Метилтрансфераза - Methyltransferase

Метилтрансферазы представляют собой большую группу ферментов, которые все метилат их субстраты, но их можно разделить на несколько подклассов в зависимости от их структурных особенностей. Самый распространенный класс метилтрансфераз - это класс I, каждый из которых содержит Россманн фолд для связывания S-Аденозилметионин (СЭМ). Метилтрансферазы класса II содержат домен SET, который представлен доменом SET. гистоновые метилтрансферазы и метилтрансферазы класса III, ассоциированные с мембраной.[1] Метилтрансферазы также можно разделить на разные типы, использующие разные субстраты в реакциях переноса метила. Эти типы включают белковые метилтрансферазы, ДНК / РНК-метилтрансферазы, натуральный продукт метилтрансферазы и независящие от SAM метилтрансферазы. SAM является классическим донором метила для метилтрасфераз, однако в природе встречаются примеры других доноров метила. Общий механизм переноса метила - это SN2 -подобная нуклеофильная атака, где метионин сера служит нуклеофил который передает метильную группу субстрату фермента. SAM конвертируется в S-Аденозил гомоцистеин (SAH) во время этого процесса. Разрыв SAM-метильной связи и образование метильной связи субстрат происходят почти одновременно. Эти ферментативные реакции обнаруживаются во многих путях и связаны с генетическими заболеваниями, раком и нарушениями обмена веществ. Другой тип переноса метила - это радикал S-аденозилметионин (SAM), который представляет собой метилирование неактивированных атомов углерода в первичных метаболитах, белках, липидах и РНК.
Функция
Генетика
Метилирование, а также другие эпигенетический модификации, влияет транскрипция, стабильность гена и родительский импринтинг.[2] Это напрямую влияет хроматин структура и может модулировать транскрипцию гена или даже полностью тишина или же активировать гены, без мутация к самому гену. Хотя механизмы этого генетического контроля сложны, гипо- и гиперметилирование ДНК связано со многими заболеваниями.
Белковая регуляция
Метилирование белков играет регулирующую роль в белок-белковые взаимодействия, белок-ДНК взаимодействия, и активация белка.
Примеры:RCC1, важно митотический белок, метилирован, поэтому он может взаимодействовать с центромеры хромосом. Это пример регуляции белок-белкового взаимодействия, поскольку метилирование регулирует прикрепление RCC1 к гистоновым белкам. H2A и H2B. Взаимодействие RCC1-хроматин также является примером взаимодействия белок-ДНК, поскольку другой домен RCC1 взаимодействует непосредственно с ДНК, когда этот белок метилирован. Когда RCC1 не метилирован, делящиеся клетки имеют несколько полюса шпинделя и обычно не может выжить.
p53 метилированный по лизину, чтобы регулировать его активацию и взаимодействие с другими белками в ответе на повреждение ДНК. Это пример регуляции белок-белковых взаимодействий и активации белков. p53 является известным подавитель опухолей что активирует Ремонт ДНК пути, инициирует апоптоз, и приостанавливает клеточный цикл. В целом, он реагирует на мутации в ДНК, передавая сигнал клетке, чтобы исправить их или инициировать гибель клетки, так что эти мутации не могут способствовать развитию рака.
NF-κB (белок, участвующий в воспалении) является известной мишенью метилирования метилтрансферазы. SETD6, который выключает передачу сигналов NF-κB путем ингибирования одной из его субъединиц, RelA. Это снижает активацию транскрипции и воспалительная реакция, превращая метилирование NF-κB в регуляторный процесс, с помощью которого снижается передача клеточных сигналов через этот путь.[3]
Метилтрансферазы природного продукта обеспечивают различные входы в метаболические пути, включая наличие кофакторов, сигнальных молекул и метаболитов. Это регулирует различные клеточные пути, контролируя активность белка.
Типы
Гистоновые метилтрансферазы
Гистоновые метилтрансферазы имеют решающее значение для генетической регуляции в эпигенетический уровень. Они модифицируют в основном лизин на ε-азоте и аргинин гуанидиний группа на гистоновых хвостах. Лизинметилтрансферазы и аргининметилтрансферазы представляют собой уникальные классы ферментов, но обе связывают SAM в качестве донора метила для своих гистон субстраты. Аминокислоты лизина могут быть модифицированы одной, двумя или тремя метильными группами, а аминокислоты аргинина могут быть модифицированы одной или двумя метильными группами. Это увеличивает силу положительного заряда и остатка. гидрофобность, позволяя другим белкам распознавать метильные метки. Эффект этой модификации зависит от расположения модификации на гистоновом хвосте и других модификациях гистонов вокруг него. Расположение модификаций может частично определяться последовательностью ДНК, а также небольшими некодирующие РНК и метилирование самой ДНК. Чаще всего у позвоночных метилируется гистон H3 или H4. Может произойти либо повышенная, либо пониженная транскрипция генов, связанных с модификацией. Повысился транскрипция является результатом уменьшения хроматин конденсация, в то время как снижение транскрипции является результатом повышенной конденсации хроматина.[4] Метильные метки на гистонах вносят свой вклад в эти изменения, служа сайтами для набора других белков, которые могут в дальнейшем модифицировать хроматин.[5]
N-концевые метилтрансферазы
N-альфа-метилтрансферазы переносят метильную группу от SAM к N-концевой азот на белковых мишенях. N-терминал метионин сначала расщепляется другим ферментом, и X-Пролин -Консенсусная последовательность лизина распознается метилтрансферазой. Для всех известных субстратов аминокислота X Аланин, Серин, или Пролин. Эта реакция дает метилированный белок и SAH. Известные мишени этих метилтрансфераз у человека включают ПКР-1 (регулятор ядерных транспортных белков) и Белок ретинобластомы (белок-супрессор опухолей, который препятствует чрезмерному делению клеток). Метилирование RCC-1 особенно важно в митоз поскольку он координирует локализацию некоторых ядерный белки в отсутствие ядерная оболочка. Когда ПКР-1 не метилирован, деление клеток происходит ненормально после образования дополнительных полюса шпинделя.[6] Функция N-концевого метилирования белка ретинобластомы неизвестна.
ДНК / РНК метилтрансферазы

Метилирование ДНК, ключевой компонент генетической регуляции, происходит в основном у 5 атомов углерода основания. цитозин, формируя 5’метилцитозин (см. слева).[7] Метилирование - это эпигенетический модификация, катализируемая Ферменты ДНК-метилтрансферазы, включая DNMT1, DNMT2 и DNMT3. Эти ферменты используют S-аденозилметионин в качестве донора метила и содержат несколько высококонсервативных структурных особенностей между тремя формами; они включают сайт связывания S-аденозилметионина, вицинальную пару пролин-цистеин, которая образует тиолат-анион, важный для механизма реакции, и карман связывания цитозинового субстрата. Многие особенности ДНК-метилтрансфераз очень высоки. консервированный во многих сферах жизни, от бактерий до млекопитающих. Помимо управления экспрессия определенных генов, существует множество белковых комплексов, многие из которых имеют значение для здоровья человека, которые связываются только с сайты узнавания метилированной ДНК. Многие из ранних ДНК-метилтрансфераз считались производными от метилтрансфераз РНК, которые, как предполагалось, были активны в Мир РНК для защиты многих видов примитивной РНК.[8]Метилирование РНК наблюдалось в различных типах видов РНК, а именно.мРНК, рРНК, тРНК, snoRNA, мяРНК, miRNA, тмРНК а также вирусные виды РНК. Специфические РНК-метилтрансферазы используются клетками для маркировки их на видах РНК в соответствии с потребностями и окружающей средой, преобладающей вокруг клеток, которые образуют часть поля, называемого молекулярным. эпигенетика. 2'-O-метилирование, m6A Метилирование, метилирование m1G, а также m5C - наиболее часто наблюдаемые метки метилирования в различных типах РНК.
6A - фермент, катализирующий следующие химические реакции:[9]
S-аденозил-L-метионин + ДНК аденин S-аденозил-L-гомоцистеин + ДНК 6-метиламинопурин
m6A в основном был обнаружен у прокариот до 2015 года, когда он был также идентифицирован у некоторых эукариот. Метилтрансферазы m6A метилируют аминогруппу в ДНК в положении C-6 специально для предотвращения переваривания системой хозяина собственного генома с помощью рестрикционных ферментов.[10]
m5C играет роль в регуляции транскрипции генов. Трансферазы m5C - это ферменты, которые продуцируют C5-метилцитозин в ДНК в положении C-5 цитозина и обнаруживаются у большинства растений и некоторых эукариот.[11]
Метилтрансферазы природных продуктов
Метилтрансферазы природных продуктов (NPMT) представляют собой разнообразную группу ферментов, которые добавляют метильные группы к естественным малым молекулам. Как и многие метилтрансферазы, SAM используется в качестве донора метила и продуцируется SAH. Метильные группы добавляются к атомам S, N, O или C и классифицируются по тому, какой из этих атомов модифицирован, при этом O-метилтрансферазы представляют самый большой класс. Метилированные продукты этих реакций выполняют множество функций, включая кофакторы, пигменты, сигнальные соединения и метаболиты. NPMT могут выполнять регулирующую роль, изменяя реактивность и доступность этих соединений. Эти ферменты не являются высококонсервативными у разных видов, так как они выполняют более специфическую функцию, обеспечивая небольшие молекулы для специализированных путей у видов или меньших групп видов. Это разнообразие отражается в разнообразии каталитических стратегий, включая общие кислотно-щелочной катализ, катализ на основе металлов, и близость и эффекты десольватации не требует каталитических аминокислот. NPMT представляют собой наиболее функционально разнообразный класс метилтрансфераз.[12]

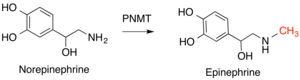
Важные примеры этого класса ферментов у людей включают: фенилэтаноламин N-метилтрансфераза (PNMT), который преобразует норэпинефрин к адреналин,[14] и гистамин N-метилтрансфераза (HNMT), который метилирует гистамин в процессе метаболизма гистамина.[15] КатехолО-метилтрансфераза (COMT) разлагает класс молекул, известный как кексоламины это включает дофамин, адреналин и норэпенеферин.[16]
Независимые от SAM метилтрансферазы
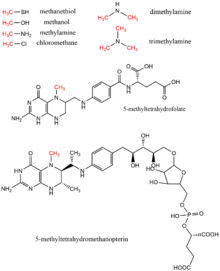
Метанол, метилтетрагидрофолат, мононуклеоз-, ди-, и триметиламин, метантиол, метилтетрагидрометаноптерин, и хлорметан все метильные доноры, обнаруженные в биологии как доноры метильных групп, обычно в ферментативных реакциях с использованием кофактор витамин B12.[17] Эти субстраты способствуют путям переноса метила, включая биосинтез метионина, метаногенез, и ацетогенез.
Радикальные метилтрансферазы SAM
Основываясь на различных белковых структурах и механизмах катализа, существует 3 различных типа радикальных метилаз SAM (RS): класса A, B и C. RS метилазы класса A лучше всего охарактеризованы из 4 ферментов и связаны как с RlmN, так и с Cfr. RlmN встречается повсеместно в бактериях, что повышает точность трансляции, а RlmN катализирует метилирование C2 аденозина 2503 (A2503) в 23 S рРНК и C2 аденозина (A37). Cfr, с другой стороны, катализирует метилирование C8 из A2503, а также катализирует метилирование C2.[18] Класс B в настоящее время является самым большим классом радикальных метилаз SAM, которые могут матилировать оба зр2-гибридные и зр3-гибридизированные атомы углерода в разных наборах субстратов в отличие от класса A, который только катализирует зр2-гибридизированные атомы углерода. Основное отличие, которое отличает класс B от других, - это дополнительный N-концевой кобаламин-связывающий домен, который связывается с доменом RS.[19] Метилаза класса C имеет гомологичную последовательность с ферментом RS, копропорфириноген III оксидазой (HemN), который также катализирует метилирование зр2-гибридизированных углеродных центра, но в нем отсутствуют 2 цистеина, необходимые для метилирования в механизме класса A.[18]
Клиническое значение
Как и в случае любого биологического процесса, который регулирует экспрессию и / или функцию генов, аномальное метилирование ДНК связано с генетическими нарушениями, такими как ICF, Синдром Ретта, и Синдром ломкой Х-хромосомы.[2] Раковые клетки обычно проявляют меньшую активность метилирования ДНК в целом, хотя часто гиперметилирование участков, которые не метилированы в нормальных клетках; это чрезмерное метилирование часто действует как способ инактивировать гены-супрессоры опухолей. Ингибирование общей активности ДНК-метилтрансферазы было предложено в качестве варианта лечения, но ингибиторы DNMT, аналоги их цитозин субстраты оказались очень токсичными из-за их сходства с цитозином (см. справа); это сходство с нуклеотидом вызывает включение ингибитора в Трансляция ДНК, вызывая синтез нефункционирующей ДНК.
Метилаза, которая изменяет сайт связывания рибосомной РНК антибиотика. линезолид вызывает перекрестную резистентность к другим антибиотикам, действующим на рибосомную РНК. Плазмида векторы, способные передавать этот ген, являются причиной потенциально опасной перекрестной устойчивости.[20]
Примеры ферментов метилтрансферазы, влияющих на заболевание:
- тиопуринметилтрансфераза: дефекты этого гена вызывают токсическое накопление тиопуриновых соединений, препаратов, используемых в химиотерапии и терапии иммунодепрессантами
- метионинсинтаза: злокачественная анемия, вызванный Витамин B12 дефицит, вызванный отсутствием кофактор для фермента метионинсинтазы
Приложения в открытии и разработке лекарств
Недавняя работа показала, что метилтрансферазы, участвующие в метилировании естественных противораковых агентов, для использования S-аденозил метионин (SAM) аналоги, содержащие альтернативные алкильные группы вместо метила. Разработка простой химико-ферментативной платформы для создания и использования дифференциально алкилированных аналогов SAM в контексте открытие лекарств и разработка лекарств известен как алкилрандомизация.[21]
Применения в лечении рака
В клетках человека было обнаружено, что m5C был связан с аномальными опухолевыми клетками при раке.[22] Роль и потенциальное применение m5C включает балансирование поврежденной ДНК при раке как гиперметилировании, так и гипометилировании. Эпигенетическая репарация ДНК может быть применена путем изменения количества m5C в обоих типах раковых клеток (гиперметилирование / гипометилирование), а также в окружающей среде рака, чтобы достичь эквивалентной точки для подавления опухолевых клеток.[23]
Примеры
Примеры включают:
- Катехол-О-метилтрансфераза
- ДНК-метилтрансфераза
- Гистоновая метилтрансфераза
- 5-метилтетрагидрофолат-гомоцистеин метилтрансфераза
- О-метилтрансфераза
- метионинсинтаза
- белок корриноид-железо-сера
Рекомендации
- ^ Katz, J. E .; Длакич, М; Кларк С. (18 июля 2003 г.). «Автоматическая идентификация предполагаемых метилтрансфераз из открытых рамок считывания генома». Молекулярная и клеточная протеомика. 2 (8): 525–40. Дои:10.1074 / mcp.M300037-MCP200. PMID 12872006.
- ^ а б Siedlecki, P; Зеленкевич, П (2006). «Метилтрансферазы ДНК млекопитающих». Acta Biochimica Polonica. 53 (2): 245–56. Дои:10.18388 / abp.2006_3337. PMID 16582985.
- ^ Леви, Дэн; и другие. (5 декабря 2010 г.). «Метилирование лизина субъединицы NF-κB RelA с помощью SETD6 связывает активность гистон-метилтрансферазы GLP на хроматине с тонической репрессией передачи сигналов NF-κB». Иммунология природы. 12 (1): 29–36. Дои:10.1038 / н.в.1968. ЧВК 3074206. PMID 21131967.
- ^ Тернер, Брайан М. (2001). Хроматин и регуляция генов: механизмы в эпигенетике. Молден, Массачусетс: Blackwell Science. ISBN 978-0865427433.
- ^ Грир, Эрик Л .; Ши, Ян (3 апреля 2012 г.). «Метилирование гистонов: динамический показатель здоровья, болезней и наследственности». Природа Обзоры Генетика. 13 (5): 343–357. Дои:10.1038 / nrg3173. ЧВК 4073795. PMID 22473383.
- ^ Кларк, Пол (май 2007 г.). «Закрепление ПКР1 за хвост». Природа клеточной биологии. 9 (5): 485–487. Дои:10.1038 / ncb0507-485. PMID 17473856.
- ^ Лан, Дж; Хуа, S; Он, Х; Чжан, Y (2010). «ДНК-метилтрансферазы и метилсвязывающие белки млекопитающих». Acta Biochimica et Biophysica Sinica. 42 (4): 243–52. Дои:10.1093 / abbs / gmq015. PMID 20383462.
- ^ Рана, Аджай К .; Анкри, Серж (01.01.2016). «Возрождение мира РНК: взгляд на появление метилтрансфераз РНК». Границы генетики. 7: 99. Дои:10.3389 / fgene.2016.00099. ЧВК 4893491. PMID 27375676.
- ^ Кесслер, Кристоф; Манта, Висентиу (01.01.1990). «Специфичность рестрикционных эндонуклеаз и метилтрансфераз модификации ДНК - обзор (издание 3)». Ген. 92 (1): 1–240. Дои:10.1016 / 0378-1119 (90) 90486-Б. ISSN 0378-1119. PMID 2172084.
- ^ Нарва, Кеннет Э .; Ван Эттен, Джеймс Л .; Слатко, Бартон Э .; Беннер, Джек С. (1988-12-25). «Аминокислотная последовательность эукариотической ДНК [N6-аденин] метилтрансферазы M · CviBIII, имеет области сходства с прокариотическим изошизомером M · TaqI и другими ДНК [N6-аденин] метилтрансферазами». Ген. 74 (1): 253–259. Дои:10.1016/0378-1119(88)90298-3. ISSN 0378-1119. PMID 3248728.
- ^ Посфаи, Янош; Bhagwat, Ashok S .; Робертс, Ричард Дж. (1988-12-25). «Мотивы последовательностей, специфичные для цитозинметилтрансфераз». Ген. 74 (1): 261–265. Дои:10.1016/0378-1119(88)90299-5. ISSN 0378-1119. PMID 3248729.
- ^ Лискомб, Дэвид К .; Луи, Гордон В .; Ноэль, Джозеф П. (2012). «Архитектура, механизмы и молекулярная эволюция метилтрансфераз природного продукта». Отчеты о натуральных продуктах. 29 (10): 1238–50. Дои:10.1039 / c2np20029e. PMID 22850796.
- ^ Ашихара, Хироши; Ёкота, Такао; Крозье, Алан (2013). Биосинтез и катаболизм пуриновых алкалоидов. Успехи ботанических исследований. 68. С. 111–138. Дои:10.1016 / B978-0-12-408061-4.00004-3. ISBN 9780124080614.
- ^ «PNMT фенилэтаноламин N-метилтрансфераза». Регистр генетического тестирования NCBI. Получено 18 февраля 2014.
- ^ «HNMT гистамин N-метилтрансфераза». Регистр генетического тестирования NCBI. Получено 18 февраля 2014.
- ^ «Катехол-О-метилтрансфераза COMT». Регистр генетического тестирования NCBI. Получено 18 февраля 2014.
- ^ Рэгсдейл, С. «Катализ переноса метильных групп с участием тетрагидрофолата и B12» Витамины и гормоны, 2008.
- ^ а б Bauerle, Matthew R .; Schwalm, Erica L .; Букер, Сквайр Дж. (13 февраля 2015 г.). «Механистическое разнообразие радикального S-аденозилметионина (SAM) -зависимого метилирования». Журнал биологической химии. 290 (7): 3995–4002. Дои:10.1074 / jbc.R114.607044. ISSN 0021-9258. ЧВК 4326810. PMID 25477520.
- ^ София, Х. Дж .; Chen, G .; Hetzler, B.G .; Reyes-Spindola, J. F .; Миллер, Н. Э. (2001-03-01). «Радикальный SAM, новое суперсемейство белков, связывающее неразрешенные этапы знакомых биосинтетических путей с радикальными механизмами: функциональная характеристика с использованием новых методов анализа и визуализации информации». Исследования нуклеиновых кислот. 29 (5): 1097–1106. Дои:10.1093 / nar / 29.5.1097. ISSN 1362-4962. ЧВК 29726. PMID 11222759.
- ^ Моралес Дж., Пиказо Дж. Дж., Баос Е., Кандель Ф. Дж., Арриби А., Пелаэс Б., Андраде Р., де ла Торре М. А., Феререс Дж., Санчес-Гарсия М. (март 2010 г.). «Устойчивость к линезолиду опосредуется геном cfr в первом отчете о вспышке устойчивого к линезолиду Staphylococcus aureus». Clin. Заразить. Dis. 50 (6): 821–5. Дои:10.1086/650574. PMID 20144045.
- ^ Сингх, S; Чжан, Дж; Хубер, Т.Д .; Сункара, М. Херли, К; Гофф, РД; Ванга, G; Чжан, Вт; Лю, К; Рор, Дж; Ван Ланен, С. Г.; Моррис, AJ; Thorson, JS (7 апреля 2014 г.). «Простые хемоэнзиматические стратегии синтеза и использования аналогов S-аденозил- (L) -метионина». Angewandte Chemie International Edition на английском языке. 53 (15): 3965–9. Дои:10.1002 / anie.201308272. ЧВК 4076696. PMID 24616228.
- ^ Джонс, Питер А. (1996-06-01). «Ошибки метилирования ДНК и рак». Исследования рака. 56 (11): 2463–2467. ISSN 0008-5472. PMID 8653676.
- ^ D, Ханахан; Ра, Вайнберг (2011-03-04). «Признаки рака: следующее поколение». Клетка. 144 (5): 646–74. Дои:10.1016 / j.cell.2011.02.013. PMID 21376230.
дальнейшее чтение
- Метилтрансферазы в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)
- Трехмерная структура ДНК-метилтрансферазы
- Новая метилтрансфераза: 7SK мяРНК, блокирующий метилфосфатный фермент как видно на Flintbox
- «Роль метилирования в экспрессии генов» о природе Scitable
- «Питание и депрессия: питание, метилирование и депрессия» по психологии сегодня
- "Метилирование ДНК - что такое метилирование ДНК?" из News-Medical.net
- «Метилирование гистонового лизина» Генетические пути с участием гистоновых метилтрансфераз из технологии передачи сигналов клеток