Дождевой червь - Earthworm - Wikipedia
| Дождевой червь | |
|---|---|
 | |
| Дождевой червь с хорошо развитым клитор | |
| Научная классификация | |
| Королевство: | |
| Тип: | |
| Учебный класс: | |
| Подкласс: | |
| Заказ: | |
An дождевой червь наземное беспозвоночное, принадлежащее к отряду Опистопора. Они демонстрируют трубку в трубке план тела, сегментированы внешне с соответствующей внутренней сегментацией и обычно имеют щетинки по всем сегментам.[1] Они встречаются по всему миру, где позволяют почва, вода и температура.[2] Дождевые черви обычно встречаются в почва, поедая самые разные органические вещества.[3] К этому органическому веществу относятся растительные вещества, живые простейшие, коловратки, нематоды, бактерии, грибы, и другие микроорганизмы.[4] Пищеварительная система дождевого червя проходит по всей длине его тела.[5] Это дышит через кожу. Имеет двойную транспортную систему из целомическая жидкость что движется внутри наполненного жидкостью целомудрие и простая закрытая система кровообращения. Оно имеет центральный и периферическая нервная система. Его центральная нервная система состоит из двух ганглии над ртом, по одному с каждой стороны, соединен с нервом, идущим вдоль его длины к двигательные нейроны и сенсорные клетки в каждом сегменте. Большое количество хеморецепторы сконцентрируйтесь возле рта. Окружные и продольные мышцы, окаймляющие каждый сегмент, позволяют червю двигаться. Подобные наборы мышц выстилают кишечник, и их действия перемещают перевариваемую пищу к анусу червя.[6]
Дождевые черви гермафродиты: каждый несет мужские и женские половые органы. Как беспозвоночные, им не хватает истинного скелет, но сохраняют свою структуру с помощью заполненных жидкостью целом камер, которые функционируют как гидростатический каркас.
«Дождевой червь» - это общее название для самых крупных представителей Oligochaeta (который является классом или подклассом в зависимости от автора). В классических системах они были в порядке Опистопора, поскольку мужские поры открываются кзади от женских пор, хотя внутренние мужские сегменты находятся впереди женских. Теоретическая кладистический исследования поместили их в подотряд Lumbricina порядка Гаплотаксида, но это может скоро измениться. Народные названия дождевого червя включают «червь росы», «дождевой червь», «ночной ползун» и «угловой червь» (от его использования в качестве рыболовная приманка ).
Более крупных наземных дождевых червей также называют мегадрилы (переводится как «большие черви»), в отличие от микродрили («маленькие черви») в полуводных семьях Tubificidae, Люмбрициды, и Enchytraeidae. Мегадрилы характеризуются отчетливым клитор (более обширная, чем у микродрилей) и сосудистая система с настоящими капиллярами.[7]
Анатомия
Форма и функция
В зависимости от вида взрослый дождевой червь может иметь длину от 10 мм (0,39 дюйма) и ширину от 1 мм (0,039 дюйма) до 3 м (9,8 фута) в длину и более 25 мм (0,98 дюйма) в ширину, но обычно Lumbricus terrestris вырастает примерно до 360 мм (14 дюймов) в длину.[8] Вероятно, самый длинный червь из подтвержденных записей - это Аминтас меконгиан расширяющийся до 3 м (10 футов) [9] в грязи по берегам 4350 км (2703 миль) Река Меконг в Юго-Восточной Азии.
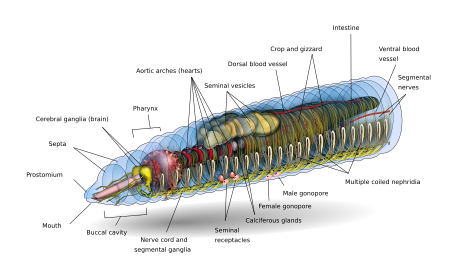
Основная форма дождевого червя спереди назад представляет собой цилиндрическую трубку в трубке, разделенную на ряд сегментов (называемых метамеризмы ), которые разделяют тело. Борозды обычно[10] внешне видны на теле, разграничивающие сегменты; дорсальные поры и нефридиопоры выделять жидкость, которая увлажняет и защищает поверхность червя, позволяя ему дышать. За исключением рта и анального сегмента, каждый сегмент несет щетиноподобные волоски, называемые боковыми. щетинки[11] используется для закрепления частей тела во время движения;[12] виды могут иметь четыре пары щетинок на каждом сегменте или более восьми, иногда образующие полный круг щетинок на сегмент.[11] Специальные вентральные щетинки используются для закрепления червей при спаривании путем их проникновения в тела своих товарищей.[13]
Как правило, в пределах одного вида количество найденных сегментов одинаково для разных экземпляров, и люди рождаются с таким количеством сегментов, которое они будут иметь на протяжении всей своей жизни. Первый сегмент тела (сегмент номер 1) включает как рот дождевого червя, так и выступающую над ним мясистую долю, называемую простомиум, который закрывает вход, когда червь находится в состоянии покоя, но также используется, чтобы чувствовать и химически определять окружение червя. Некоторые виды дождевых червей могут даже использовать цепкий простомиум, чтобы хватать и перетаскивать такие предметы, как трава и листья, в свою нору.
У взрослого дождевого червя развивается поясная железистая опухоль, называемая клитор, который охватывает несколько сегментов по направлению к передней части животного. Это часть репродуктивной системы, которая производит капсулы яйца. В задний чаще всего имеет цилиндрическую форму, как и остальная часть тела, но в зависимости от вида может быть четырехугольной, восьмиугольной, трапециевидной или уплощенной. Последний сегмент называется перипрокт; анус дождевого червя, короткая вертикальная щель, находится на этом сегменте.[11]

Внешний вид отдельного сегмента - тонкий кутикула на коже, обычно пигментированный от красного до коричневого, который имеет специализированные клетки, которые выделяют слизь над кутикулой, чтобы поддерживать тело влажным и облегчать движение через почву. Под кожей находится слой нервной ткани и два слоя мышц: тонкий внешний слой круговой мышцы и гораздо более толстый внутренний слой продольной мышцы.[14] Внутри мышечного слоя находится заполненная жидкостью камера, называемая целомудрие[15] что своим давлением обеспечивает структуру бескостного тела червя. Сегменты отделены друг от друга перегородками (множественное число от «septum»).[16] которые представляют собой перфорированные поперечные стенки, позволяющие целомической жидкости проходить между сегментами.[17] Пара структур называется нефростомы расположены в задней части каждой перегородки; почечный канальец проходит от каждого нефростома через перегородку в следующий сегмент. Затем этот каналец ведет к главному фильтрующему органу тела, нефридиум или метанефридий, который удаляет метаболические отходы из целомическая жидкость и выводит его через поры, называемые нефридиопорами, на боках червя; обычно в большинстве сегментов обнаруживаются две нефридии (иногда и больше).[18] В центре червя находится пищеварительный тракт, который проходит прямо ото рта к анальному отверстию, не скручиваясь, и фланкируется сверху и снизу кровеносными сосудами (дорсальный кровеносный сосуд и вентральный кровеносный сосуд, а также субневральный кровеносный сосуд) и брюшной нервный тяж, и окружен в каждом сегменте парой паллиальных кровеносных сосудов, которые соединяют дорсальные и субневральные кровеносные сосуды.
Многие дождевые черви могут выбрасывать целомическую жидкость через поры в спине в ответ на стресс; австралийский Didymogaster sylvaticus (известный как "синий дождевой червь-брызгун") может разбрызгивать жидкость на высоту до 30 см (12 дюймов).[19][17]
Нервная система

Центральная нервная система
ЦНС состоит из двулопастного мозг (мозговой ганглии, или надглоточный ганглии), подглоточные ганглии, окологлоточные соединительные элементы и брюшной нервный тяж.
Мозг дождевых червей состоит из пары церебральных ганглиев грушевидной формы. Они расположены на дорсальной стороне пищеварительного канала в третьем сегменте, в канавке между Полость рта и глотка.
Пара окологлоточных соединительных элементов головного мозга окружает глотку, а затем соединяется с парой подглоточных ганглиев, расположенных ниже глотки в четвертом сегменте. Такое расположение означает, что мозг, подглоточные ганглии и окологлоточные соединительные элементы образуют нервное кольцо вокруг глотки.
Брюшной нервный тяж (образованный нервными клетками и нервными волокнами) начинается от подглоточных ганглиев и простирается ниже пищеварительного канала до самого заднего сегмента тела. Брюшной нервный канатик имеет вздутие или ганглии в каждом сегменте, то есть сегментарный ганглий, который возникает с пятого до последнего сегмента тела. Также есть три гигантских аксоны, один медиальный гигантский аксон (MGA) и два боковых гигантских аксона (LGAs) на средне-дорсальной стороне вентрального нервного тяжа. MGA имеет диаметр 0,07 мм и передает в переднезаднем направлении со скоростью 32,2 м / с. LGA немного уже, 0,05 мм в диаметре и пропускают в задне-переднем направлении со скоростью 12,6 м / с. Два LGA соединены через равные промежутки между телом и поэтому считаются одним гигантским аксоном.[20][21]
Периферическая нервная система
- От церебральных ганглиев отходят от восьми до десяти нервов, которые снабжают простомиум, щечная камера и глотка.
- Три пары нервов отходят от подфарианговых ганглиев, чтобы снабжать 2, 3 и 4 сегмент.
- От каждой отходят по три пары нервов. сегментарный ганглий поставлять различные конструкции сегмента.
Симпатическая нервная система состоит из нервных сплетений в эпидермисе и пищеварительном тракте. (Сплетение - это сеть соединенных нервных клеток.) Нервы, которые проходят вдоль стенки тела, проходят между внешним круговым и внутренним продольными мышечными слоями стенки. От них отходят ветви, образующие межмышечное сплетение и субэпидермальное сплетение. Эти нервы соединяются с перепончатоглоточной соединительной частью.
Движение
На поверхности скорость ползания варьируется как внутри, так и среди людей. Дождевые черви ползают быстрее, прежде всего, делая более длинные «шаги» и более частые шаги. Больше Lumbricus terrestris черви ползают с большей абсолютной скоростью, чем более мелкие черви. Они достигают этого, делая несколько более длинные шаги, но с немного более низкой частотой шагов.[22]
Прикосновение к дождевому червю, которое вызывает реакцию "давления", а также (часто) реакцию на обезвоживающую способность соли на коже человека (токсичную для дождевых червей), стимулирует субэпидермальное нервное сплетение, которое соединяется с межмышечным сплетением и вызывает продольное мышцы сокращаются. Это вызывает извивающиеся движения, наблюдаемые, когда человек берет в руки дождевого червя. Такое поведение рефлекс и не требует ЦНС; это происходит даже при удалении нервного шнура. Каждый сегмент дождевого червя имеет собственное нервное сплетение. Сплетение одного сегмента не связано напрямую со сплетением соседних сегментов. Нервный шнур необходим для соединения нервной системы сегментов.[23]
Гигантские аксоны несут самые быстрые сигналы по нервному шнуру. Это аварийные сигналы, которые вызывают рефлекторное побегание. Более крупный дорсальный гигантский аксон передает сигналы быстрее всего от задней части к передней части животного. Если прикоснуться к задней части червя, быстро передается сигнал, заставляющий сокращаться продольные мышцы каждого сегмента. Это приводит к тому, что червь очень быстро укорачивается при попытке убежать от хищника или другой потенциальной угрозы. Два медиальных гигантских аксона соединяются друг с другом и посылают сигналы спереди назад. Их раздражение заставляет дождевого червя очень быстро отступать (возможно, он сжимается в своей норе, чтобы спастись от птицы).
Наличие нервной системы важно, чтобы животное могло испытать ноцицепция или же боль. Однако также требуются другие физиологические возможности, такие как чувствительность к опиоидам и центральная модуляция ответов анальгетиками.[24] Энкефалин и α-эндорфин -подобные вещества обнаружены у дождевых червей. Инъекции налоксон (антагонист опиоидов) подавляют реакцию бегства дождевых червей. Это указывает на то, что опиоидные вещества играют роль в сенсорной модуляции, аналогичную той, которая обнаруживается у многих позвоночных.[25]
Сенсорный прием
Светочувствительность
У дождевых червей нет глаза (хотя у некоторых червей есть), однако у них есть специализированные светочувствительные клетки, называемые «световыми клетками Гесса». Эти фоторецепторные клетки имеют центральную внутриклеточную полость (фагосома ) наполненный микроворсинки. Помимо микроворсинок, в фагосоме есть несколько сенсорных ресничек, структурно независимых от микроворсинок.[26] Фоторецепторы распределены в большинстве частей эпидермиса, но больше сосредоточены на спине и боках червя. Относительно небольшое количество встречается на вентральной поверхности 1-го сегмента. Они наиболее многочисленны в простомиуме и уменьшаются в плотности в первых трех сегментах; после третьего сегмента их очень мало.[23]
Эпидермальный рецептор (орган чувств)
Эти рецепторы многочисленны и распространены по всей эпидермис. Каждый рецептор имеет слегка приподнятую кутикулу, которая покрывает группу высоких, тонких и столбчатых рецепторных клеток. Эти клетки несут на своих внешних концах небольшие волосовидные отростки, а их внутренние концы связаны с нервными волокнами. Эпидермальные рецепторы обладают осязательной функцией. Они также озабочены изменениями температуры и реагируют на химические раздражители. Дождевые черви чрезвычайно чувствительны к прикосновению и механической вибрации.
Буккальный рецептор (орган чувств)
Эти рецепторы расположены только в эпителии щечной камеры. Эти рецепторы являются вкусовыми и обонятельными (связаны со вкусом и запахом). Они также реагируют на химические раздражители. (Хеморецептор)
Пищеварительная система
В кишка дождевого червя представляет собой прямую трубку, идущую от пасти червя до его анус. Он подразделяется на пищеварительный тракт и связанные железы, которые встроены в стенку самого пищеварительного канала. Пищеварительный канал состоит из рта, ротовой полости (обычно проходит через первые один или два сегмента дождевого червя), глотки (обычно около четырех сегментов в длину), пищевода, зоба, желудка (обычно) и кишечника.[27]
Пища попадает в рот. В глотка действует как всасывающий насос; его мускулистые стенки втягивают пищу. В глотке глоточные железы выделяют слизь. Еда перемещается в пищевод, куда кальций (из крови и проглоченный от предыдущего приема пищи) закачивается для поддержания надлежащего уровня кальция в крови и пище pH. Оттуда пища попадает в зоб и желудок. в желудок, сильные мышечные сокращения измельчают пищу с помощью минеральных частиц, попадающих вместе с пищей. Пройдя через желудок, пища попадает в кишечник для переваривания. Кишечник выделяет пепсин переваривать белки, амилаза переваривать полисахариды, целлюлаза для переваривания целлюлозы и липазы для переваривания жиров.[6] В дополнение к пищеварительным белкам дождевые черви используют класс поверхностно-активных соединений, называемых дрилодефензины, которые помогают переваривать растительный материал.[28] Вместо того, чтобы свернуться, как кишечник млекопитающих, в кишечнике дождевого червя присутствует большая срединно-спинная язычковая складка, называемая тифлозоль который увеличивает площадь поверхности для увеличения усвоения питательных веществ за счет множества складок, расположенных вдоль его длины. Кишечник имеет свою собственную пару мышечных слоев, как и тело, но в обратном порядке - внутренний круговой слой внутри внешнего продольного слоя.[29]
Сердечно-сосудистая система
Дождевые черви имеют двойную систему кровообращения, в которой как целомическая жидкость, так и замкнутая система кровообращения переносят пищу, отходы и дыхательные газы. Замкнутая система кровообращения имеет пять основных кровеносных сосудов: спинной (верхний) сосуд, который проходит над пищеварительным трактом; вентральный (нижний) сосуд, идущий ниже пищеварительного тракта; субневральный сосуд, который проходит ниже брюшного нервного канатика; и два латероневральных сосуда по обе стороны нервного канатика.[30]
Спинной сосуд - это в основном собирающая структура в области кишечника. Он получает парную комиссуральную и спинную кишки в каждом сегменте. Вентральный сосуд разветвляется на пару вентро-тегументариев и вентро-кишечных костей в каждом сегменте. Подневральный сосуд также выделяет пару спаек, идущих по задней поверхности перегородки.
Насосное действие на спинной сосуд перемещает кровь вперед, в то время как другие четыре продольных сосуда несут кровь назад. В сегментах с седьмого по одиннадцатый пара дуг аорты окружает целому и действует как сердце, перекачивая кровь к вентральному сосуду, который действует как аорта. Кровь состоит из амебоидных клеток и растворенного в плазме гемоглобина. Вторая кровеносная система происходит из клеток пищеварительной системы, выстилающих целом. Когда пищеварительные клетки наполняются, они высвобождают неживые клетки жира в заполненный жидкостью целом, где они свободно плавают, но могут проходить через стенки, разделяющие каждый сегмент, перемещая пищу в другие части и способствуя заживлению ран.[31]
Выделительная система
Экскреторная система содержит пару нефридия в каждом сегменте, кроме первых трех и последнего.[32] Три типа нефридий: покровные, перегородочные и глоточные. Покровные нефридии прикрепляются к внутренней стороне стенки тела во всех сегментах, кроме первых двух. Септальные нефридии прикрепляются к обеим сторонам перегородки позади 15-го сегмента. Нефридии глотки прикрепляются к четвертому, пятому и шестому сегментам.[32] Отходы в целом жидкости из переднего сегмента втягиваются за счет биения реснички из нефростома. Оттуда он проходит через перегородку (стенку) через трубку, которая образует серию петель, переплетенных кровеносными капиллярами, которые также переносят отходы в канальцы нефростома. Затем экскреторные отходы выводятся через поры на стороне червя.[33]
Дыхание
Особых органов дыхания у дождевых червей нет. Обмен газов происходит через влажную кожу и капилляры, где кислород улавливается гемоглобином, растворенным в плазме крови, и выделяется диоксид углерода. Вода, как и соли, также может перемещаться через кожу посредством активного транспорта.
Жизнь и физиология
При рождении дождевые черви появляются небольшими, но полностью сформированными, лишенными только половых структур, которые развиваются примерно за 60-90 дней. Они достигают полного размера примерно за год. Ученые прогнозируют, что средняя продолжительность жизни в полевых условиях составляет от четырех до восьми лет, в то время как большинство садовых сортов живут только один-два года.
Размножение



Несколько распространенных видов дождевых червей в основном партеногенетический, что означает рост и развитие эмбрионы происходит без оплодотворение.Среди люмбрицид у дождевых червей партеногенез неоднократно возникал от половых родственников.[34] Партеногенез в некоторых Aporrectodea trapezoides линии произошли от 6,4 до 1,1 миллиона лет назад от половых предков.[35] Некоторые виды демонстрируют псевдогамный партогенез, то есть спаривание необходимо для стимуляции воспроизводства, даже если мужской генетический материал не передается потомству.[36]
Спаривание дождевых червей происходит на поверхности, чаще всего ночью. Дождевые черви гермафродиты; то есть у них есть как мужские, так и женские половые органы. Половые органы расположены в сегментах с 9 по 15. У дождевых червей одна или две пары семенников содержатся в мешочках. Две или четыре пары семенные пузырьки производить, хранить и выпускать сперму через мужские поры. Яичники и яйцеводы в сегменте 13 выпускают яйца через поры самок в сегменте 14, в то время как сперма изгоняется из сегмента 15. Одна или несколько пар сперматека присутствуют в сегментах 9 и 10 (в зависимости от вида), которые представляют собой внутренние мешочки, которые принимают и хранят сперму от другого червя во время совокупления. В результате сегмент 15 одного червя выделяет сперму в сегменты 9 и 10 вместе с пузырьками хранения своего партнера. Некоторые виды используют внешние сперматофоры для передачи спермы.
В Hormogaster samnitica и Hormogaster elisae транскриптом Библиотеки ДНК были секвенированы и два пола феромоны, Аттрактин и Темптин были обнаружены во всех образцах тканей обоих разновидность.[37] Половые феромоны, вероятно, важны для дождевых червей, потому что они живут в среде, где химическая сигнализация может играть решающую роль в привлечении партнера и облегчении ауткроссинга. Ауткроссинг обеспечит преимущество маскировки экспрессии вредных рецессивных мутаций в потомстве.[38] (видеть Дополнение ).
Совокупление и воспроизведение отдельные процессы у дождевых червей. Брачная пара перекрывает передние концы вентрально, и каждый обменивается спермой друг с другом. В клитор становится от очень красноватого до розоватого цвета. Спустя какое-то время после совокупления, спустя много времени после разделения червей, клитор (позади сперматеек) выделяет материал, который образует кольцо вокруг червя. Затем червь выходит из кольца и при этом вводит в него свои собственные яйца и сперму другого червя. Таким образом, каждый червь становится генетическим отцом одного из своих потомков и генетической матерью остальных. Когда червяк выскользнул из кольца, концы кокона запечатывались, образуя инкубатор, слегка напоминающий луковицу (кокон ), в которых развиваются эмбриональные черви. Следовательно, оплодотворение внешнее. Затем кокон откладывается в почве. Через три недели вылупляется от 2 до 20 молодых особей, в среднем 4. Развитие идет напрямую, то есть без образования личинок.
Передвижение

Дождевые черви перемещаются под землей с помощью волн мышечных сокращений, которые попеременно укорачивают и удлиняют тело (перистальтика ). Укороченная часть прикреплена к окружающей почве крошечными когтеобразными щетинками (щетинки ), установленный по его длине сегментов. На всех сегментах тела, кроме первого, последнего и клитора, имеется кольцо из S-образных щетинок, внедренное в эпидермальную ямку каждого сегмента (перихетин). Весь процесс закапывания поддерживается секрецией смазывающей слизи. В результате своего движения по смазанным туннелям черви могут издавать булькающие звуки под землей, если их потревожить. Дождевые черви перемещаются по почве, с силой расширяя щели; когда силы измеряются в соответствии с массой тела, детеныши могут толкаться в 500 раз больше собственного веса, тогда как крупные взрослые особи могут толкать только в 10 раз больше собственного веса.[39]
Регенерация
Дождевые черви обладают способностью восстанавливать потерянные сегменты, но эта способность варьируется у разных видов и зависит от степени повреждения. Стивенсон (1930) посвятил этой теме главу своей монографии, а Г. Гейтс потратил 20 лет на изучение регенерации у множества видов, но «из-за того, что проявился небольшой интерес», Гейтс (1972) опубликовал лишь несколько своих выводов, которые, тем не менее, показывают, что теоретически возможно вырастить двух целых червей из разделенного пополам экземпляра. у определенных видов.
Отчеты Гейтса включали:
- Eisenia fetida (Савиньи, 1826) с регенерацией головы в переднем направлении, возможной на каждом межсегментарном уровне назад до 23/24 включительно, тогда как хвосты регенерировались на любых уровнях за 20/21, т.е. два червя могут вырасти из одного.[40]
- Lumbricus terrestris (Линней, 1758 ) замена передних сегментов от 13/14 и 16/17, но регенерация хвоста так и не была обнаружена.
- Экскаваторский перионикс (Perrier, 1872 г.) легко регенерированные утраченные части тела, в переднем направлении уже от 17/18 и в заднем направлении до 20/21.
- Lampito mauritii (Кинберг, 1867 г.) с регенерацией в переднем направлении на всех уровнях назад до 25/26 и регенерацией хвоста от 30/31; Иногда считалось, что регенерация головы вызвана внутренней ампутацией в результате Саркофага sp. личиночная инвазия.
- Criodrilus lacuum (Хоффмайстер, 1845 г.) также обладает потрясающей регенеративной способностью с регенерацией «головы» еще с 40/41.[41]
Сообщается о неопознанном тасманском дождевом черве, у которого растет новая голова.[42]
Таксономия и распространение
В мире таксономии устойчивая «классическая система» Михаэльсена (1900) и Стивенсона (1930) постепенно размывалась спорами о том, как классифицировать дождевых червей, так что Фендер и Макки-Фендер (1990) зашли так далеко, что сказали: , "На уровне семьи классификация мегасколецид дождевые черви в хаосе ".[43] За прошедшие годы многие ученые разработали свои собственные системы классификации дождевых червей, что привело к путанице, и эти системы были и продолжают пересматриваться и обновляться. Используемая здесь система классификации, разработанная Блейкмором (2000), представляет собой современный возврат к классической системе, которая исторически доказана и широко принята.[44]
Категоризация мегадрил дождевого червя в одно из его таксономических семейств под отрядами Lumbricina и Монилигастрида основывается на таких характеристиках, как состав клитора, расположение и расположение половых признаков (поры, предстательные железы и т. д.), количество желудков и форма тела.[44] В настоящее время названо более 6000 видов наземных дождевых червей, как это предусмотрено в базе данных названий видов,[45] но количество синонимов неизвестно.
Семьи с их известным распределением или происхождением:[44]
- Acanthodrilidae - (Гондванцы или пангеи?)
- Ailoscolecidae - Пиренеи и юго-восток США
- Almidae - тропический экваториальный (Южная Америка, Африка, Индо-Азия)
- Benhamiinae - эфиопский, неотропический (возможное подсемейство Octochaetidae)
- Criodrilidae - юго-западная Палеарктика: Европа, Ближний Восток, Россия и Сибирь до побережья Тихого океана; Япония (Бивадрилус); в основном водные
- Diplocardiinae / -idae - гондванские или лауразийские? (подсемейство Acanthodrilidae)
- Enchytraeidae - космополитичный, но необычный в тропиках (обычно относится к микродрилам)
- Eudrilidae - Тропическая Африка к югу от Сахары
- Exxidae - Neotropical: Центральная Америка и Карибский бассейн
- Glossoscolecidae - Неотропический: Центральная и Южная Америка, Карибский бассейн
- Haplotaxidae - космополитическое распространение (обычно относится к микродрилам)
- Hormogastridae - Средиземноморье
- Kynotidae - Малагасийцы: Мадагаскар
- Люмбрициды - Голарктика: Северная Америка, Европа, Ближний Восток, Центральная Азия и Япония.
- Lutodrilidae - Луизиана, юго-восток США
- Megascolecidae - (Пангеи?)
- Microchaetidae - наземные животные в Африке, особенно на лугах Южной Африки.
- Moniligastridae - Восточно-Индийский субрегион
- Ocnerodrilidae - Неотропы, Африка; Индия
- Octochaetidae - австралийский, индийский, восточный, эфиопский, неотропический
- Octochaetinae - австралийские, индийские, восточные (подсемейство, если принято Benhamiinae)
- Sparganophilidae - Неарктика, Неотропия: Северная и Центральная Америка
- Tumakidae - Колумбия, Южная Америка
Как инвазивный вид
Из примерно 7000 видов только около 150 широко распространены по всему миру. Это сапсаны или космополитические дождевые черви.[46]
Экология

Дождевые черви подразделяются на три основные экофизиологические категории: (1) черви, обитающие в листовой подстилке или компосте, которые не зарываются, живут на границе раздела почва и подстилка и поедают разлагающиеся органические вещества (эпигейский ) например Eisenia fetida; (2) черви, обитающие в верхнем или нижнем слое почвы, которые питаются (на почве), зарываются и закапываются в почву, создавая горизонтальные норы в верхних 10–30 см почвы (эндогенный ); и (3) черви, которые строят постоянные глубокие вертикальные норы, которые они используют, чтобы посещать поверхность для получения растительного материала для пищи, такого как листья (анекдот, что означает "тянуться вверх"), например Lumbricus terrestris.[47]
Популяции дождевых червей зависят как от физических, так и от химических свойств почвы, таких как температура, влажность, pH, соли, аэрация и текстура, а также доступная пища и способность видов к размножению и распространению. Одним из важнейших факторов окружающей среды является pH, но у дождевых червей разные предпочтения. Большинство предпочитает нейтральные или слабокислые почвы. Люмбрикус Террестрис все еще присутствует при pH 5,4, Dendrobaena octaedra при pH 4,3 и некоторых Megascolecidae присутствуют в чрезвычайно кислых гуминовых почвах. PH почвы также может влиять на количество червей, которые диапауза. Чем более кислая почва, тем раньше черви впадают в диапаузу и дольше остаются в диапаузе при pH 6,4.
На дождевых червей охотятся многие виды птицы (например, малиновки, скворцы, дрозды, чайки, вороны ), змеи, деревянные черепахи, млекопитающие (например, медведи, кабаны, лисы, ежики, свиньи, родинки [48]) и беспозвоночных (например, муравьи,[49] плоские черви, жужелицы и другие жуки, улитки, пауки, и слизни ). У дождевых червей много внутренних паразиты, включая простейшие, платигельминты и нематоды; их можно найти в червях кровь, семенные пузырьки, целомудрие, или же кишечник, или в их коконы.
Азотные удобрения имеют тенденцию создавать кислые условия, которые смертельны для червей, и мертвые образцы часто обнаруживаются на поверхности после нанесения таких веществ, как ДДТ, известь сера, и арсенат свинца. В Австралия, изменения в методах ведения сельского хозяйства, такие как применение суперфосфаты на пастбища и переход от пастырское земледелие к земледелие оказал разрушительное воздействие на население гигантского Дождевой червь Gippsland, что привело к их классификации как охраняемые виды. В глобальном масштабе некоторые популяции дождевых червей были опустошены из-за отклонения от органического производства и распыления синтетических удобрений и биоцидов, при этом по крайней мере три вида теперь внесены в список вымерших, но многие другие находятся под угрозой исчезновения.[50]
Вермикомпостирование всех органических "отходов" и добавление этого органического вещества, предпочтительно в виде поверхности мульча, на регулярной основе обеспечит дождевых червей их потребностями в пище и питательных веществах, а также создаст оптимальные условия температуры и влажности, которые естественным образом будут стимулировать их активность.
Эта деятельность дождевых червей аэрирует и перемешивает почву, способствует минерализации питательных веществ и их поглощению растительностью. Определенные виды дождевых червей выходят на поверхность и питаются более высокими концентрациями органических веществ, присутствующих там, смешивая их с минеральной почвой. Поскольку высокий уровень смешивания органических веществ связан с плодородие почвы, фермеры и садоводы считают обилие дождевых червей полезным.[51][52] Еще в 1881 году Чарльз Дарвин писал: «Можно сомневаться, есть ли много других животных, сыгравших такую важную роль в истории мира, как эти низкоорганизованные создания».[53]

Кроме того, хотя, как следует из названия, основная среда обитания дождевых червей находится в почве, они не ограничиваются этой средой обитания. Червь-бренди Eisenia fetida живет в разлагающихся растениях и навозе. Arctiostrotus vancouverensis из Остров Ванкувер и Олимпийский полуостров обычно встречается в гниющих бревнах хвойных пород. Aporrectodea limicola, Спарганофил spp., а некоторые другие встречаются в илах ручьев. Некоторые виды древесные,[нужна цитата ] некоторые водные и некоторые эвригалин (устойчивы к соленой воде) и прибрежные (живущие на берегу моря, например Pontodrilus litoralis ).[54] Даже среди почвенных видов особые среды обитания, такие как почвы, полученные из змеевик, имеют собственную фауну дождевых червей.
Воздействие на окружающую среду
Основные преимущества деятельности дождевых червей для плодородия почвы для сельского хозяйства можно резюмировать следующим образом:
- Биологические: Во многих почвах дождевые черви играют важную роль в превращении крупных частиц органического вещества в богатые перегной, тем самым улучшая плодородие почвы. Это достигается за счет того, что червь втягивает под поверхность осевшие органические вещества, такие как опадание листьев или навоз, либо в пищу, либо для закупоривания своей норы. Попав в нору, червь измельчит лист, частично его переварит и смешает с землей. Отливки червя (см. Нижний правый) могут содержать на 40 процентов больше гумуса, чем верхние 9 дюймов (23 см) почвы, в которой живет червь.[55]

- Химическая: Помимо мертвых органическая материя, дождевой червь также заглатывает любые другие достаточно мелкие частицы почвы, в том числе песчинки размером до 1/20 дюйма (1,25 мм), в свой желудок, где эти мельчайшие частицы измельчают все в мелкую пасту, которая затем переваривается в кишечнике. Когда червь выделяет это в виде слепков, откладывающихся на поверхности или глубже в почве, минералы и питательные вещества для растений превращаются в доступную для растений форму. Исследования, проведенные в США, показывают, что свежие слепки дождевых червей в пять раз богаче доступными. азот, в семь раз богаче доступными фосфаты, и в 11 раз богаче доступными калий чем окружающие верхние 6 дюймов (150 мм) почвы. В условиях обильного гумуса вес получаемых слепков может превышать 4,5 кг (10 фунтов) на червя в год.[55]
- Физический: Копание дождевого червя создает множество каналов в почве и имеет большое значение для поддержания структура почвы, обеспечивающие процессы аэрации и дренажа. Пермакультура соучредитель Билл Моллисон указывает, что, скользя по своим туннелям, дождевые черви «действуют как бесчисленная армия поршней, нагнетающих воздух в почву и из почвы в течение 24-часового цикла (быстрее ночью)».[56] Таким образом, дождевой червь не только создает проходы для воздуха и воды, проходящие через почву, но также изменяет жизненно важный органический компонент, который делает почву здоровой (см. Биотурбация ). Дождевые черви способствуют формированию богатых питательными веществами слепков (шариков почвы, устойчивых в почвенной слизи), которые обладают высокой агрегацией почвы, а также ее плодородием и качеством.[55] В подзол почвы, дождевые черви могут стереть характерный полосатый вид почвенного профиля путем смешивания органического (LFH), элювиального (E) и верхнего иллювиального (B) горизонтов, чтобы создать единый темный горизонт Ap.[57][58]
Дождевые черви ускоряют круговорот питательных веществ в системе почва-растение за счет фрагментации и перемешивания растительных остатков - физического измельчения и химического переваривания.[55] Существование дождевого червя нельзя считать само собой разумеющимся. Доктор У. Э. Шевелл-Купер наблюдаются «огромные количественные различия между соседними садами», а на популяции червей влияет множество факторов окружающей среды, на многие из которых могут повлиять надлежащие методы управления со стороны садовода или фермера.[59]
Дарвин оценил, что пахотная земля содержит до 53000 червей на акр (13 / м2), но более недавние исследования дали цифры, предполагающие, что даже плохая почва может поддерживать 250 000 / акр (62 / м2), в то время как на богатых плодородных сельскохозяйственных угодьях может быть до 1,750,000 / акр (432 / м2), meaning that the weight of earthworms beneath a farmer's soil could be greater than that of the livestock upon its surface. Richly organic topsoil populations of earthworms are much higher – averaging 500 worms m−2 and up to 400 gm−2 – such that, for the 7 billion of us, each person alive today has support of 7 million earthworms.[60]
The ability to break down organic materials and excrete concentrated nutrients makes the earthworm a functional contributor in restoration projects. In response to ecosystem disturbances, some sites have utilized earthworms to prepare soil for the return of native flora. Исследования из Station d'écologie Tropicale de Lamto asserts that the earthworms positively influence the rate of macroaggregate formation, an important feature for soil structure.[61] The stability of aggregates in response to water was also found to be improved when constructed by earthworms.[61]
Though not fully quantified yet, Выбросы парниковых газов of earthworms likely contribute to global warming, especially since top-dwelling earthworms increase the speed of carbon cycles and have been spread by humans into many new geographies.[62]
Экономическое влияние

Various species of worms are used in вермикультура, the practice of feeding organic waste to earthworms to decompose food waste. These are usually Eisenia fetida (or its close relative Eisenia andrei ) or the Brandling worm, commonly known as the tiger worm or red wiggler. They are distinct from soil-dwelling earthworms. In the tropics, the African nightcrawler Eudrilus eugeniae [63] and the Indian blue Экскаваторский перионикс используются.
Earthworms are sold all over the world; the market is sizable. According to Doug Collicut, "In 1980, 370 million worms were exported from Canada, with a Canadian export value of $13 million and an American retail value of $54 million."
Earthworms provide an excellent source of protein for fish, fowl and pigs but were also used traditionally for human consumption. Noke is a culinary term used by the Маори из Новая Зеландия, and refers to earthworms which are considered delicacies for their chiefs.
Смотрите также
- Drilosphere, the part of the soil influenced by earthworm secretions and castings
- Образование плесени на овощах под действием червей, an 1881 book by Charles Darwin
- Почвенная жизнь
- Биогумус
- Червь очаровательный
Рекомендации
- ^ Edwards, Clive A.; Lofty, J. R. (1977). Biology of Earthworms. Лондон: Чепмен и Холл. стр. предисловие. ISBN 0-412-14950-8.
- ^ Coleman, David C.; Crossley, D.A.; Hendrix, Paul F. (2004). Fundamentals of Soil Ecology. Амстердам; Бостон: Elsevier Academic Press. п. 170. ISBN 0-12-179726-0.
- ^ Bonkowski, Griffiths, and Ritz (November 2000). "Food preferences of earthworms for soil fungi". Педобиология. 44–6 (6): 667. Дои:10.1078/S0031-4056(04)70080-3.CS1 maint: несколько имен: список авторов (связь)
- ^ Lofty, Clive A.; Lofty, J. R. (1977). Biology of Earthworms. Лондон: Чепмен и Холл. п. 80. ISBN 0-412-14950-8.
- ^ Edwards, Clive A.; Lofty, J.R. (1977). Biology of Earthworms. Лондон: Чепмен и Холл. п. 19. ISBN 0-412-14950-8.
- ^ а б Cleveland P. Hickman Jr.; Larry S. Roberts; Frances M Hickman (1984). Integrated Principles of Zoology (7-е изд.). Times Mirror/Mosby College Publishing. п.344. ISBN 978-0-8016-2173-4.
- ^ Omodeo, Pietro (2000). "Evolution and biogeography of megadriles (Annelida, Clitellata)". Итальянский зоологический журнал. 67–2 (2): 179–201. Дои:10.1080/11250000009356313. S2CID 86293273.
- ^ Blakemore 2012, п. xl.
- ^ Blakemore, R. J.; и другие. (2007). "Megascolex (Promegascolex) mekongianus Cognetti, 1922 – its extent, ecology and allocation to Amynthas (Clitellata/Oligochaeta: Megascolecidae)" (PDF). Opuscula Zoologica.
- ^ Edwards & Bohlen 1996, п. 11.
- ^ а б c Sims & Gerard 1985, стр. 3–6.
- ^ Edwards & Bholen 1996, п. 3.
- ^ Feldkamp, J. (1924). "Feldkamp, J. "Untersuchungen über die Geschlechtsmerkmale und die Begattung der Regenwurmer Zoologische Jahrbücher". Anatomie. 46: 609–632.
- ^ Edwards & Bohlen 1996, п. 8-9.
- ^ Edwards & Bohlen 1996, п. 1.
- ^ Sims & Gerard 1985, п. 8.
- ^ а б Edwards & Bohlen 1996, п. 12.
- ^ Edwards & Bohlen 1996, п. 6.
- ^ Myrmecofourmis (2018). "Meet the squirting earthworm" (видео ). YouTube.
- ^ "Experiment: Comparing speeds of two nerve fiber sizes". BackyardBrains. Получено 4 апреля, 2015.
- ^ Drewes, C.D., Landa, K.B. and McFall, J.L. (1978). "Giant nerve fibre activity in intact, freely moving earthworms". Журнал экспериментальной биологии. 72: 217–227. PMID 624897.CS1 maint: несколько имен: список авторов (связь)
- ^ Quillin, K.J. (1999). "Kinematic scaling of locomotion by hydrostatic animals: ontogeny of peristaltic crawling by the earthworm lumbricus terrestris". Журнал экспериментальной биологии. 202: 661–674. PMID 10021320.
- ^ а б "Earthworm-nervous system". Cronodon. Получено 3 апреля, 2015.
- ^ Elwood, R.W. (2011). "Pain and suffering in invertebrates?". Журнал ILAR. 52 (2): 175–84. Дои:10.1093/ilar.52.2.175. PMID 21709310.
- ^ Смит, Дж. (1991). "A question of pain in invertebrates". Журнал ILAR. 33 (1–2): 25–31. Дои:10.1093/ilar.33.1-2.25.
- ^ Röhlich, P., Aros, B. and Virágh, Sz. (1970). "Fine structure of photoreceptor cells in the earthworm, Lumbricus Terrestris". Zeitschrift für Zellforschung und Mikroskopische Anatomie. 104 (3): 345–357. Дои:10.1007/BF00335687. PMID 4193489. S2CID 22771585.CS1 maint: несколько имен: список авторов (связь)
- ^ Edwards & Bohlen 1996, п. 13.
- ^ Liebeke, Manuel; Strittmatter, Nicole; Fearn, Sarah; Morgan, A. John; Kille, Peter; Fuchser, Jens; Wallis, David; Palchykov, Vitalii; Robertson, Jeremy (2015-08-04). "Unique metabolites protect earthworms against plant polyphenols". Nature Communications. 6: 7869. Bibcode:2015NatCo...6.7869L. Дои:10.1038/ncomms8869. ЧВК 4532835. PMID 26241769.
- ^ Edwards & Bohlen 1996, стр. 13–15.
- ^ Sims & Gerard 1985, п. 10.
- ^ Cleveland P. Hickman Jr.; Larry S. Roberts; Frances M Hickman (1984). Integrated Principles of Zoology (7-е изд.). Times Mirror/Mosby College Publishing. стр.344–345. ISBN 978-0-8016-2173-4.
- ^ а б Farabee, H.J. "Выделительная система". Архивировано из оригинал 30 июля 2012 г.. Получено 29 июля 2012.
- ^ Cleveland P. Hickman Jr.; Larry S. Roberts; Frances M Hickman (1984). Integrated Principles of Zoology (7-е изд.). Times Mirror/Mosby College Publishing. стр.345–346. ISBN 978-0-8016-2173-4.
- ^ Domínguez J, Aira M, Breinholt JW, Stojanovic M, James SW, Pérez-Losada M (2015). "Underground evolution: New roots for the old tree of lumbricid earthworms". Мол. Филогенет. Evol. 83: 7–19. Дои:10.1016/j.ympev.2014.10.024. ЧВК 4766815. PMID 25463017.
- ^ Fernández R, Almodóvar A, Novo M, Simancas B, Díaz Cosín DJ (2012). "Adding complexity to the complex: new insights into the phylogeny, diversification and origin of parthenogenesis in the Aporrectodea caliginosa species complex (Oligochaeta, Lumbricidae)". Мол. Филогенет. Evol. 64 (2): 368–79. Дои:10.1016/j.ympev.2012.04.011. PMID 22542691.
- ^ Cosín D.J.D., Novo M., Fernández R. (2011) Reproduction of Earthworms: Sexual Selection and Parthenogenesis. In: Karaca A. (eds) Biology of Earthworms. Soil Biology, vol 24. Springer, Berlin, Heidelberg, pp. 76ff. [1]
- ^ Novo M, Riesgo A, Fernández-Guerra A, Giribet G (2013). "Pheromone evolution, reproductive genes, and comparative transcriptomics in mediterranean earthworms (annelida, oligochaeta, hormogastridae)". Мол. Биол. Evol. 30 (7): 1614–29. Дои:10.1093/molbev/mst074. PMID 23596327.
- ^ Бернштейн Х., Хопф Ф.А., Мишод РЭ (1987). «Молекулярные основы эволюции пола». Молекулярная генетика развития. Adv. Genet. Успехи в генетике. 24. С. 323–70. Дои:10.1016 / S0065-2660 (08) 60012-7. ISBN 978-0-12-017624-3. PMID 3324702.
- ^ Quillan, K.J. (2000). "Ontogenetic scaling of burrowing forces in the earthworm Lumbricus terrestris". Журнал экспериментальной биологии. 203 (Pt 18): 2757–2770. PMID 10952876. Получено 4 апреля, 2015.
- ^ Gates, G. E. (April 1949). "Regeneration in an Earthworm, Eisenia Foetida (Savigny) 1826. I. Anterior Regeneration" (PDF). Биологический бюллетень. 96 (2): 129–139. Дои:10.2307/1538195. JSTOR 1538195. PMID 18120625. Архивировано из оригинал (PDF) on 2007-04-01.
- ^ Gates, G. E. (1 January 1953). "On Regenerative Capacity of Earthworms of the Family Lumbricidae". Американский натуралист из Мидленда. 50 (2): 414–419. Дои:10.2307/2422100. JSTOR 2422100.
- ^ "Invertebrata 20a items". Архивировано из оригинал на 2006-06-22. Получено 2006-11-20.
- ^ Fender & McKey-Fender (1990). Soil Biology Guide. Wiley-Interscience. ISBN 978-0-471-04551-9.
- ^ а б c Blakemore, R.J. (2006) (March 2006). "Revised Key to Worldwide Earthworm Families from Blakemore (2000) plus Reviews of Criodrilidae (including Biwadrilidae) and Octochaetidae" (PDF). A Series of Searchable Texts on Earthworm Biodiversity, Ecology and Systematics from Various Regions of the World. annelida.net. Получено 15 мая, 2012.
- ^ "Earthworm species name database".
- ^ [ref Blakemore (2016) Cosmopolitan Earthworms]
- ^ Earthworms: Renewers of Agroecosystems (SA Fall, 1990 (v3n1)) В архиве 2007-07-13 на Wayback Machine
- ^ Gould, Edwin; McShea, William; Grand, Theodore (1993). "Function of the Star in the Star-Nosed Mole, Condylura cristata". Журнал маммологии. 74 (1): 108–116. Дои:10.2307/1381909. ISSN 0022-2372. JSTOR 1381909.
- ^ Dejean, A .; Schatx, B. (1999). "Prey Capture Behavior of Psalidomyrmex procerus (Formicidae; Ponerinae), a Specialist Predator of Earthworms (Annelida)". Социобиология: 545–554. ISSN 0361-6525.
- ^ Blakemore, R.J. (2018) (2018). «Критическое сокращение количества дождевых червей органического происхождения в условиях интенсивного сельского хозяйства, истощающего гуминовые органические вещества». Почвенные системы. Soil Systems 2(2): 33. 2 (2): 33. Дои:10.3390 / почвенные системы2020033.
- ^ NSW Department of Primary Industries, How earthworms can help your soil
- ^ Galveston County Master Gardener Association, Beneficials in the garden: #38 Earthworms
- ^ Дарвин, Чарльз (1881). The Formation of Vegetable Mould through the Action of Worms, with Observations on their Habits. Джон Мюррей. Найдено на Project Gutenberg Etext Formation of Vegetable Mould, by Darwin
- ^ Blakemore, R.J. (2007). "Origin and means of dispersal of cosmopolitan Pontodrilus litoralis (Oligochaeta: Megascolecidae)". European Journal of Soil Biology.
- ^ а б c d Nyle C. Brady; Ray R. Weil (2009). Elements of the Nature and Properties of Soils (3-е изд.). Прентис Холл. ISBN 978-0-13-501433-2.
- ^ Mollison, Bill, Permaculture- A Designer's Manual, Tagari Press, 1988
- ^ https://harvardforest.fas.harvard.edu/sites/harvardforest.fas.harvard.edu/files/publications/pdfs/Fisher_Ecology_1928.pdf
- ^ https://www.nrcresearchpress.com/doi/pdf/10.4141/cjss64-005 Some Effects of Earthworm Invasion in Virgin Podzols
- ^ Cooper, Shewell; Soil, Humus And Health ISBN 978-0-583-12796-7
- ^ Blakemore, R.J. (2017) (2017-02-12). "Nature article to commemorate Charles Darwin's birthday on 12th February". VermEcology.
- ^ а б Blanchart, Eric (1992-12-01). "Restoration by earthworms (megascolecidae) of the macroaggregate structure of a destructured savanna soil under field conditions". Биология и биохимия почвы. 24 (12): 1587–1594. Дои:10.1016/0038-0717(92)90155-Q.
- ^ Burke, David (December 26, 2019). "The power of earthworm poop and how it could influence climate change". CBC.
- ^ Blakemore, R.J. (2015). "Eco-taxonomic profile of the iconic vermicomposter - the 'African Nightcrawler', Eudrilus eugeniae (Kinberg, 1867)". African Invertebrates 56: 527-548. Архивировано из оригинал on 2016-10-22.
дальнейшее чтение
- Edwards, Clive A., Bohlen, P.J. (Eds.) Biology and Ecology of Earthworms. Springer, 2005. 3rd edition.
- Edwards, Clive A. (Ed.) Earthworm Ecology. Boca Raton: CRC Press, 2004. Second revised edition. ISBN 0-8493-1819-X
- Lee, Keneth E. Earthworms: Their Ecology and Relationships with Soils and Land Use. Академическая пресса. Sydney, 1985. ISBN 0-12-440860-5
- Стюарт, Эми. The Earth Moved: On the Remarkable Achievements of Earthworms. Chapel Hill, N.C.: Algonquin Books, 2004. ISBN 1-56512-337-9
| Авторитетный контроль |
|---|