Lingulodinium polyedra - Lingulodinium polyedra - Wikipedia
| Lingulodinium polyedra | |
|---|---|
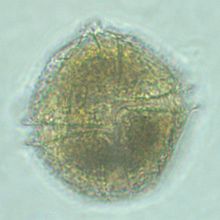 | |
| Вид снизу Lingulodinium polyedrum 900-кратное увеличение | |
| Научная классификация | |
| Домен: | |
| (без рейтинга): | |
| (без рейтинга): | |
| Тип: | |
| Учебный класс: | |
| Заказ: | |
| Семья: | |
| Род: | |
| Разновидность: | L. polyedra |
| Биномиальное имя | |
| Lingulodinium polyedra Додж, 1989 г. | |


Lingulodinium polyedra вид подвижных фотосинтетических динофлагелляты. L. polyedra часто являются причиной красных приливов в южной Калифорнии, что приводит к появлению биолюминесцентных дисплеев на местных пляжах в ночное время.
Жизненный цикл
В рамках своего жизненного цикла этот вид производит стадию покоя, динофлагеллатная киста называется Lingulodinium machaerophorum (синоним Hystrichosphaeridium machaerophorum ). Эта киста была впервые описана Дефландром и Куксоном в 1955 году из миоцена залива Балкомб, Виктория, Австралия, как: «Оболочка шаровидная, субсферическая или эллипсоидальная с жесткой мембраной, более хрупкой, чем деформируемой, покрытой многочисленными длинными, жесткими, коническими, заостренными отростки, напоминающие лезвие кинжала. Поверхность раковины зернистая или пунктированная ».[1] Его стратиграфический диапазон - верхний палеоцен востока США. [2] и Дания [3] до недавнего времени.
Морфология диноцисты с органическими стенками показано, что они контролируются изменениями солености и температуры у некоторых видов, в частности изменением длины процесса (отростки иногда называют шипами, но это неверно, потому что они не обязательно заостренные). Эта морфологическая вариация известна Lingulodinium machaerophorum из культурных экспериментов,[4] и изучение поверхностных отложений.[5]Морфологическое изменение длины процесса может быть использовано для реконструкции соленость. Изменение продолжительности процесса Lingulodinium machaerophorum был использован для реконструкции изменения солености Черного моря.[6]
Токсичность

Lingulodinium polyedra был связан с производством Йессотоксины (YTXs), группа структурно родственных полиэфирных токсинов, которые могут накапливаться в моллюсках и вызывать симптомы, аналогичные симптомам, вызываемым токсинами паралитического отравления моллюсками (PSP).[7]
Люминесценция
Lingulodinium polyedra хорошо видны при 100-кратном увеличении (в большинстве сложных микроскопов используйте 10-кратный или «сканирующий» объектив) и их сцинтиллоны свечение в ответ на поверхностное натяжение и кислотность. Люминесценция регулируется циркадными ритмами и достигает максимума ночью. Из-за этого очевидного ритма (а также из-за того, что большинство его действий, физиологических и молекулярных, ритмичны) L. polyedra был модельным организмом для изучения часов в одиночных клетках.[8]
Рекомендации
- ^ Дефландр, Г. и Куксон, И.С., 1955. Ископаемый микропланктон из отложений позднего мезозоя и третичного периода Австралии. Aust. J. Mar Freshw. Res. 6, 242-313.
- ^ ЭДВАРДС Л.Е., ГУДМАН Д.К., ВИТМЕР Р.Дж. 1984 Биостратиграфия динофлагеллат нижнего третичного периода (группа Паманки), район реки Потомак, Вирджиния и Мэриленд. В: Frederiksen, N.O., and Krafft, K. (eds.), Меловая и третичная стратиграфия, палеонтология и структура, юго-западный Мэриленд и северо-восточная Вирджиния - том и путеводитель. Фонд Американской ассоциации стратиграфических палинологов, Даллас, Техас, с. 137–152.
- ^ HEILMANN-CLAUSEN, C. 1985 Стратиграфия по динофлагеллятам от верховий датского до ипрессского ярусов в скважине Виборг 1, центральная Юлланд, Дания. Danmarks Geologiske Undersøgelse, Серия A, 7: 1–69.
- ^ Hallett, R.I., 1999. Последствия изменения окружающей среды для роста и морфологии Lingulodinium polyedrum (Dinophyceae) в культуре. Кандидат наук. Тезис. Вестминстерский университет, 109 стр.
- ^ Мертенс, К.Н., Рибейро, С. Буиметархан, И., Канер, Х., Комбуриё-Небу, Н. Дейл, Б., де Вернал, А. Эллегаард, М. Филипова, М., Годх, А. Грёсфьельд, К. .Хольцварт, У. Коттофф, У. Лерой, С., Лондекс, Л., Маррет, Ф., Мацуока, К., Муди, П., Наудтс, Л., Пенья-Манджаррес, Дж., Перссон, А. , Попеску, С., Сангиорги, Ф., ван дер Меер, М., Винк, А., Зонневельд, К., Веркаутерен, Д., Влассенброк, Дж., Лувье, С., 2009a. Изменение длины процесса в цистах динофлагелляты, Lingulodinium machaerophorum, в поверхностных отложениях, исследуя его потенциал в качестве индикатора солености. Морская микропалеонтология 70, 54–69.
- ^ Мертенс, К.Н., Брэдли, Л.Р., Такано, Ю., Муди, П.Дж., Маррет, Ф., Аксу, А.Е., Хискотт, Р.Н., Верлей, Т.Дж., Мышин, Е.А., Смирнова, Л.Л., Багери, С., Мансор, М. ., Поспелова, В., Мацуока, К. 2012. Количественная оценка изменения солености поверхности в голоцене в Черном море с использованием длины отростка цисты динофлагеллат. Обзоры четвертичной науки.
- ^ Paz et al. 2008. Йессотоксины, группа морских полиэфирных токсинов: обзор. Mar. Drugs 2008, 6, 73-102; DOI: 10.3390 / md20080005
- ^ Hastings JW. 2008. Часы Gonyaulax в 50: трансляционный контроль циркадной экспрессии. Колд Спринг Харб Symp Quant Biol. 2007; 72: 141-4. DOI: 10.1101 / sqb.2007.72.026