Гетероталлизм - Heterothallism - Wikipedia
Гетероталлический разновидность имеют полов которые проживают у разных людей. Этот термин применяется, в частности, для обозначения гетероталлических грибы, которые требуют двух совместимых партнеров для производства половых спор, от гомоталлический те, которые способны половое размножение от единого организма.
У гетероталлических грибов два разных человека вносят свои ядра в зиготу. Примеры гетероталлизма включены для Saccharomyces cerevisiae, Aspergillus fumigatus, Aspergillus flavus, Penicillium marneffei и Neurospora crassa. Гетероталлический жизненный цикл N. crassa приведен довольно подробно, поскольку аналогичные жизненные циклы присутствуют и у других гетероталлических грибов.
Жизненный цикл Saccharomyces cerevisiae
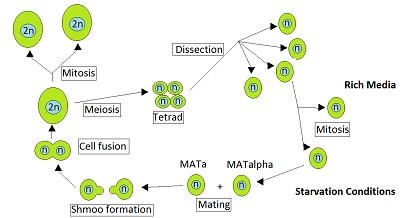
Дрожжи Saccharomyces cerevisiae гетероталлический. Это означает, что каждая дрожжевая клетка относится к определенному типу спаривания и может спариваться только с клеткой другого типа спаривания. Во время вегетативного роста, который обычно происходит при изобилии питательных веществ, С. cerevisiae размножается митозом как гаплоидные или диплоидные клетки. Однако при голодании диплоидные клетки подвергаются мейозу с образованием гаплоидных спор.[1] Спаривание происходит, когда гаплоидные клетки противоположного типа спаривания, MATa и MATα, вступают в контакт. Ruderfer et al.[2] указал, что такие контакты между близкородственными дрожжевыми клетками часты по двум причинам. Во-первых, клетки противоположного типа спаривания присутствуют вместе в одном и том же аск, мешок, содержащий тетрада клеток, непосредственно продуцируемых одним мейоз, и эти клетки могут спариваться друг с другом. Вторая причина состоит в том, что гаплоидные клетки одного типа спаривания при делении клеток часто производят клетки противоположного типа спаривания, с которыми они могут спариваться.
Кац Езов и др.[3] представили доказательства того, что в естественных С. cerevisiae В популяциях преобладают клональное размножение и тип «самооплодотворения» (в форме интратрейд-спаривания). Ruderfer et al.[2] проанализировали происхождение естественных С. cerevisiae штаммов и пришел к выводу, что ауткроссинг происходит примерно один раз на каждые 50 000 делений клеток. Таким образом, хотя С. cerevisiae является гетероталлическим, оказывается, что в природе спаривание чаще всего происходит между близкородственными клетками дрожжей. Относительная редкость мейотических событий, возникающих в результате ауткроссинга, предполагает, что возможные долгосрочные выгоды от ауткроссинга (например, формирование генетическое разнообразие ) вряд ли будет достаточно для сохранения секса в целом от поколения к поколению.[нужна цитата ] Скорее краткосрочная выгода, такая как мейотическая рекомбинационная репарация повреждений ДНК, вызванных стрессовыми условиями, такими как голод, может быть ключом к поддержанию секса в С. cerevisiae.[4][5]
Жизненный цикл Aspergillus fumigatus
Aspergillus fumigatus, представляет собой гетероталлический гриб.[6] Это один из самых распространенных Аспергиллы видов, чтобы вызвать заболевание у людей с иммунодефицит. А. фумигатус, широко распространен в природе и обычно встречается в почве и разлагающихся органических веществах, таких как компостные кучи, где он играет важную роль в углерод и азот переработка отходов. Колонии грибка образуются из конидиеносцы тысячи минут серо-зеленый конидии (2–3 мкм), которые легко переносятся по воздуху. A. fumigatus обладает полностью функциональным половым репродуктивным циклом, который приводит к производству клейстотеция и аскоспоры.[7]
Несмотря на то что А. фумигатус Встречается в районах с сильно различающимся климатом и окружающей средой, имеет низкую генетическую изменчивость и отсутствие генетической дифференциации популяций в глобальном масштабе.[8] Таким образом, способность к гетероталлическому полу сохраняется даже при небольшом генетическом разнообразии. Как и в случае с С. cereviaeКак указано выше, краткосрочная выгода от мейоза может быть ключом к адаптивному поддержанию пола у этого вида.
Жизненный цикл Aspergillus flavus
A. flavus является основным производителем канцерогенный афлатоксины в сельскохозяйственных культурах по всему миру. Это также оппортунистический человек и животное. возбудитель, вызывая аспергиллез у лиц с ослабленным иммунитетом. В 2009 году было обнаружено, что половое состояние этого гетероталлического гриба возникает при совместном культивировании штаммов противоположного типа спаривания в соответствующих условиях.[9]
Сексуальность порождает разнообразие в кластере генов афлатоксина в A. flavus,[10] предполагая, что производство генетической изменчивости может способствовать поддержанию гетероталлизма у этого вида.
Жизненный цикл Talaromyces marneffei
Henk et al.[11] показали, что гены, необходимые для мейоза, присутствуют у T. marneffei, и что спаривание и генетическая рекомбинация происходят у этого вида.
Henk et al.[11] пришли к выводу, что T. marneffei размножается половым путем, но рекомбинация в естественных популяциях, скорее всего, происходит на пространственно и генетически ограниченных расстояниях, что приводит к высококлональной структуре популяции. Пол сохраняется у этого вида, хотя и очень мало генетическая изменчивость производится. Секс может сохраняться в T. marneffei краткосрочным преимуществом мейоза, например С. cerevisiae и А. фумигатус, обсуждалось выше.
Жизненный цикл Neurospora crassa

Половой цикл N. crassa гетероталлический. Половые плодовые тела (перитеции) могут образоваться только при слиянии двух мицелий разного типа спаривания. Как и другие аскомицеты, N. crassa имеет два типа спаривания, которые в данном случае обозначаются буквами «А» и «а». Нет очевидной морфологической разницы между штаммами типа «А» и типа спаривания. Оба могут образовывать обильные протоперитеции, женскую репродуктивную структуру (см. рисунок, верх § ). Протоперитеции легче всего образуются в лаборатории, когда рост происходит на твердой (агаровой) синтетической среде с относительно низким источником азота.[12] По-видимому, азотное голодание необходимо для экспрессии генов, участвующих в половом развитии.[13] Протоперитеций состоит из аскогониума, спиральной многоклеточной гифы, которая заключена в узелковое скопление гиф. Разветвленная система тонких гиф, называемая трихогин, простирается от кончика аскогониума, выступая за покровные гифы в воздух. Половой цикл начинается (т.е. происходит оплодотворение), когда клетка (обычно конидий) противоположного типа спаривания контактирует с частью трихогина (см. рисунок, верх § ). За таким контактом может последовать слияние клеток, ведущее к одному или нескольким ядрам из оплодотворяющей клетки, мигрирующим вниз по трихогину в аскогониум. Поскольку оба штамма «А» и «а» имеют одинаковую половую структуру, ни один из штаммов не может рассматриваться как исключительно мужской или женский. Однако в качестве реципиента протоперитеций штаммов «А» и «а» можно рассматривать как женскую структуру, а оплодотворяющий конидий можно рассматривать как участника мужского пола.
Последующие шаги после слияния гаплоидных клеток «А» и «а» были описаны Fincham и Day,[14] и Вагнером и Митчеллом.[15] После слияния клеток дальнейшее слияние их ядер задерживается. Вместо этого ядро оплодотворяющей клетки и ядро аскогониума связываются и начинают синхронно делиться. Продукты этих ядерных делений (все еще в парах разного типа спаривания, т. Е. "A" / "a") мигрируют в многочисленные аскогенные гифы, которые затем начинают расти из аскогониума. Каждая из этих аскогенных гиф изгибается, образуя крючок (или крючок) на своем конце, и пара гаплоидных ядер «А» и «а» в пределах крозье разделяется синхронно. Затем формируются перегородки, чтобы разделить жердь на три ячейки. Центральная ячейка на изгибе крючка содержит одно ядро «А» и одно ядро «а» (см. рисунок, верх § ). Эта биядерная клетка инициирует образование аска и называется «первичной аскусной» клеткой. Далее два безъядерный клетки по обе стороны от первой аскообразующей клетки сливаются друг с другом, образуя двуядерная клетка которые могут вырасти, чтобы сформировать еще одну клетку, которая затем может сформировать свою собственную начальную клетку аска. Затем этот процесс можно повторять несколько раз.
После образования аскуса-начальной клетки ядра «А» и «а» сливаются друг с другом, образуя диплоидное ядро (см. рисунок, верх § ). Это ядро - единственное диплоидное ядро во всем жизненном цикле N. crassa. Диплоидное ядро состоит из 14 хромосом, образованных из двух слитых гаплоидных ядер, каждое из которых имеет по 7 хромосом. Сразу за образованием диплоидного ядра следует мейоз. Два последовательных деления мейоза приводят к четырем гаплоидным ядрам, двум типу спаривания «А» и двум типу спаривания «а». Одно дальнейшее митотическое деление приводит к четырем ядрам «А» и четырем «а» в каждом. аск. Мейоз является неотъемлемой частью жизненного цикла всех организмов, размножающихся половым путем, и в основных его чертах мейоз в N. crassa кажется типичным для мейоза в целом.
По мере того, как происходят указанные выше события, мицелиальная оболочка, которая окружала аскогоний, развивается как стенка перитеция, пропитывается меланином и чернеет. Зрелый перитеций имеет колбовидное строение.
Зрелый перитеций может содержать до 300 асков, каждый из которых происходит из идентичных диплоидных ядер слияния. Обычно в природе, когда перитеции созревают, аскоспоры довольно сильно выбрасываются в воздух. Эти аскоспоры термостойки и в лаборатории требуют нагревания при 60 ° C в течение 30 минут, чтобы вызвать прорастание. У нормальных штаммов весь половой цикл занимает от 10 до 15 дней. В зрелом аске, содержащем 8 аскоспор, пары соседних спор идентичны по генетической конституции, поскольку последнее деление является митотическим, и поскольку аскоспоры содержатся в мешочке асков, который удерживает их в определенном порядке, определяемом направлением ядерных сегрегаций во время мейоз. Поскольку четыре первичных продукта также расположены последовательно, паттерн генетических маркеров сегрегации первого деления можно отличить от маркеров паттерна сегрегации второго деления.
Смотрите также
Рекомендации
- ^ Херсковиц I (декабрь 1988 г.). «Жизненный цикл зародышевых дрожжей Saccharomyces cerevisiae». Microbiol. Rev. 52 (4): 536–53. Дои:10.1128 / MMBR.52.4.536-553.1988. ЧВК 373162. PMID 3070323.
- ^ а б Рудерфер Д.М., Пратт С.К., Зайдель Х.С., Кругляк Л. (сентябрь 2006 г.). «Популяционный геномный анализ ауткроссинга и рекомбинации у дрожжей». Nat. Genet. 38 (9): 1077–81. Дои:10,1038 / ng1859. PMID 16892060.
- ^ Кац Эзов Т., Чанг С.Л., Френкель З., Сегре А.В., Бахалул М., Мюррей А.В., Лей Дж.Й., Король А., Каши Ю. (январь 2010 г.). «Гетероталлизм в изолятах Saccharomyces cerevisiae от природы: влияние локуса HO на способ размножения». Мол. Ecol. 19 (1): 121–31. Дои:10.1111 / j.1365-294X.2009.04436.x. ЧВК 3892377. PMID 20002587.
- ^ Бердселл Дж. А., Уиллс С. (2003). Эволюционное происхождение и поддержание половой рекомбинации: обзор современных моделей. Серия «Эволюционная биология» >> Эволюционная биология, Vol. 33 с. 27-137. Макинтайр, Росс Дж .; Клегг, Майкл, Т. (ред.), Спрингер. ISBN 978-0306472619
- ^ Эльвира Хёрандл (2013). Мейоз и парадокс секса в природе, Мейоз, ISBN 978-953-51-1197-9, InTech, DOI: 10.5772 / 56542
- ^ Sugui JA, Losada L, Wang W, Varga J, Ngamskulrungroj P, Abu-Asab M, Chang YC, O'Gorman CM, Wickes BL, Nierman WC, Dyer PS, Kwon-Chung KJ (2011). «Идентификация и характеристика пары« суперматер »Aspergillus fumigatus». мБио. 2 (6): e00234–11. Дои:10,1128 / mBio.00234-11. ЧВК 3225970. PMID 22108383.
- ^ О'Горман CM, Фуллер Х., Дайер П.С. (январь 2009 г.). «Открытие полового цикла у условно-патогенного грибка Aspergillus fumigatus». Природа. 457 (7228): 471–4. Bibcode:2009Натура.457..471O. Дои:10.1038 / природа07528. PMID 19043401.
- ^ Rydholm C, Szakacs G, Lutzoni F (апрель 2006 г.). «Низкая генетическая изменчивость и отсутствие обнаруживаемой структуры популяции у aspergillus fumigatus по сравнению с близкородственными видами Neosartorya». Эукариотическая клетка. 5 (4): 650–7. Дои:10.1128 / EC.5.4.650-657.2006. ЧВК 1459663. PMID 16607012.
- ^ Хорн Б.В., Мур Г.Г., Карбон I (2009). «Половое размножение у Aspergillus flavus». Микология. 101 (3): 423–9. Дои:10.3852/09-011. PMID 19537215.
- ^ Мур Г.Г., Эллиотт Дж. Л., Сингх Р., Хорн Б. В., Дорнер Дж. В., Стоун Э. А., Чулз С. Н., Баррос Г. Г., Наик М. К., Райт Г. К., Ад К., Карбон I (2013). «Сексуальность порождает разнообразие в кластере генов афлатоксина: доказательства в глобальном масштабе». PLoS Pathog. 9 (8): e1003574. Дои:10.1371 / journal.ppat.1003574. ЧВК 3757046. PMID 24009506.
- ^ а б Хенк Д.А., Шахар-Голан Р., Деви К.Р., Бойс К.Дж., Жан Н., Федорова Н.Д., Нирман В.С., Сюэ П.Р., Юен К.Ю., Сьё Т.П., Кинь Н.В., Вертхайм Х., Бейкер С.Г., Дэй Дж.Н., Ваниттанаком Н., Бигнелл Е. Андрианопулос А., Фишер М.С. (2012). «Клональность, несмотря на пол: эволюция связанных с хозяином половых соседей в патогенном грибке Penicillium marneffei». PLoS Pathog. 8 (10): e1002851. Дои:10.1371 / journal.ppat.1002851. ЧВК 3464222. PMID 23055919.
- ^ Вестергаард М, Митчелл HK (1947). «Neurospora. Часть V. Синтетическая среда, способствующая половому размножению». Американский журнал ботаники. 34: 573–577.
- ^ Нельсон М.А., Метценберг Р.Л. (сентябрь 1992 г.). «Гены полового развития Neurospora crassa». Генетика. 132 (1): 149–162. ЧВК 1205113. PMID 1356883.
- ^ Fincham J RS, День PR (1963). Грибковая генетика. Оксфорд, Великобритания: Научные публикации Blackwell. КАК В B000W851KO.
- ^ Вагнер Р.П., Митчелл HK (1964). Генетика и метаболизм. Нью-Йорк, Нью-Йорк: Джон Вили и сыновья. КАК В B00BXTC5BO.