Синаптическая усталость - Synaptic fatigue
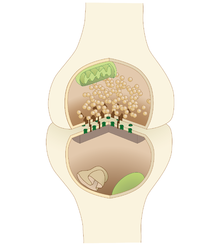
Синаптическая усталость, или краткосрочный синаптическая депрессия, это форма краткосрочного синаптическая пластичность что приводит к временной неспособности нейроны стрелять и, следовательно, передавать входной сигнал. Считается, что это форма негативный отзыв для физиологического контроля определенных форм нервная система Мероприятия.[1]
Это вызвано временным истощением синаптические везикулы этот дом нейротрансмиттеры в синапсе, как правило, вызывается стойкой высокочастотной стимуляцией нейронов. Нейротрансмиттеры высвобождаются синапсом для передачи сигнала постсинаптической клетке. Также была выдвинута гипотеза, что синаптическая усталость может быть результатом постсинаптического рецептор десенсибилизация или изменения постсинаптического пассивного проводимость, но недавние данные свидетельствуют о том, что это прежде всего пресинаптический феномен.[2][3]
Фон
Химические синапсы позволяют передавать сигнал пресинаптической клеткой, высвобождая нейротрансмиттеры в синапс для связывания с рецепторами постсинаптической клетки. Эти нейротрансмиттеры синтезируются в пресинаптической клетке и размещаются в пузырьках до высвобождения. Как только нейротрансмиттеры попадают в синаптическую щель и передается сигнал, повторное поглощение начинается процесс очистки транспортных белков от нейротрансмиттеров из синапса и их рециркуляции, чтобы обеспечить распространение нового сигнала. Если стимуляция происходит с достаточно высокой частотой и достаточной силой, нейротрансмиттеры будут высвобождаться с большей скоростью, чем повторное поглощение может их переработать, что в конечном итоге приведет к их истощению, пока не исчезнут легко высвобождаемые везикулы и сигнал больше не будет передаваться .
Функциональное значение
Ранее было показано, что повторяющиеся короткие поезда потенциалы действия вызывает экспоненциальное затухание амплитуд синаптических ответов в нейронах многих нейронных сетей, в частности ретикулярного ядра каудального моста (PnC). Недавние исследования показали, что только повторная импульсная стимуляция, в отличие от одиночной или парной импульсной стимуляции с очень высокой частотой, может привести к SF.[2] Некоторые клетки, такие как нейроны барорецепторов аорты, могут иметь разрушительные эффекты, включая неспособность регулировать артериальное давление в аорте, если на них повлияет начало синаптической усталости. Активация метаботропных глутаматных ауторецепторов в этих нейронах может подавлять синаптическую передачу, подавляя приток кальция, уменьшая экзоцитоз синаптических везикул и модулируя механизмы, регулирующие восстановление синаптических везикул и эндоцитоз.[4] Эти ауторецепторы глутамата способны подавлять синаптическую усталость, чтобы предотвратить пагубные физиологические последствия, которые могут возникнуть в результате дисфункциональной регуляции артериального давления в аорте (неверно).
Синаптическое восстановление
Когда синаптические везикулы высвобождают в синапс нейротрансмиттеры, которые связываются с белками постсинаптических мембран для передачи сигнала, происходит повторное поглощение нейромедиаторов, чтобы повторно использовать нейротрансмиттеры в пресинаптической клетке для повторного высвобождения. Везикулы нейротрансмиттеров рециркулируются в процессе эндоцитоз. Поскольку каждая пресинаптическая клетка может соединять до тысяч соединений с другими нейронами, синаптическая усталость и ее восстановление могут вызывать взаимодействия с другими нейронными цепями и могут влиять на кинетику с другими процессами нейронов.[5] Важно, чтобы рециклинг нейротрансмиттеров происходил с эффективной и действенной скоростью, чтобы предотвратить негативное влияние синаптической усталости на передачу сигнала.
Время
Поддержание легко высвобождаемого пула везикул важно для обеспечения постоянной способности передавать физиологические сигналы между нейронами. Время, необходимое для того, чтобы нейромедиатор высвободился в синаптическую щель, а затем вернулся в пресинаптическую клетку для повторного использования, в настоящее время недостаточно изучен. В настоящее время предлагаются две модели, чтобы попытаться понять этот процесс. Одна модель предсказывает, что везикула подвергается полному слиянию с пресинаптической клеточной мембраной, когда все ее содержимое опорожняется. Затем он должен извлечь везикулярную мембрану с других участков, что может занять до десятков секунд.[6] Вторая модель пытается объяснить этот феномен, предполагая, что везикулы сразу же начинают перерабатывать нейротрансмиттеры после высвобождения, что занимает меньше секунды для завершения эндоцитоза.[6] Одно исследование показало, что время полного эндоцитоза варьируется от 5,5 до 38,9 секунды. Это также указывало на то, что это время полностью не зависело от длительной или хронической активности.[7]
Пораженные клетки
Синаптическая усталость может повлиять на многие синапсы разных типов нейронов.[5] Существование и наблюдения синаптической усталости признаются повсеместно, хотя точные механизмы, лежащие в основе этого явления, полностью не изучены. Обычно это наблюдается в зрелых клетках при высоких частотах стимулов (> 1 Гц). Одним из конкретных примеров является то, что рефлекс отдергивания жабр Аплизия это вызвано гомосинаптический депрессия.[8] Хотя гомосинаптические и гетеросинаптический депрессия может привести к длительная депрессия и / или потенцирование этот конкретный случай является краткосрочным примером того, как гомосинаптическая депрессия вызывает синаптическую усталость. Перфорантный путь - гранулярные клетки (PP-GC) в зубчатые извилины из гиппокамп у взрослых крыс было показано, что они утомляются при более низких частотах (0,05-0,2 Гц).[9] Было показано, что в развивающихся PP-GC крыс два типа синаптической пластичности приводят к синаптической усталости. Низкочастотная обратимая депрессия высвобождения пресинаптических везикул и форма необратимой депрессии, вызванная: AMPA молчание. Вторая форма пластичности исчезает по мере созревания ПП-ГК, хотя обратимая низкочастотная депрессия остается неизменной.[9]
Роль в нейронной пластичности
Считается, что синаптические везикулы являются частью трех отдельных пулов: легко высвобождаемого пула (составляет примерно 5% от общего количества везикул), рециклирующего пула (около 15%) и резервного пула (оставшиеся 80%).[10] Резервный пул, кажется, начинает выпускать пузырьки только в ответ на интенсивную стимуляцию. Было проведено несколько исследований, которые предполагают, что резервные везикулы редко когда-либо высвобождаются в ответ на физиологические стимулы, что вызывает вопросы об их важности.[10] Это высвобождение в везикулах, независимо от того, из какого пула они высвобождаются, считается формой краткосрочной синаптической пластичности, потому что оно изменяет функциональные характеристики пресинаптической клетки, в конечном итоге временно изменяя ее активирующие свойства. Разница между этим и долгосрочным потенцированием заключается в том, что это явление возникает только в течение времени, необходимого для рециркуляции и повторного использования нейротрансмиттеров, в отличие от долгосрочного, такого как характеристики, лежащие в основе долгосрочного потенцирования. Необходимо провести дальнейшие исследования, чтобы определить важность везикул резервного пула в пресинаптических клетках.
Роль в патологиях ЦНС
Не было доказано, что синаптическая усталость напрямую вызывает или приводит к Центральная нервная система патология, хотя степень его активации в клетках была изучена в результате конкретных патологий и заболеваний. Долгосрочные изменения в нейроне или синапсе, приводящие к постоянному изменению возбуждающих свойств нейрона, могут вызывать синаптическую усталость из-за гораздо большей или меньшей активации, которая потенциально может привести к некоторому виду физиологической аномалии.
Болезнь Альцгеймера
Признаки Болезнь Альцгеймера (AD) - нарушение познания, скопление β-амилоидный пептид (Aβ), нейрофибриллярная дегенерация, потеря нейронов с ускоренным атрофия определенных областей мозга и уменьшение количества синапсов в выживших нейронах. Исследования указывают на пре- и постсинаптические механизмы, приводящие к БА. Одна специфическая аномалия включает повышенное количество пресинаптического белка. ПРИЛОЖЕНИЕ. Было проведено исследование, в котором сравнивали синаптическую усталость у трансгенных мышей, сверхэкспрессирующих APP / PS1, с их однопометниками, у которых не было избыточной экспрессии белка. Результаты показали, что утомляемость была более выражена у мышей APP / PS1, что указывает на уменьшение количества легко высвобождаемых пулов везикул в пресинаптическом нейроне. Выводы этого исследования заключаются в том, что синаптическая усталость является в первую очередь пресинаптическим феноменом и не зависит от постсинаптического воздействия. десенсибилизация рецепторов, синаптическая усталость не является результатом Ca2+ ионы, накапливающиеся в терминале, и, что наиболее важно, синаптическая усталость играет важную роль и может быть изучена при исследовании причин и следствий некоторых нейродегенеративный болезни.[3]
Депрессия
Антидепрессанты имеют краткосрочные и долгосрочные эффекты в подавленный пациенты. Краткосрочные эффекты объясняются гипотезой о том, что депрессия резко вызывается немедленным снижением катехоламины в мозгу. Антидепрессанты немедленно препятствуют этому снижению и восстанавливают нормальные уровни этих нейромедиаторов в головном мозге. В стрессовых условиях везикула экзоцитоз потенцируется, и высвобождение катехоламинов вызывает угнетение пресинаптических клеток из-за истощения нейротрансмиттеров. Лечебные дозы флуоксетин было показано, что они снижают эти состояния утомления нейронов, ингибируя высвобождение везикул и тем самым предотвращая синаптическую усталость в гиппокамп нейроны. Эти результаты показывают, что флуоксетин, а также другие антидепрессанты, которые действуют через те же механизмы, что и флуоксетин, усиливают восстановление нервной системы и нейротрансмиссию, снижая риск депрессии.[11]
Вопросы без ответов
- Хотя сейчас синаптическая усталость считается в первую очередь пресинаптическим феноменом, могут ли постсинаптические процессы составлять большую часть причин, которые в настоящее время понимаются для синаптической усталости?
- Рециркуляция белков мембран синаптических везикул происходит быстро, на что указывает способность многих нейронов активироваться пятьдесят раз в секунду, и весьма специфична, поскольку некоторые мембранные белки, уникальные для синаптических везикул, специфически интернализуются за счет эндоцитоза. Эндоцитоз обычно вовлекает везикулы, покрытые клатрином, хотя могут также использоваться везикулы, не покрытые клатрином. Однако после того, как эндоцитозные везикулы теряют клатриновую оболочку, они обычно не сливаются с более крупными эндосомами с низким pH, как это происходит во время эндоцитоза белков плазматической мембраны в других клетках (см. Рис. 17-46). Скорее, переработанные везикулы немедленно пополняются нейромедиатором.
https://www.ncbi.nlm.nih.gov/books/NBK21521/
Рекомендации
- ^ Килпатрик, З. П. (2010). Пространственно-структурированные волны и колебания в нейронных сетях с синаптической депрессией и адаптацией. Доктор философии, Университет Юты.
- ^ а б Саймонс-Вайденмайер, Н.С., Вебер, М., Плапперт, К.Ф., Пильц, П.К.Д. и Шмид, С. (2006). Синаптическая депрессия и кратковременное привыкание расположены в сенсорной части пути испуга млекопитающих. BMC Neuroscience, 7, 38-38.
- ^ а б Чжан, Х., Гонг, Б., Лю, С., Фа, М., Нинан, И., Станишевский, А., и Арансио, О. (2005). Синаптическая усталость более выражена в модели болезни Альцгеймера у трансгенных мышей APP / PS1. Текущее исследование болезни Альцгеймера, 2(2), 137-140.
- ^ Хэй, М., Хоанг, К. Дж., И Памидимуккала, Дж. (2001). Клеточные механизмы, регулирующие экзоцитоз и эндоцитоз синаптических пузырьков в нейронах барорецепторов аорты. Анналы Нью-Йоркской академии наук, 940, 119-131.
- ^ а б Надим Ф., Мэнор Ю., Копелл Н. и Мардер Э. (1999). Синаптическая депрессия создает переключатель, который контролирует частоту колебательного контура. Слушания Национальной академии наук Соединенных Штатов Америки, 96 (14), 8206-8211.
- ^ а б Райан, Т. А., Смит, С. Дж., И Рейтер, Х. (1996). Сроки эндоцитоза синаптических везикул. Слушания Национальной академии наук Соединенных Штатов Америки, 93 (11), 5567-5571.
- ^ Армбрустер, М., и Райан, Т. А. (2011). Время извлечения синаптических пузырьков является свойством всей клетки, а не индивидуального синапса. [Статья]. Nature Neuroscience, 14 (7), 824-826. Дои:10.1038 / № 2828
- ^ Кастелуччи, В. Ф., и Кандел, Е. Р. (1974). Количественный анализ синаптической депрессии, лежащей в основе привыкания к рефлексу отдергивания жабр при аплизии. Слушания Национальной академии наук Соединенных Штатов Америки, 71 (12), 5004-5008.
- ^ а б Абрахамссон, Т., Густафссон, Б., и Ханс, Э. (2005). Синаптическая усталость в синапсе наивных перфорант-зубчатых гранулярных клеток у крысы. Журнал физиологии, 569 (Pt 3), 737-750.
- ^ а б Икеда, К., и Беккерс, Дж. М. (2009). Подсчет количества высвобождаемых синаптических пузырьков в пресинаптическом окончании. [Статья]. Слушания Национальной академии наук Соединенных Штатов Америки, 106 (8), 2945-2950. Дои:10.1073 / pnas.0811017106
- ^ Хенкель А. В., Вельцель О., Громер Т. В., Трипал П., Роттер А. и Корнхубер Дж. (2010). Флуоксетин предотвращает зависимое от стимуляции утомление экзоцитоза синаптических везикул в нейронах гиппокампа. [Статья]. Журнал нейрохимии, 114 (3), 697-705. Дои:10.1111 / j.1471-4159.2010.06795.x