Обнаружение совпадений в нейробиологии - Coincidence detection in neurobiology
Эта статья поднимает множество проблем. Пожалуйста помоги Улучши это или обсудите эти вопросы на страница обсуждения. (Узнайте, как и когда удалить эти сообщения-шаблоны) (Узнайте, как и когда удалить этот шаблон сообщения)
|
Обнаружение совпадений в контексте нейробиологии это процесс, посредством которого нейрон или нейронная цепь может кодировать информацию, обнаруживая появление близких по времени, но пространственно распределенных входных сигналов. Детекторы совпадений влияют на обработку нейронной информации, уменьшая временные дрожь,[1] снижение спонтанной активности и формирование ассоциаций между отдельными нейронными событиями. Эта концепция привела к большему пониманию нейронных процессов и формированию вычислительных карт в мозге.
Принципы обнаружения совпадений
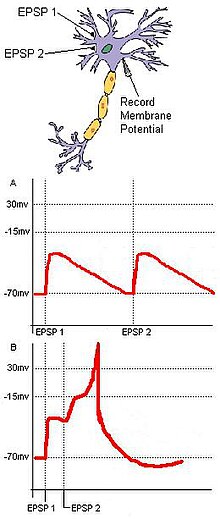
Обнаружение совпадений полагается на отдельные входные данные, сходящиеся к общей цели. Рассмотрим базовую нейронную цепь с двумя входными нейронами, A и B, которые имеют возбуждающие синаптические терминалы, сходящиеся к одному выходному нейрону C (рис. 1). Если каждый входной нейрон EPSP является подпороговым для потенциал действия на C, то C не сработает, если два входа от A и B не будут временно близки друг к другу. Синхронное поступление этих двух входов может подтолкнуть мембранный потенциал целевого нейрона выше порога, необходимого для создания потенциала действия. Если два входа поступают слишком далеко друг от друга, деполяризация первого входа может иметь время значительно снизиться, не позволяя мембранному потенциалу целевого нейрона достичь порога потенциала действия. Этот пример включает в себя принципы пространственный и временное суммирование. Кроме того, обнаружение совпадений может уменьшить дрожание, вызванное спонтанной активностью. В то время как случайные подпороговые стимуляции нейронными клетками не часто могут срабатывать случайно, совпадающие синаптические входы, полученные от единого внешнего стимула, будут гарантировать, что целевой нейрон срабатывает в результате стимула.
Обнаружение дистального совпадения
Приведенное выше описание хорошо применимо к входным сигналам с прямой связью в нейроны, которые обеспечивают входные данные либо от сенсорных нервов, либо от низкоуровневых областей мозга. Однако около 90% межнейронных связей носят не прямой, а прогнозирующий (или модулирующий, или привлекающий внимание) характер. Эти соединения получают входные данные в основном от соседних ячеек в том же слое, что и принимающая ячейка, а также от удаленных подключений, которые проходят через Уровень 1. Дендриты, которые получают эти входные данные, довольно удалены от тела ячейки, и поэтому они демонстрируют разные электрические и электрические характеристики. поведение обработки сигналов по сравнению с проксимальными (или прямолинейными) дендритами, описанными выше.
В коротком разделе (возможно, 40мкм long) дистального дендрита, реакция на активации, поступающие от синапсов к дендритным шипам, увеличивает общий локальный потенциал с каждым входящим сигналом. Этот растущий потенциал действует на фоне спада потенциала обратно на уровень покоя. Если достаточное количество сигналов получено в течение короткого периода времени (то есть до того, как общее напряжение упадет до фонового значения), напряжение сегмента поднимется выше порогового значения, вызывая нелинейный дендритный всплеск, который распространяется, фактически не уменьшаясь, все путь к телу клетки, что приводит к его частичной деполяризации.
Это, пожалуй, самая важная форма обнаружения совпадений дендритов в головном мозге. Более понятная проксимальная активация действует в течение гораздо более длительных периодов времени и, таким образом, гораздо менее чувствительна к временному фактору при обнаружении совпадений.
Звуковая локализация

Было показано, что обнаружение совпадений является основным фактором звуковая локализация вдоль азимут самолет в нескольких организмах. В 1948 г. Ллойд А. Джеффресс предположили, что некоторые организмы могут иметь набор нейронов, которые получают звуковой сигнал от каждого уха.[2] Нейронные пути к этим нейронам называются линиями задержки. Джеффресс утверждал, что нейроны, с которыми связаны линии задержки, действуют как детекторы совпадений, максимально срабатывая при получении одновременных сигналов от обоих ушей. Когда звук слышен, звуковые волны могут достигать ушей в разное время. Это называется межуральная разница во времени (ITD). Из-за разной длины и конечной скорости проводимости в аксонах линий задержки разные нейроны детектора совпадений срабатывают, когда звук исходит из разных положений по азимуту. Модель Джеффресса предполагает, что два сигнала даже от асинхронного поступления звука в улитку каждого уха будут синхронно сходиться на детекторе совпадений в слуховой коре в зависимости от величины ITD (рис. 2). Следовательно, ITD должен соответствовать анатомической карте, которую можно найти в головном мозге. Масакадзу Кониси учиться на сипухи показывает, что это правда.[3] Сенсорная информация от волосковые клетки ушной раковины переходит к ипсилатеральному магноцеллюлярному ядру. Отсюда сигналы проецируются ипсилатерально и контралатерально к двум ламинарным ядрам. Каждое ламинарное ядро содержит детекторы совпадений, которые получают слуховой сигнал от левого и правого уха. Поскольку ипсилатеральные аксоны входят в ламинарное ядро дорсально, а контралатеральные аксоны входят вентрально, звуки из разных положений по азимуту напрямую соответствуют стимуляции ламинарного ядра разной глубины. На основе этой информации была сформирована нейронная карта слухового пространства. Функция ламинарного ядра аналогична функции ламинарного ядра. медиальная верхняя олива у млекопитающих.[4]
Синаптическая пластичность и ассоциативность
В 1949 г. Дональд Хебб постулировали, что синаптическая эффективность будет увеличиваться за счет многократной и постоянной стимуляции постсинаптической клетки пресинаптической клеткой. Это часто неофициально резюмируется как "ячейки, которые срабатывают вместе, соединяются вместе". Теория была частично подтверждена открытием долгосрочное потенцирование. Исследования LTP на множественных пресинаптических клетках, стимулирующих постсинаптическую клетку, открыли свойство ассоциативности. Слабая нейрональная стимуляция пирамидный нейрон не может вызвать долгосрочное потенцирование. Однако та же самая стимуляция в паре с одновременной сильной стимуляцией от другого нейрона усилит оба синапса.[5] Этот процесс предполагает, что два нейрональных пути, сходящиеся к одной и той же клетке, могут усилиться, если стимулировать их по совпадению.
Молекулярный механизм долговременной потенциации
LTP в гиппокамп требует длительной деполяризации, которая может вытеснить Mg2+ блок постсинаптического Рецепторы NMDA. Удаление Mg2+ блок позволяет поток Ca2+ в камеру. Активируется большое повышение уровня кальция протеинкиназы которые в конечном итоге увеличивают количество постсинаптических Рецепторы AMPA. Это увеличивает чувствительность постсинаптической клетки к глутамат. В результате укрепляются оба синапса. Длительная деполяризация, необходимая для изгнания Mg2+ от рецепторов NMDA требует высокочастотной стимуляции.[6] Ассоциативность становится фактором, потому что это может быть достигнуто за счет двух одновременных входов, которые могут быть недостаточно сильными, чтобы активировать LTP сами по себе.
Помимо процессов, основанных на NMDA-рецепторах, другие клеточные механизмы позволяют установить связь между двумя разными входными сигналами, сходящимися на одном нейроне, в определенный период времени. При одновременном повышении внутриклеточных концентраций цАМФ и Са2+, коактиватор транскрипции под названием TORC1 (CRTC1 ) активируется, что преобразует временное совпадение двух вторые мессенджеры в долгосрочные изменения, такие как LTP.[7] Этот клеточный механизм через кальций-зависимый аденилатциклаза активация, может также учитывать обнаружение повторяющейся стимуляции данного синапс.
Аденилилциклаза (также широко известная как аденилциклаза и аденилатциклаза) был вовлечен в формирование памяти как детектор совпадений.[8][9][10][11]
Молекулярный механизм длительной депрессии
Длительная депрессия также работает через ассоциативные свойства, хотя это не всегда процесс, обратный ДП. LTD в мозжечок требует одновременной стимуляции параллельные волокна и лазание по волокнам. Глутамат, высвобождаемый из параллельных волокон, активирует рецепторы AMPA, которые деполяризуют постсинаптическую клетку. Параллельные волокна также активируют метаботропный рецепторы глутамата, которые высвобождают вторичные мессенджеры IP3 и DAG. Волокна для лазания стимулируют значительное увеличение постсинаптического Ca2+ уровни при активации. CA2+, IP3, и DAG работать вместе в путь передачи сигнала для интернализации рецепторов AMPA и снижения чувствительности постсинаптических клеток к глутамату.[6]
Смотрите также
Рекомендации
- ^ Marsálek, P .; Koch, C .; Маунселл, Дж. (1997). «О связи между синаптическим входом и джиттером спайкового выхода в отдельных нейронах». Труды Национальной академии наук Соединенных Штатов Америки. 94 (2): 735–740. Дои:10.1073 / пнас.94.2.735. ЧВК 19583. PMID 9012854.
- ^ Джеффресс, Л. А. (1948). «Теория места локализации звука». Журнал сравнительной и физиологической психологии. 41 (1): 35–39. Дои:10,1037 / ч0061495. PMID 18904764.
- ^ Carr, C.E .; Кониши, М. (1988). «Линии аксональной задержки для измерения времени в стволе мозга совы». Труды Национальной академии наук Соединенных Штатов Америки. 85 (21): 8311–8315. Дои:10.1073 / pnas.85.21.8311. ЧВК 282419. PMID 3186725.
- ^ Zupanc, G.K.H. 2004. Поведенческая нейробиология: интегративный подход. Издательство Оксфордского университета: Оксфорд, Великобритания. стр.133-150
- ^ Фрей, Уве; Моррис, Ричард Г. М. (февраль 1997 г.). «Синаптическая маркировка и долгосрочное потенцирование». Природа. 385 (6616): 533–536. Дои:10.1038 / 385533a0. ISSN 0028-0836. PMID 9020359.
- ^ а б Purves, Дейл (2004). Неврология (3-е изд.). Сандерленд, Массачусетс: Sinauer Associates. С. 575–608. ISBN 9780878937257.
- ^ Ковач, К. А .; Steullet, P .; Steinmann, M .; Do, K. Q .; Magistretti, P.J .; Halfon, O .; Кардино, Ж. -Р. (2007). «TORC1 - это кальций- и цАМФ-чувствительный детектор совпадений, участвующий в долговременной синаптической пластичности гиппокампа». Труды Национальной академии наук. 104 (11): 4700–4705. Дои:10.1073 / pnas.0607524104. ЧВК 1838663. PMID 17360587.
- ^ Уиллоби, Дебби; Купер, Дермот М. Ф. (июль 2007 г.). «Организация и регуляция Са2 + аденилилциклаз в микродоменах цАМФ». Физиологические обзоры. 87 (3): 965–1010. CiteSeerX 10.1.1.336.3746. Дои:10.1152 / Physrev.00049.2006. ISSN 0031-9333. PMID 17615394.
- ^ Mons, N .; Guillou, J.-L .; Джаффард, Р. (1999-04-01). «Роль Ca2 + / кальмодулин-стимулирующих аденилилциклаз как детекторов молекулярных совпадений в формировании памяти». Клеточные и молекулярные науки о жизни. 55 (4): 525–533. Дои:10.1007 / с000180050311. ISSN 1420-682X. PMID 10357223.
- ^ Hanoune, J .; Дефер, Н. (2001). «Регуляция и роль изоформ аденилатциклазы». Ежегодный обзор фармакологии и токсикологии. 41: 145–174. Дои:10.1146 / annurev.pharmtox.41.1.145. ISSN 0362-1642. PMID 11264454.
- ^ Neve, Kim A .; Seamans, Джереми К .; Трантам-Дэвидсон, Хизер (август 2004 г.). «Передача сигналов дофаминового рецептора». Журнал исследований рецепторов и передачи сигналов. 24 (3): 165–205. CiteSeerX 10.1.1.465.5011. Дои:10.1081 / RRS-200029981. ISSN 1079-9893. PMID 15521361.
дальнейшее чтение
- Бендер, В. А .; Бендер, К. Дж .; Brasier, D.J .; Фельдман Д. Э. (2006). «Два детектора совпадения для зависимой от времени пластичности спайков в соматосенсорной коре». Журнал неврологии. 26 (16): 4166–4177. Дои:10.1523 / JNEUROSCI.0176-06.2006. ЧВК 3071735. PMID 16624937.
- Caillard, O .; Бен-Ари, Й .; Гайарса, Дж. Л. (1999). «Долгосрочное усиление ГАМКергической синаптической передачи в гиппокампе новорожденных крыс». Журнал физиологии. 518 (Pt 1): 109–119. Дои:10.1111 / j.1469-7793.1999.0109r.x. ЧВК 2269393. PMID 10373693.
- Joris, P. X .; Smith, P.H .; Инь, Т. С. (1998). «Обнаружение совпадений в слуховой системе: 50 лет после Джеффресса». Нейрон. 21 (6): 1235–1238. Дои:10.1016 / S0896-6273 (00) 80643-1. PMID 9883717.
- https://web.archive.org/web/20040519194818/http://bbsonline.cup.cam.ac.uk/Preprints/OldArchive/bbs.neur4.crepel.html
внешняя ссылка
- Слуховая локализация с помощью ITD-анализа: модель Джеффресса - Интерактивное онлайн-руководство (Flash)