Переворот оснований ДНК - DNA base flipping - Wikipedia
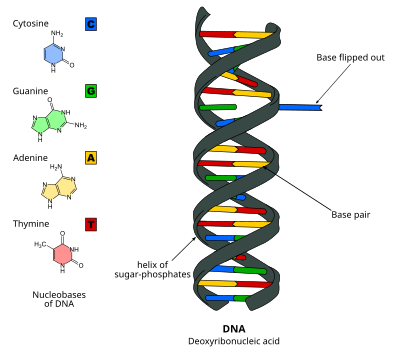
Переворот основания ДНК, или же переворот нуклеотидов, это механизм, в котором один нуклеотид база, или азотистое основание, вращается вне двойной спирали нуклеиновой кислоты.[1] Это происходит, когда процессинг нуклеиновой кислоты фермент необходим доступ к основанию для выполнения работ с ним, таких как его удаление для замены на другую основу во время Ремонт ДНК. Впервые наблюдался в 1994 г. Рентгеновская кристаллография в метилтрансфераза фермент, катализирующий метилирование цитозин основание в ДНК. С тех пор было показано, что он используется различными ферментами во многих биологических процессах, таких как Метилирование ДНК, разные Ремонт ДНК механизмы и Репликация ДНК. Это также может происходить в двойных спиралях РНК. [2] или в ДНК: промежуточные соединения РНК, образующиеся во время Транскрипция РНК.
Переворачивание оснований ДНК происходит при нарушении водородные связи между базами и разделение базы от соседей. Это может происходить через активный процесс, когда фермент связывается с ДНК и затем способствует вращению основания, или пассивный процесс, когда основание вращается самопроизвольно, и это состояние распознается и связывается ферментом. Его можно обнаружить с помощью Рентгеновская кристаллография, ЯМР-спектроскопия, флуоресцентная спектроскопия, или же зонды гибридизации.
Открытие
Переворот базы впервые был обнаружен в 1994 году, когда исследователи Климасаускас, Кумар, Робертс и Ченг использовали Рентгеновская кристаллография для просмотра промежуточного этапа химической реакции метилтрансфераза привязан к ДНК.[3] Метилтрансфераза, которую они использовали, была C5-цитозинметилтрансфераза из Haemophilus haemolyticus (М. HhaI). Этот фермент распознает определенную последовательность ДНК (5'-GCGC-3 ') и метилирует первую цитозин основание последовательности в ее местоположении C5.[3] После кристаллизации комплекса M. HhaI-ДНК они увидели, что целевое цитозиновое основание полностью развернулось из двойной спирали и расположилось в активном центре M. HhaI. Это поддерживалось многочисленными взаимодействиями между M. HhaI и ДНК.[3]
Авторы предположили, что переворот оснований был механизмом, используемым многими другими ферментами, такими как геликасы, рекомбинация ферменты, РНК-полимеразы, ДНК-полимеразы, и Топоизомеразы типа II.[3] За годы, последовавшие за этим открытием, было проведено много исследований, и было обнаружено, что переворот оснований - это механизм, используемый во многих биологических процессах, предлагаемых авторами.[4][5][6]
Механизм
ДНК нуклеотиды удерживаются вместе с водородные связи, которые относительно слабы и легко ломаются. Базовое переключение происходит в миллисекундном масштабе времени[7] путем разрыва водородных связей между основаниями и отделения основания от его соседей.[8] Основание повернуто из двойная спираль на 180 град.,[9] обычно через большая канавка,[10] и в активный центр фермента. Это открытие приводит к небольшим конформационным изменениям в основной цепи ДНК.[11] которые быстро стабилизируются за счет усиления взаимодействия фермент-ДНК.[12] Исследования, посвященные свободная энергия профили переворота основания показали, что барьер свободной энергии для переворота может быть снижен на 17 ккал / моль для M.HhaI в закрытом конформация.[10]
Существует два механизма переворота оснований ДНК: активный и пассивный.[13] В активном механизме фермент связывается с ДНК, а затем активно вращает основание, в то время как в пассивном механизме поврежденное основание сначала спонтанно вращается, а затем распознается и связывается ферментом.[8] Исследования продемонстрировали оба механизма: урацил-ДНК гликозилаза следует пассивному механизму[8] и Tn10 транспозаза следует за активным механизмом.[14]
Более того, исследования показали, что переворачивание оснований ДНК используется многими различными ферментами в различных биологических процессах, таких как Метилирование ДНК, разные Ремонт ДНК механизмы, Транскрипция РНК и Репликация ДНК.[4][5][6]
Биологические процессы
Модификация и ремонт ДНК

ДНК может иметь мутации которые вызывают повреждение основания в цепи ДНК. Чтобы гарантировать генетическую целостность ДНК, ферменты должны восстанавливать любые повреждения. Есть много видов Ремонт ДНК. Базовая эксцизионная пластика использует переворот основания, чтобы вытащить поврежденное основание из двойной спирали[5] и в карман специфики гликозилаза который гидролизует гликозидную связь и удаляет основание.[15] ДНК-гликозилазы взаимодействуют с ДНК, меняя основания, чтобы определить несоответствие. Пример эксцизионной репарации оснований происходит, когда основание цитозина дезаминируется и становится основанием урацила. Это вызывает неправильную пару U: G, которая обнаруживается Урацил ДНК гликозилаза. Основание урацила переворачивается в активный карман гликозилазы, где оно удаляется из цепи ДНК.[16] Переворот базы используется для исправления таких мутаций, как 8-оксогуанин (oxoG)[17] и димеры тимина создается УФ-излучением.[15][18]
Репликация, транскрипция и рекомбинация
Репликация ДНК и Транскрипция РНК оба используют базовое переворачивание.[5] ДНК-полимераза это фермент, выполняющий репликацию. Его можно рассматривать как руку, которая сжимает однонитевую матрицу ДНК.[15] Когда шаблон проходит через область ладони полимеразы, основания шаблона выворачиваются из спирали в сторону от dNTP сайт привязки.[19] Во время транскрипции РНК-полимераза катализирует синтез РНК. Вовремя инициация фаза, две базы в -10 элемент вывернуть из спирали в два кармана в РНК-полимеразе. Эти новые взаимодействия стабилизируют элемент -10 и способствуют разделению или плавлению цепей ДНК.[15][20]
Переворот базы происходит на последних этапах рекомбинация.[21] RecA белок, который способствует нить вторжения[15] в течение гомологичная рекомбинация. Переворот базы был предложен в качестве механизма, с помощью которого RecA может позволить одной нити распознавать гомология в дуплексной ДНК.[22] Другие исследования показывают, что он также участвует в V (D) J Рекомбинация.[23]
Метилирование ДНК

Метилирование ДНК это процесс, в котором метильная группа добавляется либо к цитозин или же аденин.[24] Этот процесс вызывает активацию или деактивацию экспрессия гена, тем самым приводя к регуляции генов в эукариотических клетках. Также известно, что процесс метилирования ДНК участвует в определенных типах рак формирование.[25][26][27] Для того, чтобы эта химическая модификация произошла, необходимо, чтобы основание-мишень выворачивалось из двойной спирали ДНК, чтобы позволить метилтрансферазам катализировать реакцию.[5]
Распознавание мишеней эндонуклеазами рестрикции
Эндонуклеазы рестрикции, также известные как рестрикционные ферменты являются ферментами, которые расщепляют сахарно-фосфатную основу ДНК на конкретных нуклеотиды последовательности, которые обычно имеют длину от четырех до шести нуклеотидов.[28] Исследования, проведенные Хортоном и его коллегами, показали, что механизм, с помощью которого эти ферменты расщепляют ДНК, включает в себя переворачивание оснований, а также изгиб ДНК и расширение малая бороздка.[29] В 2006 году Хортон и его коллеги, рентгеновская кристаллография Были представлены доказательства, показывающие, что рестрикционная эндонуклеаза HinP1I использует переворот оснований для распознавания своей целевой последовательности. Этот фермент как известно, расщепляет ДНК в палиндромный тетрануклеотидная последовательность G ↓ CGC.
Экспериментальные подходы к обнаружению
Рентгеновская кристаллография

Рентгеновская кристаллография это метод измерения углов и интенсивности кристаллические атомы для определения атомной и молекулярной структуры интересующего кристалла. После этого кристаллографы могут создать трехмерное изображение, на котором положение атомы, химические связи а также другие важные характеристики.[30] Климасаукас и его коллеги использовали эту технику для наблюдения за первым феноменом переворота базы, в котором их экспериментальная процедура включала несколько этапов:[3]
- Очищение
- Кристаллизация
- Сбор информации
- Определение и уточнение структуры
Во время очистки Haemophilus haemolyticus метилтрансфераза был чрезмерно выраженный и очищают с использованием стадии обратной экстракции с высоким содержанием соли для селективной солюбилизации M.HhaI с последующей быстрой жидкостной хроматографией белка (FPLC ), как это сделали ранее Кумар и его коллеги.[31] Авторы использовали анионообменную колонку Mono-Q для удаления небольшого количества белковый материалы и нежелательная ДНК до стадии кристаллизации. После успешной очистки M.HhaI образец был затем выращен с использованием метода, который смешивает раствор, содержащий комплекс, при температуре 16 ° C и метод диффузии пара висячей капли для получения кристаллов. Затем авторы смогли собрать рентгеновские данные в соответствии с техникой, которую использовали Ченг и его коллеги в 1993 году.[32] Этот метод включал измерение интенсивности дифракции на детекторе FAST, где время экспозиции для вращения на 0,1 ° составляло 5 или 10 секунд. Для определения и уточнения структуры Климасаукас и его коллеги использовали молекулярную замену уточненной структуры апо, описанную Ченгом и его коллегами в 1993 г.[32] где поиск моделей X-PLOR, MERLOT, и TRNSUM использовались для решения функций вращения и сдвига.[33][34] Эта часть исследования включает использование разнообразного программного обеспечения и компьютерных алгоритмов для определения структур и характеристик интересующего кристалла.
ЯМР-спектроскопия
ЯМР-спектроскопия это техника, которая использовалась на протяжении многих лет для изучения важных динамических аспектов переворота базы. Этот метод позволяет исследователям определять физические и химические свойства атомы и другие молекулы, используя магнитные свойства атомные ядра.[35] Кроме того, ЯМР может предоставить различную информацию, включая структуру, состояния реакции, химическое окружение молекул и динамика.[36][37] Во время эксперимента по открытию переворота оснований ДНК исследователи использовали ЯМР-спектроскопию для изучения индуцированного ферментом переворота оснований метилтрансферазы HhaI. Для проведения этого эксперимента два 5-фторцитозин остатки были включены в мишень и ссылочную позицию с субстратом ДНК, так что 19F может быть выполнен анализ химического сдвига. Однажды 19F Был проведен анализ химического сдвига, затем был сделан вывод о существовании комплексов ДНК с множественными формами целевого 5-фторцитозина по пути переворота оснований.[38]
Флуоресцентная спектроскопия
Флуоресцентная спектроскопия это метод, который используется для анализа образца с помощью флуоресцентного зонда. Сами нуклеотиды ДНК не являются хорошими кандидатами для этого метода, потому что они с трудом переизлучают свет при возбуждении светом.[39] Флуоресцентный маркер необходим для обнаружения переворота основания. 2-аминопурин основа, структурно похожая на аденин, но очень флуоресцирует при выходе из дуплекса ДНК.[40] Он обычно используется для обнаружения переворота оснований и имеет возбуждение при 305–320 нм и излучение при 370 нм, так что он хорошо отделен от возбуждений белков и ДНК. Другие флуоресцентные зонды, используемые для изучения переворота оснований ДНК: 6КАРТА (4 ‑ амино ‑ 6 ‑ метил ‑ 7 (8H) ‑ птеридон)[41] и Pyrrolo ‑ C (3- [β-D-2-рибофуранозил] -6-метилпирроло [2,3-d] пиримидин-2 (3H) -он).[42][43] Флуоресцентная спектроскопия с временным разрешением также используется для получения более подробной картины степени переворота основания, а также конформационной динамики, происходящей во время переворота основания.[44]
Гибридизационное зондирование
Зонды для гибридизации может использоваться для обнаружения переворота базы. В этом методе используется молекула, которая имеет последовательность, комплементарную последовательности, которую вы хотели бы обнаружить, так что она связывается с однонитевой ДНК или РНК. Несколько зондов гибридизации были использованы для обнаружения переворота оснований. Перманганат калия используется для обнаружения тимин остатки, которые были сброшены цитозин-C5 и аденин-N6 метилтрансферазы.[45] Хлорацетальдегид используется для обнаружения цитозин остатки перевернуты ДНК цитозин-5-метилтрансферазой HhaI (M. HhaI).[46]

Смотрите также
- Ремонт ДНК
- Базовая эксцизионная пластика
- Репликация ДНК
- Транскрипция РНК
- Метилирование ДНК
- ДНК-метилтрансфераза
- Генетическая рекомбинация
- Гомологичная рекомбинация
- ДНК
- Эпигенетика
- Эпигеномика
Рекомендации
- ^ Робертс, RJ; Ченг, X (1998). "Базовое переворачивание". Ежегодный обзор биохимии. 67 (1): 181–198. Дои:10.1146 / annurev.biochem.67.1.181. PMID 9759487.
- ^ Райтер, штат Нью-Джерси; Blad, H; Abildgaard, F; Мясник, SE (2004). «Динамика внутримолекулярной стебель-петли РНК U6: конформационное изменение переворота основания». Биохимия. 43 (43): 13739–47. Дои:10.1021 / bi048815y. PMID 15504036. S2CID 25391616.
- ^ а б c d е Климасаускас, Саулюс; Кумар, Санджай; Робертс, Ричард Дж .; Чэн, Сяодун (январь 1994 г.). «Хал-метилтрансфераза переворачивает целевое основание из спирали ДНК». Клетка. 76 (2): 357–369. Дои:10.1016/0092-8674(94)90342-5. PMID 8293469.
- ^ а б Браун, Том. "Книга нуклеиновых кислот". Получено 26 февраля 2014.
- ^ а б c d е Хуанг, Ниу; Нилеш К. Банавали; Александр Д. МакКерелл (7 января 2003 г.). «Облегченное белком переворачивание оснований в ДНК цитозин-5-метилтрансферазой». PNAS. 100 (1): 68–73. Bibcode:2003ПНАС..100 ... 68Н. Дои:10.1073 / pnas.0135427100. ЧВК 140885. PMID 12506195.
- ^ а б Грубмюллер, Гельмут. "Перевертывание базы ДНК". Получено 26 февраля 2014.
- ^ Бувье, Бенджамин; Грубмюллер, Гельмут (август 2007 г.). «Исследование молекулярной динамики медленного переворота оснований в ДНК с использованием конформационного затопления» (PDF). Биофизический журнал. 93 (3): 770–786. Bibcode:2007BpJ .... 93..770B. Дои:10.1529 / biophysj.106.091751. ЧВК 1913169. PMID 17496048.
- ^ а б c Ларивьер, Л. (23 июня 2004 г.). «Структурные доказательства пассивного механизма переключения оснований для глюкозилтрансферазы». Журнал биологической химии. 279 (33): 34715–34720. Дои:10.1074 / jbc.M404394200. PMID 15178685.
- ^ Грожан, [отредактированный] Анри (2009). Ферменты модификации ДНК и РНК: структура, механизм, функции и эволюция. Остин, Техас: Landes Bioscience. ISBN 978-1-58706-329-9. Архивировано из оригинал на 2014-04-07. Получено 2014-03-10.CS1 maint: дополнительный текст: список авторов (связь)
- ^ а б Huang, N .; Banavali, N.K .; МакКерелл, А. Д. (27 декабря 2002 г.). «Облегченное белком переворачивание оснований в ДНК цитозин-5-метилтрансферазой». Труды Национальной академии наук. 100 (1): 68–73. Дои:10.1073 / pnas.0135427100. ЧВК 140885. PMID 12506195.
- ^ Джудиче, Э. (1 марта 2003 г.). «Открытие пары оснований в B-ДНК: пути свободной энергии для пар GC и AT из моделирования зонтичного отбора проб». Исследования нуклеиновых кислот. 31 (5): 1434–1443. Дои:10.1093 / нар / gkg239. ЧВК 149832. PMID 12595551. Получено 15 марта 2014.
- ^ Huang, N .; Banavali, N.K .; МакКерелл, А. Д. (27 декабря 2002 г.). «Облегченное белком переворачивание оснований в ДНК цитозин-5-метилтрансферазой». Труды Национальной академии наук. 100 (1): 68–73. Дои:10.1073 / pnas.0135427100. ЧВК 140885. PMID 12506195.
- ^ О'Нил, Лорен. Перевернутое основание в ДНК: обнаружение, структуры и энергия, Диссертация. ISBN 9780549590743. Получено 15 марта 2014.
- ^ Bischerour, Julien; Чалмерс, Рональд; Белинский, Аня-Катрин (10 июля 2009 г.). «Переворот базы в транспозиции Tn10: активный механизм переворота и захвата». PLOS ONE. 4 (7): e6201. Bibcode:2009PLoSO ... 4.6201B. Дои:10.1371 / journal.pone.0006201. ЧВК 2705183. PMID 19593448.
- ^ а б c d е Университет, Джеймс Д. Уотсон, Лаборатория Колд-Спринг-Харбор, Таня А. Бейкер, Массачусетский технологический институт, Александр Ганн, Лаборатория Колд-Спринг-Харбор, Майкл Левин, Калифорнийский университет, Беркли, Ричард Лосик, Гарвард (2014). Молекулярная биология гена (Седьмое изд.). Бостон: Пирсон / CSH Press. ISBN 978-0-321-76243-6.
- ^ Krokan, Hans E; Драблёш, Финн; Слуппхауг, Гейр (16 декабря 2002 г.). «Урацил в ДНК - возникновение, последствия и восстановление». Онкоген. 21 (58): 8935–8948. Дои:10.1038 / sj.onc.1205996. PMID 12483510.
- ^ Банерджи, Анирбан; Ян, Вэй; Карплюс, Мартин; Вердин, Грегори Л. (31 марта 2005 г.). «Структура фермента репарации, исследующего неповрежденную ДНК, проясняет распознавание поврежденной ДНК». Природа. 434 (7033): 612–618. Bibcode:2005Натура.434..612Б. Дои:10.1038 / природа03458. PMID 15800616.
- ^ Фуксрайтер, Моника; Ло, Нин; Едловски, Пал; Саймон, Иштван; Осман, Роман (ноябрь 2002 г.). «Роль переворота оснований в специфическом распознавании поврежденной ДНК восстанавливающими ферментами». Журнал молекулярной биологии. 323 (5): 823–834. Дои:10.1016 / S0022-2836 (02) 00999-3. PMID 12417196.
- ^ Patel, Premal H .; Сузуки, Мотоши; Адман, Элинор; Синкай, Акео; Лоэб, Лоуренс А. (май 2001 г.). «Прокариотическая ДНК-полимераза I: эволюция, структура и механизм« переворота оснований »для выбора нуклеотидов». Журнал молекулярной биологии. 308 (5): 823–837. Дои:10.1006 / jmbi.2001.4619. PMID 11352575. S2CID 16277925.
- ^ Lim, H.M .; Lee, H.J .; Рой, С .; Адхья, С. (4 декабря 2001 г.). «Мастер» в разрыве оснований во время изомеризации промотора после связывания РНК-полимеразы ». Труды Национальной академии наук. 98 (26): 14849–14852. Bibcode:2001ПНАС ... 9814849Л. Дои:10.1073 / pnas.261517398. ЧВК 64947. PMID 11734629.
- ^ Волошин, Олег Н .; Камерини-Отеро, Р. Даниэль (сентябрь 2004 г.). «Возвращение к синаптическому комплексу». Молекулярная клетка. 15 (6): 846–847. Дои:10.1016 / j.molcel.2004.09.010. PMID 15383274.
- ^ Фолта-Стогнев, E; О'Мэлли, S; Gupta, R; Андерсон, KS; Раддинг, CM (24 сентября 2004 г.). «Обмен парами оснований ДНК, совпадающий с распознаванием гомологии, обеспечиваемой белком RecA E. coli». Молекулярная клетка. 15 (6): 965–75. Дои:10.1016 / j.molcel.2004.08.017. PMID 15383285.
- ^ Bischerour, J .; Lu, C .; Roth, D. B .; Чалмерс, Р. (31 августа 2009 г.). «Перевернутая основа в рекомбинации V (D) J: понимание механизма образования шпильки, правила 12/23 и координации двунитевых разрывов». Молекулярная и клеточная биология. 29 (21): 5889–5899. Дои:10.1128 / MCB.00187-09. ЧВК 2772739. PMID 19720743.
- ^ Клозе, Роберт Дж .; Адриан П. Берд (2006). «Метилирование геномной ДНК: метка и ее медиаторы». Тенденции в биохимических науках. 31 (2): 89–97. Дои:10.1016 / j.tibs.2005.12.008. ISSN 0968-0004. PMID 16403636.
- ^ Накао, М. (2001). «Эпигенетика: взаимодействие метилирования ДНК и хроматина». Ген. 278 (1–2): 25–31. Дои:10.1016 / s0378-1119 (01) 00721-1. PMID 11707319.
- ^ Plass, C; Soloway, PD (2002). «Метилирование ДНК, импринтинг и рак». Eur J Hum Genet. 10 (1): 6–16. Дои:10.1038 / sj.ejhg.5200768. PMID 11896451.
- ^ Эстеллер, М; Герман, JG (2002). «Рак как эпигенетическое заболевание: метилирование ДНК и изменения хроматина в опухолях человека». Джей Патол. 196 (1): 1–7. Дои:10.1002 / путь.1024. PMID 11748635.
- ^ «Архивная копия». Архивировано из оригинал на 2014-04-18. Получено 2014-04-03.CS1 maint: заархивированная копия как заголовок (связь)
- ^ Хортон, Джон Р .; Чжан, Син; Маунус, Роберт; Ян, Чжэ; Уилсон, Джеффри; Робертс, Ричард; Чэн, Сяодун (2006). "Раскалывание ДНК эндонуклеазой HinP1I: изгиб, переворот основания и расширение незначительной бороздки". Исследования нуклеиновых кислот. 34 (3): 939–948. Дои:10.1093 / нар / gkj484. ЧВК 1363774. PMID 16473850.
- ^ Рентгеновская кристаллография
- ^ Кумар, S; Ченг, X; Pflugrath, JW; Робертс, Р.Дж. (1992). «Очистка, кристаллизация и предварительный рентгеноструктурный анализ комплекса M.HhaI-AdoMet». Биохимия. 31 (36): 8648–8653. Дои:10.1021 / bi00151a035. PMID 1390649.
- ^ а б Ченг, X; Кумар, S; Posfai, J; Pflugrath, JW; Робертс, Р.Дж. (1993). «Кристаллическая структура ДНК-метилтрансферазы HhaI в комплексе с S-аденозил-L-метионином». Клетка. 74 (2): 299–307. Дои:10.1016 / 0092-8674 (93) 90421-л. PMID 8343957.
- ^ Брюнгер А.Т. (1992) «X-PLOR, версия 3.1: система для рентгеновской кристаллографии и ЯМР» (Нью-Хейвен, Коннектикут: Yale University Press)
- ^ Фитцджеральд, П.М.Д. (1988). «MERLOT, интегральный пакет компьютерных программ для определения кристаллической структуры путем молекулярного замещения». J. Appl. Кристаллогр. 21 (3): 273–288. Дои:10.1107 / s0021889887012299.
- ^ ЯМР-спектроскопия
- ^ Герон М. и Дж. Л. Лерой. 1995. Исследование кинетики пар оснований с помощью ЯМР-измерения протонного обмена. В ядерном магнитном резонансе и нуклеиновых кислотах. Academic Press, Сан-Диего, Калифорния.
- ^ Leijon, M .; Граслунд, А. (1992). «Влияние последовательности и длины на кинетику имино-протонного обмена и раскрытия пары оснований в дуплексах олигонуклеотидов ДНК». Нуклеиновые кислоты Res. 20 (20): 5339–5343. Дои:10.1093 / nar / 20.20.5339. ЧВК 334339. PMID 1331987.
- ^ Климасаукас, Салиус и Зита Люткевичуте. «Экспериментальные подходы к изучению смещения оснований ДНК». Ферменты модификации ДНК и РНК: структура, механизм, функция и эволюция. Landes Bioscience, 2009. 37-50. Интернет. 16 марта 2014 г. <https://www.landesbioscience.com/pdf/04GrosjeanKlimasauskas.pdf В архиве 2014-04-07 в Wayback Machine >.
- ^ Грожан, [отредактированный] Анри (2009). Ферменты модификации ДНК и РНК: структура, механизм, функции и эволюция (PDF). Остин, Техас: Landes Bioscience. п. 43. ISBN 978-1-58706-329-9.CS1 maint: дополнительный текст: список авторов (связь)
- ^ Хольц, Б. (15 февраля 1998 г.). «2-Аминопурин как флуоресцентный зонд для переворота оснований ДНК метилтрансферазами». Исследования нуклеиновых кислот. 26 (4): 1076–1083. Дои:10.1093 / nar / 26.4.1076. ЧВК 147370. PMID 9461471.
- ^ Ян, К; Мацика, С; Стэнли, RJ (6 сентября 2007 г.). «6MAP, флуоресцентный аналог аденина, представляет собой пробу переворота оснований ДНК-фотолиазой». Журнал физической химии B. 111 (35): 10615–25. Дои:10.1021 / jp071035p. PMID 17696385. S2CID 4998287.
- ^ Ян, К; Стэнли, Р. Дж. (Май – июнь 2008 г.). «Степень деформации ДНК в комплексах ДНК-фотолиаза-субстрат: исследование флуоресценции состояния раствора». Фотохимия и фотобиология. 84 (3): 741–9. Дои:10.1111 / j.1751-1097.2007.00251.x. PMID 18086248.
- ^ Берри, Дэвид А .; Юнг, Ки-Йонг; Wise, Dean S .; Sercel, Anthony D .; Пирсон, Уильям Х .; Маки, Хью; Рэндольф, Джон Б.; Сомерс, Роберт Л. (март 2004 г.). «Пирроло-dC и пирроло-C: флуоресцентные аналоги цитидина и 2'-дезоксицитидина для исследования олигонуклеотидов». Буквы Тетраэдра. 45 (11): 2457–2461. Дои:10.1016 / j.tetlet.2004.01.108.
- ^ Neely, R.K .; Tamulaitis, G .; Chen, K .; Кубала, М .; Сиксныс, В .; Джонс, А. К. (8 сентября 2009 г.). «Флуоресцентные исследования с временным разрешением для переворота нуклеотидов рестрикционными ферментами». Исследования нуклеиновых кислот. 37 (20): 6859–6870. Дои:10.1093 / nar / gkp688. ЧВК 2777440. PMID 19740769.
- ^ Serva, S (1 августа 1998 г.). «Химическое отображение остатков тимина, нарушенных ДНК-метилтрансферазами». Исследования нуклеиновых кислот. 26 (15): 3473–3479. Дои:10.1093 / nar / 26.15.3473. ЧВК 147733. PMID 9671807.
- ^ Daujotyte, D .; Liutkeviciute, Z .; Tamulaitis, G .; Климасаускас, С. (15 апреля 2008 г.). «Химическое картирование цитозинов, ферментативно вывернутых из спирали ДНК». Исследования нуклеиновых кислот. 36 (10): e57. Дои:10.1093 / nar / gkn200. ЧВК 2425465. PMID 18450817.