Подергивание моторики - Twitching motility - Wikipedia
Подергивание моторики это форма ползающей подвижности бактерий, используемая для передвижения по поверхности. Подергивание опосредуется активностью волосообразных нитей, называемых типом IV. пили которые выходят за пределы ячейки, связываются с окружающими твердыми субстратами и втягиваются, вытягивая ячейку вперед аналогично действию якорь.[1][2][3] Название подергивание моторики возникает из-за характерных резких и нерегулярных движений отдельных клеток при просмотре под микроскопом.[4] Он наблюдался у многих видов бактерий, но наиболее хорошо изучен у Синегнойная палочка, Neisseria gonorrhoeae и Myxococcus xanthus. Было показано, что активное движение, опосредованное системой подергивания, является важным компонентом патогенных механизмов нескольких видов.[2]
Механизмы
Структура пилуса
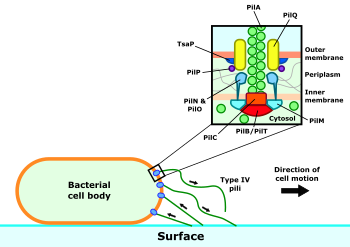
Комплекс пилуса типа IV состоит как из самого пилуса, так и из механизмов, необходимых для его строительства и двигательной активности. Нить пилуса в основном состоит из белка PilA с более редкими второстепенными пилинами на кончике. Считается, что они играют роль в инициировании образования пилуса.[5] В нормальных условиях субъединицы пилина расположены в виде спирали с пятью субъединицами в каждом витке,[5][6] но пили под напряжением способны растягиваться и перестраивать свои субъединицы во вторую конфигурацию с примерно1 2⁄3 субъединиц за ход.[7]
Три подкомплекса образуют аппарат, отвечающий за сборку и втягивание пилей IV типа.[8] Ядром этого механизма является моторный подкомплекс, состоящий из белка PilC и цитозольных АТФаз PilB и PilT. Эти АТФазы управляют расширением или ретракцией пилуса соответственно, в зависимости от того, какая из двух в настоящее время связана с комплексом пилуса. Моторный комплекс окружает подкомплекс выравнивания, образованный из белков PilM, PilN, PilO и PilP. Эти белки образуют мост между внутренней и внешней мембранами и создают связь между моторным субкомплексом внутренней мембраны и субкомплексом секреции внешней мембраны. Он состоит из поры, образованной из белка PilQ, через которую собранный пилус может выйти из клетки.[9]
Регулирование
Регуляторные белки, связанные с системой подергивания движений, имеют сильную последовательность и структурное сходство с белками, которые регулируют бактериальный хемотаксис с помощью жгутиков.[2][10] В P. aeruginosa например, всего четыре гомологичный присутствуют хемосенсорные пути, три из которых регулируют подвижность при плавании и один - при подергивании.[11] Эти хемотаксические системы позволяют клеткам регулировать подергивание, чтобы двигаться к хемоаттрактантам, таким как фосфолипиды и жирные кислоты.[12] Однако в отличие от модели хемотаксиса, связанной с жгутиковыми клетками, движение к хемоаттрактантам в дергающихся клетках, по-видимому, опосредуется посредством регуляции времени изменения направления.[13]
Паттерны подвижности

Подергивание моторики способно управлять движением отдельных клеток.[1][13] Образец подвижности, который возникает в результате, сильно зависит от формы клеток и распределения пилей по поверхности клетки.[14] В N. gonorrhoeae например, примерно сферическая форма клеток и равномерное распределение пилей приводит к тому, что клетки принимают 2D случайная прогулка по поверхности, к которой они прикреплены.[15] Напротив, такие виды, как P. aeruginosa и М. xanthus существуют как удлиненные стержни с пилями, локализованными на их полюсах, и демонстрируют гораздо большую устойчивость в направлении во время ползания из-за возникающего в результате смещения в направлении генерирования силы.[16] P. aeruginosa и М. xanthus также способны изменять направление движения во время ползания, переключая полюс локализации пилуса.[13][14] Пили типа IV также опосредуют форму подвижности при ходьбе у P. aeruginosa, где пили используются для вытягивания стержня клетки в вертикальное положение и перемещения его с гораздо более высокими скоростями, чем во время движения горизонтального ползания.[16][17]
Существование множества пилей, одновременно тянущих к телу клетки, приводит к равновесию сил, определяющих движение тела клетки. Это известно как модель подергивания моторики перетягивания каната.[14][15] Внезапные изменения баланса сил, вызванные отслоением или высвобождением отдельных пилей, приводят к быстрому рывку (или «рогатке»), который сочетает в себе быстрые вращательные и боковые движения, в отличие от более медленных боковых движений, наблюдаемых во время более длительных периодов между выстрелами из рогаток.[18]
Роли
Патогенез
И наличие пилей IV типа, и активное движение ворсинок, по-видимому, вносят важный вклад в патогенность нескольких видов.[8] В P. aeruginosa, потеря ретракции пилуса приводит к снижению бактериальной вирулентности при пневмонии[19] и уменьшает колонизацию роговицы.[20] Некоторые бактерии также способны подергиваться вдоль стенок сосудов против направления потока жидкости внутри них.[21] который, как считается, позволяет колонизировать недоступные иным образом участки сосудистой сети растений и животных.
Бактериальные клетки также могут быть нацелены на подергивание: во время фазы клеточной инвазии в жизненном цикле Bdellovibrio пили типа IV используются клетками для прохождения через щели, образованные в клеточной стенке хищных бактерий.[22] Оказавшись внутри, Bdellovibrio могут использовать ресурсы клетки-хозяина для роста и размножения, в конечном итоге лизируя клеточную стенку бактерии-жертвы и убегая, чтобы вторгнуться в другие клетки.
Биопленки
Подергивание моторики также важно при формировании биопленки.[8] Во время образования и роста биопленки подвижные бактерии могут взаимодействовать с секретируемыми внеклеточные полимерные вещества (ЭПС) такие как Psl, альгинат и внеклеточная ДНК.[23] Так как они сталкиваются с участками высоких отложений EPS, P. aeruginosa клетки замедляются, накапливают и откладывают дополнительные компоненты EPS. Эта положительная обратная связь является важным инициирующим фактором для создания микроколонии, предшественники полноценной биопленки.[24] Вдобавок, как только биопленки сформировались, их распространение, опосредованное подергиванием, облегчается и организуется компонентами EPS.[25]
Подергивание также может влиять на структуру биопленок. Во время своего становления способные к подергиванию клетки способны ползать поверх клеток, лишенных подергивающейся подвижности, и доминировать над быстрорастущей внешней поверхностью биопленки.[23][26]
Таксономическое распространение и эволюция
Пили IV типа и связанные с ними структуры можно найти практически во всех типах Бактерии и Археи,[27] однако окончательная подергивающая подвижность была показана у более ограниченного круга прокариот. Наиболее изученными и широко распространенными являются подергивания протеобактерии, Такие как Neisseria gonorrhoeae, Myxococcus xanthus и Синегнойная палочка.[14][8] Тем не менее, подергивания наблюдались и у других типов. Например, подергивание моторики наблюдается у цианобактерии Synechocystis,[28] а также грамположительный фирмикуты Streptococcus sanguinis.[29]
Другие структуры и системы, тесно связанные с пилями IV типа, также наблюдались у прокариот. В Archea, например, наблюдались пучки нитей типа IV, образующие спиральные структуры, сходные как по форме, так и по функциям с бактериальными жгутик. Эти связанные с плаванием структуры получили название архаэлла.[30] Также тесно связаны с пилусом IV типа система секреции типа II,[31] сам широко распространен среди грамотрицательные бактерии. В этой системе секреции грузы, предназначенные для экспорта, связаны с кончиками псевдопилей типа IV в периплазме. Расширение псевдопилей за счет белков секретина, подобных PilQ, позволяет этим белкам-грузам пересекать внешнюю мембрану и проникать во внеклеточную среду.
Из-за этого широкого, но неоднородного распространения пилообразных механизмов типа IV было высказано предположение, что генетический материал, кодирующий его, передавался между видами через горизонтальный перенос генов после своего первоначального развития у одного вида протеобактерий.[6]
Смотрите также
Рекомендации
- ^ а б Skerker, J.M .; Берг, Х.С. (05.06.2001). «Прямое наблюдение за разгибанием и втягиванием пилей IV типа». Труды Национальной академии наук Соединенных Штатов Америки. 98 (12): 6901–6904. Bibcode:2001PNAS ... 98.6901S. Дои:10.1073 / pnas.121171698. ISSN 0027-8424. ЧВК 34450. PMID 11381130.
- ^ а б c Мэттик, Джон С. (2002). «Пили IV типа и подергивание моторики». Ежегодный обзор микробиологии. 56: 289–314. Дои:10.1146 / annurev.micro.56.012302.160938. ISSN 0066-4227. PMID 12142488.
- ^ Merz, A.J .; Итак, М .; Шитц, М. П. (2000-09-07). «Втягивание пилуса усиливает моторику подергивания бактерий». Природа. 407 (6800): 98–102. Bibcode:2000Натура 407 ... 98М. Дои:10.1038/35024105. ISSN 0028-0836. PMID 10993081.
- ^ Хенрихсен, Дж. (Декабрь 1972 г.). «Бактериальная поверхностная транслокация: обзор и классификация». Бактериологические обзоры. 36 (4): 478–503. ISSN 0005-3678. ЧВК 408329. PMID 4631369.
- ^ а б Leighton, Tiffany L .; Buensuceso, Ryan N.C .; Хауэлл, П. Линн; Берроуз, Лори Л. (01.11.2015). «Биогенез пилей Pseudomonas aeruginosa IV типа и регуляция их функции». Экологическая микробиология. 17 (11): 4148–4163. Дои:10.1111/1462-2920.12849. ISSN 1462-2920. PMID 25808785.
- ^ а б Нудлеман, Эрик; Кайзер, Дейл (2004). «Сближение с пилями IV типа». Журнал молекулярной микробиологии и биотехнологии. 7 (1–2): 52–62. Дои:10.1159/000077869. ISSN 1464-1801. PMID 15170403.
- ^ Биэ, Николас; Хигаши, Дастин Л .; Бруич, Ясна; Итак, Магдалина; Sheetz, Майкл П. (2010-06-22). «Силозависимый полиморфизм пилей IV типа выявляет скрытые эпитопы». Труды Национальной академии наук Соединенных Штатов Америки. 107 (25): 11358–11363. Bibcode:2010PNAS..10711358B. Дои:10.1073 / pnas.0911328107. ISSN 1091-6490. ЧВК 2895099. PMID 20534431.
- ^ а б c d Берроуз, Лори Л. (2012). «Подергивание подергивания Pseudomonas aeruginosa: пили типа IV в действии». Ежегодный обзор микробиологии. 66: 493–520. Дои:10.1146 / annurev-micro-092611-150055. ISSN 1545-3251. PMID 22746331.
- ^ Чанг, И-Вэй; Rettberg, Lee A .; Treuner-Lange, Anke; Иваса, Джанет; Согаард-Андерсен, Лотте; Дженсен, Грант Дж. (2016-03-11). «Архитектура пилуса типа IVa». Наука. 351 (6278): aad2001. Bibcode:2016BpJ ... 110..468C. Дои:10.1126 / science.aad2001. ISSN 1095-9203. ЧВК 5929464. PMID 26965631.
- ^ Сампедро, Инмакулада; Паралес, Ребекка Э .; Крелл, Тино; Хилл, Джейн Э. (январь 2015 г.). «Хемотаксис псевдомонад». Обзор микробиологии FEMS. 39 (1): 17–46. Дои:10.1111/1574-6976.12081. ISSN 1574-6976. PMID 25100612.
- ^ Ortega, Davi R .; Fleetwood, Aaron D .; Крелл, Тино; Харвуд, Кэролайн С.; Дженсен, Грант Дж .; Жулин, Игорь Б. (13.11.2017). «Назначение хеморецепторов на хемосенсорные пути у Pseudomonas aeruginosa». Труды Национальной академии наук Соединенных Штатов Америки. 114 (48): 12809–12814. Дои:10.1073 / pnas.1708842114. ISSN 1091-6490. ЧВК 5715753. PMID 29133402.
- ^ Miller, Rhea M .; Томарас, Андрей П .; Barker, Adam P .; Voelker, Dennis R .; Чан, Эдвард Д .; Василь, Адриана I .; Василь, Михаил Леонидович (01.06.2008). "Хемотаксис, опосредованный подергиванием подвижности Pseudomonas aeruginosa в отношении фосфолипидов и жирных кислот: специфичность и метаболические требования". Журнал бактериологии. 190 (11): 4038–4049. Дои:10.1128 / jb.00129-08. ISSN 0021-9193. ЧВК 2395028. PMID 18390654.
- ^ а б c Oliveira, Nuno M .; Фостер, Кевин Р .; Дарем, Уильям М. (2016-06-07). «Подергивающийся хемотаксис одиночных клеток в развивающихся биопленках». Труды Национальной академии наук. 113 (23): 6532–6537. Дои:10.1073 / pnas.1600760113. ISSN 0027-8424. ЧВК 4988597. PMID 27222583.
- ^ а б c d Майер, Беренике; Вонг, Джерард С. Л. (декабрь 2015 г.). «Как бактерии используют механизмы пили типа IV на поверхности». Тенденции в микробиологии. 23 (12): 775–788. Дои:10.1016 / j.tim.2015.09.002. ISSN 1878-4380. PMID 26497940.
- ^ а б Марат, Рахул; Меил, Клаудия; Schmidt, Nora C .; Девентер, Лена; Курре, Райнер; Грюн, Лило; Шмидт, М. Александр; Мюллер, Мелани Дж. И .; Липовски, Рейнхард (07.05.2014). «Подергивание бактерий координируется двумерным перетягиванием каната с направленной памятью». Nature Communications. 5: 3759. Bibcode:2014НатКо ... 5,3759 млн. Дои:10.1038 / ncomms4759. ISSN 2041-1723. PMID 24806757.
- ^ а б Конрад, Хасинта С .; Гибянский, Максим Л .; Джин, Фан; Гордон, Вернита Д .; Девиз, Dominick A .; Мэтьюсон, Марджи А .; Стопка, Виктор Г .; Zelasko, Daria C .; Shrout, Джошуа Д. (2011-04-06). «Жгутики и пили-опосредованные механизмы приповерхностной одноклеточной подвижности у P. aeruginosa». Биофизический журнал. 100 (7): 1608–1616. Bibcode:2011BpJ ... 100.1608C. Дои:10.1016 / j.bpj.2011.02.020. ISSN 1542-0086. ЧВК 3072661. PMID 21463573.
- ^ Гибянский, Максим Л .; Конрад, Хасинта С .; Джин, Фан; Гордон, Вернита Д .; Девиз, Dominick A .; Мэтьюсон, Марджи А .; Стопка, Виктор Г .; Zelasko, Daria C .; Шраут, Джошуа Д. (08.10.2010). «Бактерии используют пили типа IV, чтобы ходить вертикально и отделяться от поверхностей». Наука. 330 (6001): 197. Bibcode:2010Sci ... 330..197G. Дои:10.1126 / science.1194238. HDL:2152/39116. ISSN 1095-9203. PMID 20929769.
- ^ Джин, Фан; Конрад, Хасинта С .; Гибянский, Максим Л .; Вонг, Джерард С. Л. (2011-08-02). «Бактерии используют пили типа IV для стрельбы из рогатки по поверхности». Труды Национальной академии наук Соединенных Штатов Америки. 108 (31): 12617–12622. Дои:10.1073 / pnas.1105073108. ISSN 1091-6490. ЧВК 3150923. PMID 21768344.
- ^ Comolli, J.C .; Hauser, A.R .; Waite, L .; Whitchurch, C.B .; Mattick, J. S .; Энгель, Дж. Н. (июль 1999 г.). «Продукты гена Pseudomonas aeruginosa PilT и PilU необходимы для цитотоксичности in vitro и вирулентности на мышиной модели острой пневмонии». Инфекция и иммунитет. 67 (7): 3625–3630. ISSN 0019-9567. ЧВК 116553. PMID 10377148.
- ^ Золфагар, Ирандохт; Эванс, Дэвид Дж .; Флейзиг, Сюзанна М. Дж. (01.09.2003). «Подергивание подвижности способствует роли пилей в инфекции роговицы, вызванной синегнойной палочкой». Инфекция и иммунитет. 71 (9): 5389–5393. Дои:10.1128 / iai.71.9.5389-5393.2003. ISSN 0019-9567. ЧВК 187331. PMID 12933890.
- ^ Шен, Йи; Сириапорн, Альберт; Лекюер, Сиголен; Гитай, Земер; Стоун, Ховард А. (2012-07-03). «Поток побуждает прикрепленные к поверхности бактерии двигаться вверх по течению». Биофизический журнал. 103 (1): 146–151. Bibcode:2012BpJ ... 103..146S. Дои:10.1016 / j.bpj.2012.05.045. ISSN 1542-0086. ЧВК 3388212. PMID 22828341.
- ^ Сокет, Рене Элизабет (2009). «Хищный образ жизни Bdellovibrio bacteriovorus». Ежегодный обзор микробиологии. 63: 523–539. Дои:10.1146 / annurev.micro.091208.073346. ISSN 1545-3251. PMID 19575566.
- ^ а б Parsek, Matthew R .; Толкер-Нильсен, Тим (декабрь 2008 г.). «Формирование паттернов в биопленках Pseudomonas aeruginosa». Текущее мнение в микробиологии. 11 (6): 560–566. Дои:10.1016 / j.mib.2008.09.015. ISSN 1879-0364. PMID 18935979.
- ^ Чжао, Кунь; Ценг, Бу Шань; Бекерман, Бернард; Джин, Фан; Гибянский, Максим Л .; Харрисон, Джо Дж .; Луйтен, Эрик; Parsek, Matthew R .; Вонг, Джерард С. Л. (2013-05-16). «Следы Psl направляют исследования и формирование микроколоний в биопленках Pseudomonas aeruginosa». Природа. 497 (7449): 388–391. Bibcode:2013Натура.497..388Z. Дои:10.1038 / природа12155. ISSN 1476-4687. ЧВК 4109411. PMID 23657259.
- ^ Gloag, Erin S .; Тернбулл, Линн; Хуанг, Алан; Валлотон, Паскаль; Ван, Хуабинь; Нолан, Лаура М .; Милилли, Лиза; Хант, Кэмерон; Лу, Цзин (09.07.2013). «Самоорганизации бактериальных биопленок способствует внеклеточная ДНК». Труды Национальной академии наук Соединенных Штатов Америки. 110 (28): 11541–11546. Bibcode:2013ПНАС..11011541Г. Дои:10.1073 / pnas.1218898110. ISSN 1091-6490. ЧВК 3710876. PMID 23798445.
- ^ Клаузен, Миккель; Ааес-Йоргенсен, Андерс; Молин, Сорен; Толкер-Нильсен, Тим (01.10.2003). «Участие миграции бактерий в развитии сложных многоклеточных структур в биопленках Pseudomonas aeruginosa». Молекулярная микробиология. 50 (1): 61–68. Дои:10.1046 / j.1365-2958.2003.03677.x. ISSN 1365-2958. PMID 14507363.
- ^ Берри, Джейми-Ли; Пеличич, Владимир (январь 2015). «Чрезвычайно распространенные наномашины, состоящие из пилинов типа IV: прокариотические ножи швейцарской армии». Обзор микробиологии FEMS. 39 (1): 134–154. Дои:10.1093 / femsre / fuu001. ISSN 1574-6976. ЧВК 4471445. PMID 25793961.
- ^ Bhaya, D .; Bianco, N.R .; Bryant, D .; Гроссман, А. (август 2000 г.). «Биогенез и подвижность пилуса IV типа в цианобактериях Synechocystis sp. PCC6803». Молекулярная микробиология. 37 (4): 941–951. Дои:10.1046 / j.1365-2958.2000.02068.x. ISSN 0950-382X. PMID 10972813.
- ^ Гурунг, Ишвори; Спилман, Ингрид; Дэвис, Марк Р .; Лала, Раджан; Гаустад, Питер; Биэ, Николас; Пеличич, Владимир (01.01.2016). «Функциональный анализ необычной пилуса IV типа на грамположительный Streptococcus sanguinis». Молекулярная микробиология. 99 (2): 380–392. Дои:10.1111 / мм. 13237. ISSN 1365-2958. ЧВК 4832360. PMID 26435398.
- ^ Ng, Sandy Y.M .; Чабан, Бонни; Джаррелл, Кен Ф. (2006). «Жгутики архей, бактериальные жгутики и пили типа IV: сравнение генов и посттрансляционных модификаций». Журнал молекулярной микробиологии и биотехнологии. 11 (3–5): 167–191. Дои:10.1159/000094053. ISSN 1464-1801. PMID 16983194.
- ^ Пибоди, Кристофер Р .; Чунг, Ён Джун; Йен, Мин-Рен; Видаль-Ингильярди, Доминик; Пагсли, Энтони П .; Сайер, Милтон Х. (ноябрь 2003 г.). «Секреция белка типа II и ее связь с бактериальными пилями IV типа и жгутиками архей». Микробиология. 149 (Pt 11): 3051–3072. Дои:10.1099 / мик.0.26364-0. ISSN 1350-0872. PMID 14600218.