Базилярная мембрана - Basilar membrane - Wikipedia
| Базилярная мембрана. | |
|---|---|
 Раздел через кортиевый орган, показывая базилярную мембрану | |
 Поперечный разрез улитки. | |
| Подробности | |
| Идентификаторы | |
| латинский | мембрана базиларис протока кохлеарис |
| MeSH | D001489 |
| Анатомическая терминология | |
В базилярная мембрана является жестким структурным элементом внутри улитка из внутреннее ухо который разделяет две заполненные жидкостью трубки, идущие вдоль спирали улитки, scala media и scala tympani. Базилярная мембрана перемещается вверх и вниз в ответ на входящие звуковые волны, которые преобразуются в бегущие волны на базилярной мембране.
Структура
Базилярная мембрана - это псевдорезонансная структура.[1] которые, как и струны на инструменте, различаются по ширине и жесткости. Но в отличие от параллельных струн гитары, базилярная мембрана представляет собой единую структуру с разной шириной, жесткостью, массой, демпфированием и размерами каналов в разных точках по длине. Движение базилярной мембраны обычно описывается как бегущая волна.[2] Свойства мембраны в данной точке по длине определяют ее характеристическую частоту (CF), частоту, на которой она наиболее чувствительна к звуковым колебаниям. Базилярная мембрана наиболее широкая (0,42–0,65 мм) и наименее жесткая на вершине улитки, а наиболее узкая (0,08–0,16 мм) и жесткая - у основания (около круглого и овального окон).[3] Высокочастотные звуки локализуются около основания улитки, а низкочастотные звуки - около верхушки.
Функция

Разделение эндолимфы / перилимфы
Вместе с вестибулярная перепонка, несколько тканей, удерживаемых базилярной мембраной, разделяют жидкости эндолимфа и перилимфа, такие как внутренние и внешние клетки борозды (показаны желтым) и ретикулярная пластинка кортиевого органа (показаны пурпурным цветом). Для кортиевого органа базилярная мембрана проницаема для перилимфы. Здесь граница между эндолимфой и перилимфой проходит на ретикулярной пластинке, на стороне эндолимфы орган Корти.[6]
А основание для сенсорных клеток
Базилярная мембрана также является основание для волосковые клетки. Эта функция присутствует у всех наземных позвоночных. Благодаря своему расположению базилярная мембрана размещает волосковые клетки рядом как с эндолимфой, так и с перилимфой, что является предварительным условием функционирования волосковых клеток.
Частотная дисперсия
Третья, эволюционно более молодая, функция базилярной мембраны сильно развита в улитке большинства видов млекопитающих и слабо развита у некоторых видов птиц:[7] то разброс входящих звуковых волн для пространственного разделения частот. Вкратце, мембрана сужается и с одного конца жестче, чем с другого. Более того, звуковые волны, идущие к «более гибкому» концу базилярной мембраны, должны проходить через более длинный столб жидкости, чем звуковые волны, идущие к более близкому и более жесткому концу. Таким образом, каждую часть базилярной мембраны вместе с окружающей жидкостью можно рассматривать как систему «масса-пружина» с различными резонансными свойствами: высокой жесткостью и малой массой, следовательно, высокими резонансными частотами на ближнем (базовом) конце и низкая жесткость и большая масса, следовательно, низкие резонансные частоты на дальнем (вершинном) конце.[8] Это приводит к тому, что входящий звук определенной частоты вызывает вибрацию в одних местах мембраны сильнее, чем в других. Распределение частот по местам называется тонотопической организацией улитки.
Звуковые колебания распространяются как волны вдоль этой мембраны, вдоль которой у людей лежит около 3500 внутренние волосковые клетки расположены в один ряд. Каждая ячейка прикреплена к крохотной треугольной рамке. «Волосы» - это крошечные отростки на конце клетки, очень чувствительные к движению. Когда вибрация мембраны сотрясает треугольные рамки, волоски на клетках многократно смещаются, и это создает потоки соответствующих импульсов в нервных волокнах, которые передаются в слуховой проход.[9] В внешние волосковые клетки обратная энергия для усиления бегущей волны до 65 дБ в некоторых местах.[10][11] В мембране наружных волосковых клеток есть моторные белки, связанные с мембраной. Эти белки активируются вызванными звуком рецепторными потенциалами, когда базилярная мембрана перемещается вверх и вниз. Эти моторные белки могут усиливать движение, заставляя базилярную мембрану двигаться немного больше, усиливая бегущую волну. Следовательно, внутренние волосковые клетки получают большее смещение своих ресничек, двигаются немного больше и получают больше информации, чем в пассивной улитке.
Генерирование рецепторного потенциала
Движение базилярной мембраны вызывает волосковая клетка движение стереоцилий. Волосковые клетки прикреплены к базилярной мембране, и при перемещении базилярной мембраны движутся текториальная мембрана и волосковые клетки, причем стереоцилии изгибаются вместе с относительным движением текториальной мембраны. Это может вызвать открытие и закрытие механически закрытых калиевых каналов на ресничках волосковой клетки. Реснички волосковой клетки находятся в эндолимфа. В отличие от нормального клеточного раствора с низкой концентрацией калия и высоким содержанием натрия, в эндолимфе высокая концентрация калия и низкая концентрация натрия. И он изолирован, что означает, что он не имеет потенциала покоя -70 мВ по сравнению с другими нормальными клетками, а скорее поддерживает потенциал около +80 мВ. Однако основание волосковой клетки находится в перилимфа, с потенциалом 0 мВ. Это приводит к тому, что волосковая клетка имеет потенциал покоя -45 мВ. Когда базилярная мембрана движется вверх, реснички движутся в направлении, вызывающем открытие механически закрытого калиевого канала. Приток ионов калия приводит к деполяризации. Напротив, реснички перемещаются в другую сторону по мере того, как базилярная мембрана перемещается вниз, закрывая более механически управляемые калиевые каналы и приводя к гиперполяризации. Деполяризация откроет потенциалзависимый кальциевый канал, высвобождая нейротрансмиттер (глутамат) в нервном окончании, воздействуя на спиральные ганглиозные клетки, первичные слуховые нейроны, повышая вероятность их возникновения. Гиперполяризация вызывает меньший приток кальция, следовательно, меньшее высвобождение нейротрансмиттеров и снижение вероятности всплеска спиральных ганглиозных клеток.
Дополнительные изображения

Схематический продольный разрез улитки.
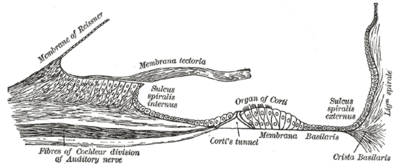
Пол улиткового протока.
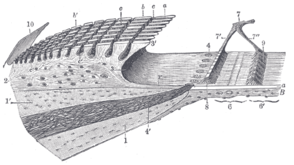
Спиральный лимб и базилярная мембрана.

Разрез спирального органа Кортиева (увеличено)
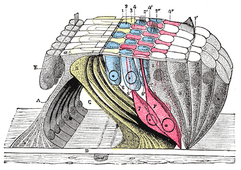
В ретикулярная мембрана и нижележащие конструкции.




Смотрите также
Рекомендации
- ^ Холмс М, Коул Дж.С. (1983). «Псевдорезонанс в улитке». In de Boer E, Viergever MA (ред.). Механика слуха. Делфт: Материалы симпозиума IUTAM / ICA. С. 45–52.
- ^ Фэй Р.Р., Поппер А.Н., Бэкон С.П. (2004). Компрессия: от улитки до кохлеарных имплантатов. Springer. ISBN 0-387-00496-3.
- ^ Oghalai JS (октябрь 2004 г.). «Кохлеарный усилитель: увеличение бегущей волны во внутреннем ухе». Текущее мнение в области отоларингологии и хирургии головы и шеи. 12 (5): 431–8. Дои:10.1097 / 01.moo.0000134449.05454.82. ЧВК 1315292. PMID 15377957.
- ^ Shera CA (ноябрь 2007 г.). "Лазерное усиление с поворотом: распространение бегущей волны и функции усиления по всей улитке". Журнал акустического общества Америки. 122 (5): 2738–58. Bibcode:2007ASAJ..122.2738S. Дои:10.1121/1.2783205. PMID 18189566. Архивировано из оригинал 3 июля 2013 г.
- ^ Роблес Л., Руджеро М.А. (июль 2001 г.). «Механика улитки млекопитающих». Физиологические обзоры. 81 (3): 1305–52. Дои:10.1152 / Physrev.2001.81.3.1305. ЧВК 3590856. PMID 11427697.
- ^ Соль А.Н., Кониши Т. (1986). «Улитковые жидкости: перилимфа и эндолимфа». В Altschuler RA, Hoffman DW, Bobbin RP (ред.). Нейробиология слуха: улитка. Нью-Йорк: Raven Press. С. 109–122.
- ^ Fritzsch B: Переход от воды к суше: эволюция базилярного сосочка четвероногих; среднее ухо и слуховые ядра. В: Вебстер Д. Б., Фэй Р. А., Поппер А. Н., ред. (1992). Эволюционная биология слуха. Берлин: Springer-Verlag. С. 351–375. ISBN 0-387-97588-8.
- ^ Шнупп Дж, Нелькен I, Король А (2011). Слуховая неврология. Кембридж, Массачусетс: MIT Press. ISBN 978-0-262-11318-2.
- ^ Бимент Дж (2001). «Как мы слышим музыку: взаимосвязь между музыкой и механизмом слуха». Вудбридж: Бойделл Пресс: 97. Цитировать журнал требует
| журнал =(помощь) - ^ Нильсен К.Е., Рассел И.Дж. (июль 1999 г.). «Время кохлеарной обратной связи: пространственное и временное представление тона через базилярную мембрану». Природа Неврология. 2 (7): 642–8. Дои:10.1038/10197. PMID 10404197. S2CID 2380374.
- ^ Нильсен К.Е., Рассел И.Дж. (октябрь 2000 г.). «Пространственное и временное представление тона на базилярной мембране морской свинки». Труды Национальной академии наук Соединенных Штатов Америки. 97 (22): 11751–8. Bibcode:2000PNAS ... 9711751N. Дои:10.1073 / pnas.97.22.11751. ЧВК 34345. PMID 11050205.
внешняя ссылка
- Слуховая неврология | Ухо несколько анимаций, показывающих движение базилярной мембраны при различных условиях стимула
- Функциональная анатомия внутреннего уха: множество изображений, анимаций и очень кратких функциональных объяснений
- Симулятор основной мембраны Видео и сценарии для моделирования основной мембраны
- Роль базилярной мембраны в звуковой рецепции: хорошее объяснение и схемы