Они также сообщили об их присутствии в грибах и бактериях.
Изменение имени
Недавно были предприняты попытки переименовать белки антифриза как белки, структурирующие лед, чтобы более точно представить их функцию и устранить любую предполагаемую отрицательную связь между AFP и автомобильным антифризом. этиленгликоль. Эти две вещи представляют собой совершенно разные сущности и слабо похожи только по своим функциям.[48]
Коммерческие и медицинские приложения
Многие области могут получить выгоду от защиты от повреждения тканей замораживанием. В настоящее время компании изучают возможность использования этих белков в:[нужна цитата ]
- Повышение морозостойкости сельскохозяйственных культур и продление сезона сбора урожая в более прохладном климате
- Улучшение производства рыбы на фермах в более прохладном климате
- Увеличение срока хранения замороженных продуктов
- Улучшение криохирургия
- Повышение сохранности тканей для трансплантации или переливания в медицине[24]
- Терапия от переохлаждения
- Криоконсервация человека (крионика)
Unilever получил одобрение Великобритании на использование генетически модифицированный дрожжи для производства белков-антифризов из рыбы, для использования в производстве мороженого.[49]
Свежие новости
Одним из недавних успешных бизнес-начинаний стало внедрение AFP в мороженое и йогуртовые продукты. Этот ингредиент, обозначенный как белок, структурирующий лед, был одобрен Управление по контролю за продуктами и лекарствами. Белки выделяются из рыбы и реплицируются в большем масштабе в генетически модифицированных дрожжах.[50]
Есть опасения со стороны организаций, выступающих против генетически модифицированные организмы (ГМО), которые считают, что антифризы могут вызывать воспаление.[51] Потребление AFP с пищей, вероятно, уже достаточно в большинстве северных и умеренных регионов.[8] Учитывая известное историческое потребление AFP, можно с уверенностью заключить, что их функциональные свойства не передают никаких токсикологических или аллергенный эффекты у людей.[8]
Также трансгенный процесс производства ISP уже широко используется в обществе. Инсулин и сычужный фермент производятся по этой технологии. Процесс не влияет на продукт; это просто делает производство более эффективным и предотвращает гибель рыбы, которая в противном случае была бы убита для извлечения белка.
В настоящее время, Unilever включает AFP в некоторые из своих американских продуктов, в том числе в некоторые Эскимо ледяные хлопки и новая линия Брейерс Легкие батончики для мороженого с двойным взбиванием. В мороженом AFP позволяют производить очень сливочное, плотное мороженое с пониженным содержанием жира с меньшим количеством добавок.[52] Они контролируют рост кристаллов льда, вызванный таянием на погрузочной платформе или кухонном столе, что снижает качество текстуры.[53]
В ноябре 2009 г. Труды Национальной академии наук опубликовал открытие молекулы у аляскинского жука, которая ведет себя как AFP, но состоит из сахариды и жирные кислоты.[6]
Исследование 2010 года продемонстрировало стабильность кристаллов перегретого водяного льда в растворе AFP, показав, что, хотя белки могут препятствовать замораживанию, они также могут препятствовать таянию.[54]
Рекомендации
- ^ Дейли М.Э., Спайракопулос Л., Джиа З., Дэвис П.Л., Сайкс Б.Д. (апрель 2002 г.). «Структура и динамика бета-спирального антифриза». Биохимия. 41 (17): 5515–25. Дои:10.1021 / bi0121252. PMID 11969412.
- ^ Лейнала Е.К., Дэвис П.Л., Дусе Д., Тышенко М.Г., Уокер В.К., Цзя З. (сентябрь 2002 г.). «Бета-спиральная изоформа белка антифриза с повышенной активностью. Структурные и функциональные идеи». Журнал биологической химии. 277 (36): 33349–52. Дои:10.1074 / jbc.M205575200. PMID 12105229.
- ^ а б c Гудселл Д. (декабрь 2009 г.). «Молекула месяца: антифризы». Исследовательский институт Скриппса и RCSB PDB. Дои:10.2210 / rcsb_pdb / mom_2009_12.
- ^ а б c d е ж Флетчер Г.Л., Хью С.Л., Дэвис П.Л. (2001). «Антифризные протеины костистых рыб». Ежегодный обзор физиологии. 63: 359–90. Дои:10.1146 / annurev.physiol.63.1.359. PMID 11181960.
- ^ а б c d Жоров А, Жоров Б.С., Ян Д.С. (июнь 2004 г.). «Теоретическое исследование взаимодействия протеина антифриза озимой камбалы со льдом». Белковая наука. 13 (6): 1524–37. Дои:10.1110 / пс. 04641104. ЧВК 2279984. PMID 15152087.
- ^ а б Уолтерс К.Р., Серианни А.С., Сформо Т, Барнс Б.М., Думан Дж.Г. (декабрь 2009 г.). «Небелковый антифриз ксиломаннана, продуцирующий термический гистерезис, у морозостойкого аляскинского жука Upis ceramboides». Труды Национальной академии наук Соединенных Штатов Америки. 106 (48): 20210–5. Bibcode:2009PNAS..10620210W. Дои:10.1073 / pnas.0909872106. ЧВК 2787118. PMID 19934038.
- ^ а б c Думан Дж. Г. (2001). «Антифриз и белки-нуклеаторы льда у наземных членистоногих». Ежегодный обзор физиологии. 63: 327–57. Дои:10.1146 / annurev.physiol.63.1.327. PMID 11181959.
- ^ а б c d Кревель Р.В., Федык Дж.К., Сперджен М.Дж. (июль 2002 г.). «Антифризные протеины: характеристики, возникновение и воздействие на человека». Пищевая и химическая токсикология. 40 (7): 899–903. Дои:10.1016 / S0278-6915 (02) 00042-X. PMID 12065210.
- ^ Chen L, DeVries AL, Cheng CH (апрель 1997 г.). «Эволюция гена гликопротеина антифриза из гена трипсиногена у антарктических нототениоидных рыб». Труды Национальной академии наук Соединенных Штатов Америки. 94 (8): 3811–6. Bibcode:1997PNAS ... 94.3811C. Дои:10.1073 / пнас.94.8.3811. ЧВК 20523. PMID 9108060.
- ^ а б Думан Дж. Г., де Фрис А. Л. (1976). «Выделение, характеристика и физические свойства протеиновых антифризов из озимой камбалы Pseudopleuronectes americanus». Сравнительная биохимия и физиология. B, Сравнительная биохимия. 54 (3): 375–80. Дои:10.1016/0305-0491(76)90260-1. PMID 1277804.
- ^ Скоттер А.Дж., Маршалл С.Б., Грэм Л.А., Гилберт Дж.А., Гарнхэм С.П., Дэвис П.Л. (октябрь 2006 г.). «Основа гиперактивности антифризовых белков». Криобиология. 53 (2): 229–39. Дои:10.1016 / j.cryobiol.2006.06.006. PMID 16887111.
- ^ Грэм Л.А., Маршалл С.Б., Лин Ф.Х., Кэмпбелл Р.Л., Дэвис П.Л. (февраль 2008 г.). «Гиперактивный протеин-антифриз из рыбы содержит множество участков связывания льда». Биохимия. 47 (7): 2051–63. Дои:10.1021 / bi7020316. PMID 18225917.
- ^ Ng NF, Hew CL (август 1992 г.). «Структура полипептида антифриза морского ворона. Дисульфидные связи и сходство с лектинсвязывающими белками». Журнал биологической химии. 267 (23): 16069–75. PMID 1644794.
- ^ Юарт К.В., Рубинский Б., Флетчер Г.Л. (май 1992 г.). «Структурное и функциональное сходство между белками антифризов рыб и кальций-зависимыми лектинами». Сообщения о биохимических и биофизических исследованиях. 185 (1): 335–40. Дои:10.1016 / s0006-291x (05) 90005-3. PMID 1599470.
- ^ Грэм Л.А., Лугид СК, Юарт К.В., Дэвис П.Л. (июль 2008 г.). «Боковой перенос гена лектин-подобного антифриза у рыб». PLOS ONE. 3 (7): e2616. Bibcode:2008PLoSO ... 3,26 16G. Дои:10.1371 / journal.pone.0002616. ЧВК 2440524. PMID 18612417.

- ^ Келли Дж. Л., Агард Дж. Э., Маккосс М. Дж., Суонсон В. Дж. (Август 2010 г.). «Функциональная диверсификация и эволюция белков-антифризов у антарктической рыбы Lycodichthys dearborni». Журнал молекулярной эволюции. 71 (2): 111–8. Bibcode:2010JMolE..71..111K. Дои:10.1007 / s00239-010-9367-6. PMID 20686757. S2CID 25737518.
- ^ а б c Дэн Г., Эндрюс Д.В., Лаурсен Р.А. (январь 1997 г.). «Аминокислотная последовательность антифриза нового типа, из длиннорогого быка Myoxocephalus octodecimspinosis». Письма FEBS. 402 (1): 17–20. Дои:10.1016 / S0014-5793 (96) 01466-4. PMID 9013849.
- ^ а б Гриффит М., Ала П., Ян Д.С., Хон В.К., Моффатт Б.А. (октябрь 1992 г.). «Антифризный белок, вырабатываемый эндогенно в листьях озимой ржи». Физиология растений. 100 (2): 593–6. Дои:10.1104 / стр.100.2.593. ЧВК 1075599. PMID 16653033.
- ^ а б c Гриффит М., Яиш М.В. (август 2004 г.). «Белки-антифризы у зимующих растений: рассказ о двух действиях». Тенденции в растениеводстве. 9 (8): 399–405. Дои:10.1016 / j.tplants.2004.06.007. PMID 15358271.
- ^ Лиу YC, Tocilj A, Davies PL, Jia Z (июль 2000 г.). «Мимикрия структуры льда поверхностными гидроксилами и водой антифриза бета-спирали». Природа. 406 (6793): 322–4. Bibcode:2000Натурал.406..322л. Дои:10.1038/35018604. PMID 10917536. S2CID 4385352.
- ^ Hakim A, Nguyen JB, Basu K, Zhu DF, Thakral D, Davies PL, et al. (Апрель 2013). «Кристаллическая структура белка антифриза насекомых и ее значение для связывания льда». Журнал биологической химии. 288 (17): 12295–304. Дои:10.1074 / jbc.M113.450973. ЧВК 3636913. PMID 23486477.
- ^ Лин Ф. Х., Дэвис П. Л., Грэм Л. А. (май 2011 г.). «Гиперактивный антифриз, богатый Thr- и Ala, извлекается из складок дюймового червя в виде плоской шелкоподобной β-спирали». Биохимия. 50 (21): 4467–78. Дои:10.1021 / bi2003108. PMID 21486083.
- ^ Басу К., Вассерман СС, Джеронимо П.С., Грэм Л.А., Дэвис П.Л. (апрель 2016 г.). «Промежуточная активность антифриза от мошек обусловлена богатым тирозином сайтом связывания со льдом и атипичным сродством с ледяной плоскостью». Журнал FEBS. 283 (8): 1504–15. Дои:10.1111 / febs.13687. PMID 26896764. S2CID 37207016.
- ^ а б Грэм Л.А., Дэвис П.Л. (октябрь 2005 г.). «Антифризные протеины от снежных блох, богатые глицином». Наука. 310 (5747): 461. Дои:10.1126 / science.1115145. PMID 16239469. S2CID 30032276. Сложить резюме – Science Daily.
- ^ Пентелут Б.Л., Гейтс З.П., Терешко В., Дашнау Ю.Л., Вандеркой Ю.М., Коссяков А.А., Кент С.Б. (июль 2008 г.). «Рентгеновская структура белка антифриза снежных блох, определенная рацемической кристаллизацией синтетических белковых энантиомеров». Журнал Американского химического общества. 130 (30): 9695–701. Дои:10.1021 / ja8013538. ЧВК 2719301. PMID 18598029.
- ^ Уолтерс К.Р., Серианни А.С., Сформо Т, Барнс Б.М., Думан Дж.Г. (декабрь 2009 г.). «Небелковый антифриз ксиломаннана, вызывающий термический гистерезис, у морозоустойчивого аляскинского жука Upis ceramboides». Труды Национальной академии наук Соединенных Штатов Америки. 106 (48): 20210–5. Дои:10.1073 / pnas.0909872106. PMID 19934038. S2CID 25741145.
- ^ Bayer-Giraldi M, Uhlig C, John U, Mock T, Valentin K (апрель 2010 г.). «Белки-антифризы в диатомовых водорослях полярного морского льда: разнообразие и экспрессия генов в роде Fragilariopsis». Экологическая микробиология. 12 (4): 1041–52. Дои:10.1111 / j.1462-2920.2009.02149.x. PMID 20105220.
- ^ Раймонд Дж. А., Фритсен С., Шен К. (август 2007 г.). «Связывающий лед белок из бактерии антарктического морского льда». FEMS Microbiology Ecology. 61 (2): 214–21. Дои:10.1111 / j.1574-6941.2007.00345.x. PMID 17651136.
- ^ Кико Р. (апрель 2010 г.). «Приобретение защиты от замерзания у ракообразных морского льда посредством горизонтального переноса генов?». Полярная биология. 33 (4): 543–56. Дои:10.1007 / s00300-009-0732-0. S2CID 20952951.
- ^ Раймонд Дж. А., Кристнер Британская Колумбия, Шустер СК (сентябрь 2008 г.). «Бактериальный связывающий лед белок из ледяного керна Востока». Экстремофилов. 12 (5): 713–7. Дои:10.1007 / s00792-008-0178-2. PMID 18622572. S2CID 505953.
- ^ Нан Сяо, Сигеки Инаба, Мотоаки Тодзё, Йосуке Дегава, Сэйити Фуджиу, Сакаэ Кудо, Тамоцу Хосино (22 декабря 2010 г.). «Антифризная активность различных грибов и Stramenopila, выделенных из Антарктиды». Североамериканские грибы. 5: 215–220. Дои:10.2509 / наф2010.005.00514.CS1 maint: использует параметр авторов (связь)
- ^ Хосино Т., Кириаки М., Охия С., Фудзивара М., Кондо Х, Нисимия Ю., Юмото И., Цуда С. (декабрь 2003 г.). «Протеины-антифризы от грибков снежной плесени». Канадский журнал ботаники. 81 (12): 1175–81. Дои:10.1139 / b03-116.
- ^ Раймонд JA, Janech MG (апрель 2009 г.).«Связывающие лед протеины из грибов эноки и шиитаке». Криобиология. 58 (2): 151–6. Дои:10.1016 / j.cryobiol.2008.11.009. PMID 19121299.
- ^ Ханада Ю., Нисимия Ю., Миура А., Цуда С., Кондо Х. (август 2014 г.). «Гиперактивный антифризный белок из антарктической ледяной бактерии Colwellia sp. Имеет сложный участок связывания со льдом без повторяющихся последовательностей». Журнал FEBS. 281 (16): 3576–90. Дои:10.1111 / фев.12878. PMID 24938370. S2CID 8388070.
- ^ До Х, Ким СДжей, Ким ХД, Ли ДжХ (апрель 2014 г.). «Характеристика на основе структуры и антифризные свойства гиперактивного связывающего лед протеина из антарктической бактерии Flavobacterium frigoris PS1». Acta Crystallographica. Раздел D, Биологическая кристаллография. 70 (Pt 4): 1061–73. Дои:10.1107 / S1399004714000996. PMID 24699650.
- ^ а б c Манджиагалли М., Бар-Долев М., Тедеско П., Наталелло А., Каледа А., Брокка С. и др. (Январь 2017 г.). «Криозащитный эффект связывающего лед белка, полученного из антарктических бактерий». Журнал FEBS. 284 (1): 163–177. Дои:10.1111 / фев.13965. HDL:11581/397803. PMID 27860412. S2CID 43854468.
- ^ Chen L, DeVries AL, Cheng CH (апрель 1997 г.). «Конвергентная эволюция гликопротеинов антифриза в антарктических нототениоидных рыбах и арктической треске». Труды Национальной академии наук Соединенных Штатов Америки. 94 (8): 3817–22. Bibcode:1997PNAS ... 94.3817C. Дои:10.1073 / пнас.94.8.3817. ЧВК 20524. PMID 9108061.
- ^ Раймонд Дж. А., Де Вриз А. Л. (июнь 1977 г.). «Ингибирование адсорбции как механизм морозостойкости полярных рыб».. Труды Национальной академии наук Соединенных Штатов Америки. 74 (6): 2589–93. Bibcode:1977PNAS ... 74.2589R. Дои:10.1073 / pnas.74.6.2589. ЧВК 432219. PMID 267952.
- ^ Раймонд Дж. А., Уилсон П., Де Вриз А. Л. (февраль 1989 г.). «Торможение роста небазальных плоскостей во льду рыбными антифризами». Труды Национальной академии наук Соединенных Штатов Америки. 86 (3): 881–5. Bibcode:1989ПНАС ... 86..881Р. Дои:10.1073 / pnas.86.3.881. ЧВК 286582. PMID 2915983.
- ^ Ян Д.С., Хон В.К., Бубанко С., Сюэ Ю., Ситхараман Дж., Хью С.Л., Сичери Ф. (май 1998 г.). «Идентификация поверхности связывания льда на протеине антифриза типа III с помощью алгоритма« функции плоскостности »». Биофизический журнал. 74 (5): 2142–51. Bibcode:1998BpJ .... 74.2142Y. Дои:10.1016 / S0006-3495 (98) 77923-8. ЧВК 1299557. PMID 9591641.
- ^ Knight CA, Cheng CC, DeVries AL (февраль 1991 г.). «Адсорбция альфа-спиральных антифризовых пептидов на определенных плоскостях поверхности кристаллов льда». Биофизический журнал. 59 (2): 409–18. Bibcode:1991BpJ .... 59..409K. Дои:10.1016 / S0006-3495 (91) 82234-2. ЧВК 1281157. PMID 2009357.
- ^ Haymet AD, Ward LG, Harding MM, Knight CA (июль 1998 г.). «Валин заменил зимнюю камбалу« антифризом »: сохранение гистерезиса нарастания льда». Письма FEBS. 430 (3): 301–6. Дои:10.1016 / S0014-5793 (98) 00652-8. PMID 9688560. S2CID 42371841.
- ^ а б c Chou KC (январь 1992 г.). «Энергетически оптимизированная структура антифриза и механизм его связывания». Журнал молекулярной биологии. 223 (2): 509–17. Дои:10.1016/0022-2836(92)90666-8. PMID 1738160.
- ^ DeVries AL, Wohlschlag DE (март 1969 г.). «Морозостойкость некоторых антарктических рыб». Наука. 163 (3871): 1073–5. Bibcode:1969Sci ... 163.1073D. Дои:10.1126 / science.163.3871.1073. PMID 5764871. S2CID 42048517.
- ^ ДеВриз А.Л., Комацу СК, Фини Р.Э. (июнь 1970 г.). «Химические и физические свойства гликопротеинов антарктических рыб, снижающих температуру замерзания». Журнал биологической химии. 245 (11): 2901–8. PMID 5488456.
- ^ Уррутия М.Э., Думан Дж. Г., Knight CA (май 1992 г.). «Растительные белки теплового гистерезиса». Biochimica et Biophysica Acta (BBA) - Структура белка и молекулярная энзимология. 1121 (1–2): 199–206. Дои:10.1016 / 0167-4838 (92) 90355-ч. PMID 1599942.
- ^ Думан Дж. Г., Олсен Т. М. (1993). «Термический гистерезис активности белков у бактерий, грибов и филогенетически разнообразных растений». Криобиология. 30 (3): 322–328. Дои:10.1006 / cryo.1993.1031.
- ^ Кларк CJ, Бакли SL, Линднер N (2002). «Белки, структурирующие лед - новое название антифризов». Крио письма. 23 (2): 89–92. PMID 12050776.
- ^ Мерретт Н. (31 июля 2007 г.). «Белок Unilever продвигает Великобританию». DairyReporter.
- ^ «Может ли мороженое быть вкусным и полезным?». Журнал Impact. Ноттингемский университет. 18 сентября 2014 г.
- ^ Дортч Э. (2006). «Рыбные ГМ дрожжи, используемые для приготовления мороженого». Сеть заинтересованных фермеров. Архивировано из оригинал 14 июля 2011 г.. Получено 9 октября 2006.
- ^ Москин Дж. (26 июля 2006 г.). «Сливочное, более полезное мороженое? В чем выгода?». Нью-Йорк Таймс.
- ^ Реганд А., Гофф HD (январь 2006 г.). «Ингибирование перекристаллизации льда в мороженом под воздействием структурирующих лед протеинов из озимой пшеницы». Журнал молочной науки. 89 (1): 49–57. Дои:10.3168 / jds.S0022-0302 (06) 72068-9. PMID 16357267.
- ^ Челик Ю., Грэм Л.А., Мок Ю.Ф., Бар М., Дэвис П.Л., Браславский И. (2010). «Перегрев льда в присутствии связывающих его белков». Биофизический журнал. 98 (3): 245a. Bibcode:2010BpJ .... 98..245C. Дои:10.1016 / j.bpj.2009.12.1331. Сложить резюме – Physorg.com.
дальнейшее чтение
- Хаймет А.Д., Уорд Л.Г., Хардинг М.М. (1999). «Белки зимней камбалы« антифриз »: синтез и ингибирование роста льда аналогов, которые исследуют относительную важность гидрофобных и водородных взаимодействий». Журнал Американского химического общества. 121 (5): 941–948. Дои:10.1021 / ja9801341. ISSN 0002-7863.
- Сичери Ф., Ян Д.С. (июнь 1995 г.). «Связующая со льдом структура и механизм антифриза из камбалы озимой». Природа. 375 (6530): 427–31. Bibcode:1995Натура 375..427S. Дои:10.1038 / 375427a0. HDL:11375/7005. PMID 7760940. S2CID 758990.
внешняя ссылка
- Холодный, твердый факт: рыбный антифриз, производимый поджелудочной железой
- Белки-антифризы: молекула месяца, Дэвид Гудселл, RCSB Protein Data Bank
- Обзор всей структурной информации, доступной в PDB за UniProt: Q9GTP0 (Тепловой гистерезис или антифризный протеин) на PDBe-KB.