G-квадруплекс - G-quadruplex
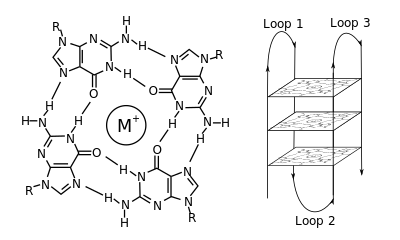
В молекулярной биологии G-квадруплекс второстепенные конструкции (G4) образуются в нуклеиновых кислотах последовательностями, богатыми гуанин.[2] Они имеют спиралевидную форму и содержат тетрады гуанина, которые могут образовываться из одного,[3] два[4] или четыре пряди.[5] Мономолекулярные формы часто встречаются в природе вблизи концов хромосом, более известных как теломерные области, а также в регуляторных областях транскрипции множества генов, как у микробов.[6][7] и среди позвоночных [8][7] в том числе онкогены у человека.[9] Четыре гуаниновых основания могут связываться через Hoogsteen водородная связь с образованием плоской квадратной структуры, называемой гуаниновая тетрада (G-тетрада или G-квартет), и две или более тетрады гуанина (из G-трактов, непрерывных прогонов гуанина) могут накладываться друг на друга, образуя G-квадруплекс.
Размещение и соединение для образования G-квадруплексов не случайно и служит очень необычным функциональным целям. Квадруплексная структура дополнительно стабилизируется наличием катион, особенно калий, который находится в центральном канале между каждой парой тетрад.[3] Они могут состоять из ДНК, РНК, LNA, и PNA, и возможно внутримолекулярный, бимолекулярный, или тетрамолекулярный.[10] В зависимости от направления нитей или частей нити, образующих тетрады, структуры можно описать как параллельно или же антипараллельный. Структуры G-квадруплексов могут быть предсказаны с помощью вычислений на основе мотивов последовательностей ДНК или РНК,[11][12] но их фактические структуры могут сильно различаться внутри и между мотивами, которых может быть более 100000 на геном. Их деятельность в основных генетических процессах является активной областью исследований в области теломер, регуляции генов и функциональной геномики.[13][14]
История
Выявление конструкций с высоким гуанин ассоциация стала очевидной в начале 1960-х годов, когда были идентифицированы гелеобразные вещества, связанные с гуанинами.[15] В частности, это исследование подробно описало четырехцепочечный ДНК структуры с высокой ассоциацией гуанинов, которые позже были идентифицированы в эукариотических теломерный области ДНК в 1980-х гг.[16] Важность открытия структуры G-квадруплекса была описана следующим образом: «Если G-квадруплексы образуются так легко, in vitro, Природа нашла способ их использовать in vivo” - Аарон Клуг, Лауреат Нобелевской премии по химии (1982). Интерес к in vivo Функция G-квадруплексов резко возросла после того, как крупномасштабный полногеномный анализ показал преобладание потенциальных последовательностей, образующих G-квадруплекс (pG4), в промоторах генов человека, шимпанзе, мыши и крысы, что было представлено на Первой международной встрече G-квадруплексов в апреле 2007 года в Луисвилле, Кентукки.[7] В 2006 году сообщалось о преобладании G-квадруплексов в промоторах генов нескольких бактериальных геномов, которые предсказывают опосредованную G-квадруплексами регуляцию генов.[6] С обилием G-квадруплексов in vivoэти структуры играют биологически значимую роль благодаря взаимодействиям с промоторными областями онкогенов и теломерными областями цепей ДНК. Текущие исследования состоят в определении биологической функции этих структур G-квадруплексов в отношении конкретных онкогенов и обнаружении эффективных терапевтических методов лечения рака, основанных на взаимодействии с G-квадруплексами.
Топология
Длина последовательностей нуклеиновых кислот, участвующих в образовании тетрад, определяет, как складывается квадруплекс. Короткие последовательности, состоящие только из одной непрерывной серии из трех или более оснований гуанина, требуют четырех отдельных цепей для образования квадруплекса. Такой квадруплекс описывается как тетрамолекулярный, что отражает потребность в четырех отдельных цепях. Термин ДНК G4 был первоначально зарезервирован для этих тетрамолекулярных структур, которые могут играть роль в мейоз.[5] Однако, как в настоящее время используется в молекулярной биологии, термин G4 может обозначать G-квадруплексы любой молекулярности. Более длинные последовательности, которые содержат два непрерывных ряда из трех или более оснований гуанина, где участки гуанина разделены одним или несколькими основаниями, требуют только двух таких последовательностей, чтобы обеспечить достаточное количество оснований гуанина для образования квадруплекса. Эти структуры, образованные из двух отдельных G-богатых цепей, называются бимолекулярными квадруплексами. Наконец, последовательности, которые содержат четыре различных ряда оснований гуанина, могут сами по себе образовывать стабильные квадруплексные структуры, и квадруплекс, полностью сформированный из одной цепи, называется внутримолекулярным квадруплексом.[17]
В зависимости от того, как отдельные ряды оснований гуанина расположены в бимолекулярном или внутримолекулярном квадруплексе, квадруплекс может принимать одну из нескольких топологий с различными конфигурациями петель.[18] Если все цепи ДНК движутся в одном направлении, квадруплекс называется параллельным. Для внутримолекулярных квадруплексов это означает, что любые присутствующие области петель должны быть пропеллерного типа и располагаться по бокам квадруплекса. Если один или несколько рядов оснований гуанина имеют направление 5’-3 ’, противоположное другим рядам оснований гуанина, считается, что квадруплекс принял антипараллельную топологию. Петли, соединяющие ряды оснований гуанина во внутримолекулярных антипараллельных квадруплексах, являются либо диагональными, соединяющими два диагонально противоположных ряда оснований гуанина, либо петлями латерального (на ребро) типа, соединяющими два соседних ряда пар оснований гуанина.
В квадруплексах, образованных из двухцепочечной ДНК, также обсуждались возможные межцепочечные топологии.[19].[20]Межцепочечные квадруплексы содержат гуанины, происходящие из обеих цепей дцДНК.
Структура и функциональная роль в геноме
После секвенирования человеческого геном было обнаружено множество богатых гуанином последовательностей, которые могли образовывать квадруплексы.[21] В зависимости от типа клетки и клеточного цикла, опосредующие факторы, такие как ДНК-связывающие белки, на хроматин, состоящий из ДНК, плотно намотанной на гистон белки, а также другие условия окружающей среды и стрессы влияют на динамическое образование квадруплексов. Например, количественные оценки термодинамика из молекулярное скопление указывают на то, что антипараллельный g-квадруплекс стабилизируется молекулярным краудингом.[22] Этот эффект, по-видимому, опосредован изменением гидратации ДНК и его влиянием на Базовая пара Hoogsteen склеивание.[23] Эти квадруплексы, по-видимому, легко возникали на концах хромосома. Кроме того, склонность к образованию g-квадруплексов во время транскрипции в последовательностях РНК с потенциалом образования взаимоисключающих заколка для волос или G-квадруплексные структуры сильно зависят от положения образующей шпильку последовательности.[24]
Поскольку ферменты репарации естественным образом распознают концы линейных хромосом как поврежденную ДНК и обрабатывают их как таковые, оказывая вредное воздействие на клетку, необходимы четкая передача сигналов и жесткая регуляция на концах линейных хромосом. Теломеры функция для обеспечения этой сигнализации. Теломеры, богатые гуанином и склонные к образованию g-квадруплексов, расположены на концевых концах хромосом и помогают поддерживать целостность генома, защищая эти уязвимые концевые концы от нестабильности.
Эти теломерные области характеризуются длинными участками двухцепочечных повторов CCCTAA: TTAGGG. Повторы заканчиваются 3’-выступом из 10-50 однонитевых повторов TTAGGG. Гетеродимерный сложный рибонуклеопротеиновый фермент теломераза добавляет повторы TTAGGG на 3’-конце цепей ДНК. На этих 3’-концевых выступах богатый G выступ может образовывать вторичные структуры, такие как G-квадруплексы, если выступ длиннее четырех повторов TTAGGG. Наличие этих структур предотвращает удлинение теломер за счет теломеразного комплекса.[25]
Теломерные квадруплексы
Теломерный было показано, что повторы в различных организмах образуют эти квадруплексные структуры. in vitro, и впоследствии было показано, что они образуют in vivo.[26][27] Человеческий теломерный повтор (который одинаков для всех позвоночные ) состоит из множества повторов секвенированного (GGTTAG), и квадруплексы, образованные этой структурой, могут иметь форму бусинок размером от 5 до 8 нм и были хорошо изучены ЯМР, ТЕА и рентгеновский снимок Кристальная структура решимость.[28] Было показано, что образование этих квадруплексов в теломерах снижает активность фермент теломераза, который отвечает за поддержание длины теломер и участвует примерно в 85% всех раки. Это активная цель открытия новых лекарств, в том числе теломестатин.
Нетеломерные квадруплексы
Квадруплексы присутствуют в других местах, кроме теломер. Анализ геномов человека, шимпанзе, мыши и крысы показал огромное количество потенциально образующих G-квадруплекс (pG4) последовательностей в нетеломерных областях. Большое количество нетеломерных G-квадруплексов было обнаружено в промоторах генов и сохранялось у всех видов.[6][7] Точно так же большое количество G-квадруплексов было обнаружено в E. coli и сотнях других микробных геномов. Здесь также, как и у позвоночных, G-квадруплексы были обогащены промоторами генов.[6] Хотя эти исследования предсказывают опосредованную G-quadruplex регуляцию генов, маловероятно, что все pG4s будут формироваться in vivo. В протоонкоген c-myc образует квадруплекс в нуклеаза гиперчувствительная область, критическая для активности гена.[29][30] Показано, что другие гены образуют G-квадруплексы в своих промоутер регионы включают курица β-глобин ген, человек убиквитин -лигаза RFP2 и протоонкогены c-kit, bcl-2, VEGF, H-ras и N-ras.[31][32][33]
Геном были проведены широкие исследования на основе правила складывания квадруплексов, которые выявили 376000 предполагаемых квадруплексных последовательностей (PQS) в человек геном, хотя не все из них, вероятно, образуют in vivo.[34] Подобные исследования выявили предполагаемые G-квадруплексы в прокариоты.[35] Существует несколько возможных моделей того, как квадруплексы могут влиять на активность генов: усиление регулирования или же подавление. Одна модель показана ниже, с образованием G-квадруплекса в или около промоутер блокировка транскрипция гена и, следовательно, деактивируя его. В другой модели квадруплекс, образованный на некодирующей цепи ДНК, помогает поддерживать открытую конформацию кодирующей цепи ДНК и усиливать экспрессию соответствующего гена.
Функция
Было высказано предположение, что образование квадруплексов играет роль в тяжелая цепь иммуноглобулина переключение.[5] По мере развития клеток развивались механизмы разрешения (т. Е. Раскручивания) образующихся квадруплексов. Образование квадруплексов может быть потенциально опасным для клетки; геликасы WRN и Белок синдрома Блума обладают высоким сродством к разрешению G-квадруплексов ДНК.[36] Геликаза DEAH / RHA, DHX36, также был идентифицирован как ключевая резольваза G-квадруплекса.[37][38] В 2009 году было обнаружено, что белок-супрессор метастазов NM23H2 (также известный как NME2) напрямую взаимодействует с G-квадруплексом в промоторе гена c-myc и транскрипционно регулирует c-myc.[39][40] Совсем недавно было сообщено, что NM23H2 взаимодействует с G-квадруплексом в промоторе гена теломеразы человека (hTERT) и регулирует экспрессию hTERT. [41] В 2019 году было показано, что теломер-связывающий фактор-2 (TRF2 или TERF2) связывается с тысячами нетеломерных G-квадруплексов в геноме человека посредством TRF2 ChIP-seq.[42] Существует множество исследований, в которых квадруплексы участвуют как в позитивной, так и в негативной регуляции транскрипции, включая эпигенетическую регуляцию таких генов, как hTERT.[41] Сообщалось также, что функция G-квадруплексов позволяет запрограммировать рекомбинацию тяжелых генов иммунологлобина и систему антигенных вариаций пилина патогенного микроорганизма. Neisseria.[43] Роль квадруплексной структуры в управлении трансляцией изучена недостаточно. Прямая визуализация структур G-квадруплексов в клетках человека[44] а также сокристаллическая структура РНК-геликазы, связанной с G-квадруплексом[45] предоставили важные подтверждения их актуальности для клеточной биологии. Возможные положительные и отрицательные роли квадруплексов в репликации и функции теломер остаются спорными. Т-петли и G-квадруплексы описываются как две третичные структуры ДНК, которые защищают концы теломер и регулируют длину теломер.[46]
Рак
Теломеры
Последовательности, образующие G-квадруплекс, преобладают в эукариотических клетках, особенно в теломерах, 5` нетранслируемых цепях и горячих точках транслокации. G-квадруплексы могут подавлять нормальную функцию клеток, и в здоровых клетках они легко и легко разворачиваются с помощью геликаза. Однако в раковых клетках, которые имеют мутировавшую геликазу, эти комплексы не могут быть размотаны и приводят к потенциальному повреждению клетки. Это вызывает репликацию поврежденных и раковых клеток. Для достижения терапевтических успехов стабилизация G-квадруплексов раковых клеток может подавлять рост и репликацию клеток, что приводит к смерть клетки.[47]
Регионы промоутеров
Наряду с ассоциацией G-квадруплексов в теломерный области ДНК, G-квадруплексные структуры были идентифицированы в различных прото-онкоген промоутерские регионы. Структуры, наиболее присутствующие в промоторных областях этих онкогенов, как правило, представляют собой структуры G-квадруплекса с параллельными цепями ДНК.[48] Некоторые из этих онкогенов включают c-KIT, PDGF-A, c-Myc и VEGF, показывающий важность этой вторичной структуры в росте и развитии рака. Хотя формирование структуры G-квадруплекса до некоторой степени варьируется для разных промоторных областей онкогенов, последовательная стабилизация этих структур была обнаружена при развитии рака.[49] Текущие терапевтические исследования активно фокусируются на стабилизации структур G-квадруплекса для остановки нерегулируемого роста и деления клеток.
Один конкретный участок гена, путь c-myc, играет важную роль в регуляции белкового продукта c-Myc. С этим продуктом белок c-Myc действует в процессах апоптоза и роста или развития клеток, а также в качестве контроля транскрипции у человека. теломераза обратная транскриптаза.[50] В 2009 г. было показано, что взаимодействие G-квадруплекса промотора c-Myc с NM23H2 регулирует c-Myc в раковых клетках. [39]
Регуляция c-myc через обратную транскриптазу теломеразы человека (hTERT) также напрямую регулируется через промотор G-квадруплекс путем взаимодействия с фактором транскрипции NM23H2, где эпигенетические модификации зависят от ассоциации NM23H2-G-квадруплекса.[41] Недавно сообщалось, что эпигенетическая регуляция hTERT опосредуется посредством взаимодействия G-квадруплекса промотора hTERT с теломерным фактором TRF2.[51]
Другой путь гена связан с геном VEGF, фактором роста эндотелия сосудов, который остается вовлеченным в процесс ангиогенеза или образования новых кровеносных сосудов. Формирование внутримолекулярной структуры G-квадруплекса было показано в исследованиях полипуринового тракта промоутер регион гена VEGF. Благодаря недавним исследованиям роли функции G-квадруплекса in vivo было показано, что стабилизация структур G-квадруплекса регулирует транскрипцию гена VEGF с ингибированием факторов транскрипции в этом пути. Внутримолекулярные структуры G-квадруплексов образуются в основном за счет обильной гуаниновой последовательности в промоторной области этого специфического пути.[52] Ген CDKN1A (также известный как p21), ген ингибитора-1 киназы контрольной точки клеточного цикла, содержит промоторный G-квадруплекс. Взаимодействие этого G-квадруплекса с TRF2 (также известным как TERF2) привело к эпигенетической регуляции p21, что было протестировано с использованием лиганда, связывающего G-квадруплекс 360A.[53]
Фактор 1ɑ, индуцируемый гипоксией, HIF-1ɑ, остается вовлеченным в передачу сигналов рака через его связывание с элементом ответа на гипоксию, HRE, в присутствии гипоксии, чтобы запустить процесс ангиогенез. Благодаря недавним исследованиям этого специфического генного пути участок полипурина и полипиримидина позволяет транскрипцию этого специфического гена и формирование внутримолекулярной структуры G-квадруплекса. Однако необходимы дополнительные исследования, чтобы определить, регулирует ли образование G-квадруплекса экспрессию этого гена положительным или отрицательным образом.[54]
Онкоген c-kit имеет дело с путем, кодирующим RTK, который, как было показано, имеет повышенные уровни экспрессии при определенных типах рака. Богатая гуаниновая последовательность этой промоторной области показала способность образовывать множество квадруплексов. Текущие исследования этого пути сосредоточены на обнаружении биологической функции этого специфического образования квадруплекса на пути c-kit, в то время как эта квадруплексная последовательность была замечена у различных видов.[33]
Онкоген RET участвует в транскрипции киназа который часто встречается при некоторых типах рака. Богатая гуанином последовательность в промоторной области этого пути обусловливает необходимость базовой транскрипции этой рецепторной тирозинкиназы. При определенных типах рака уровень экспрессии белка RET повышен. Исследование этого пути предполагает образование G-квадруплекса в промоторной области и подходящую мишень для терапевтического лечения.[55]
Другой путь онкогена с участием PDGF-A, фактора роста тромбоцитов, включает процесс заживления ран и функционирует как митогенные факторы роста клеток. Высокие уровни экспрессии PDGF были связаны с повышенным ростом клеток и раком. Присутствие богатой гуанином последовательности в промоторной области PDGF-A продемонстрировало способность образовывать внутримолекулярные параллельные G-квадруплексные структуры и, как предполагается, играет роль в регуляции транскрипции PDGF-A. Однако исследования также выявили присутствие структур G-квадруплекса в этой области из-за взаимодействия TMPyP4 с этой последовательностью промотора.[56]
Терапия
Теломеры обычно состоят из G-квадруплексов и остаются важными объектами терапевтических исследований и открытий. Эти комплексы обладают высоким сродством к порфириновые кольца что делает их эффективными противораковыми средствами. Однако использование TMPyP4 ограничено из-за его неизбирательности по отношению к теломерам раковых клеток и нормальной двухцепочечной ДНК (дцДНК). Для решения этой проблемы был синтезирован аналог TMPyP4, известный как 5Me, который нацелен только на G-квадруплексную ДНК, которая ингибирует рост рака более эффективно, чем TMPyP4.[57]
Дизайн и разработка лигандов остается важной областью исследований терапевтических реагентов из-за обилия G-квадруплексов и их многочисленных конформационных различий. Один тип лиганда, включающий производное хиндолина, SYUIQ-05, использует стабилизацию G-квадруплексов в промоторных областях для ингибирования продукции как белкового продукта c-Myc, так и обратной транскриптазы теломеразы человека (hTERT). Этот основной путь нацеливания на эту область приводит к отсутствию элонгации теломеразы, что приводит к остановке развития клеток. По-прежнему необходимы дальнейшие исследования для открытия единственного гена-мишени, чтобы минимизировать нежелательную реактивность с более эффективной противоопухолевой активностью.[50]
Лиганды, связывающие квадруплексы
Одним из способов индуцирования или стабилизации образования G-квадруплекса является введение молекулы, которая может связываться со структурой G-квадруплекса. Номер лиганды, которые могут быть как небольшими молекулами, так и белки, может связываться с G-квадруплексом. Эти лиганды могут быть встречающимися в природе или синтетическими. Это становится все более обширной областью исследований в области генетики, биохимии и фармакологии.
Катионный порфирины было показано интеркалятивно связываться с G-квадруплексами, а также с молекулой теломестатин.
Связывание лигандов с G-квадруплексами жизненно важно для борьбы с раком, потому что G-квадруплексы обычно обнаруживаются в горячих точках транслокаций. MM41, лиганд, который селективно связывается с квадруплексом на BCL-2 промотор имеет форму с центральным ядром и 4 боковыми цепями, пространственно разветвляющимися. Форма лиганда жизненно важна, потому что она точно соответствует квадруплексу, у которого сложены квартеты и петли нуклеиновые кислоты удерживая его вместе. При связывании центральный хромофор MM41 располагается наверху 3’-концевого G-квартета, а боковые цепи лиганда связываются с петлями квадруплекса. Квартет и хромофор связаны π-π связь в то время как боковые цепи и петли не связаны, а находятся в непосредственной близости. Что делает это связывание прочным, так это плавность положения петель для лучшего связывания с боковыми цепями лиганда.[58]
TMPyP4, катионный порфирин, является более известным лигандом связывания G4, который помогает подавлять c-Myc. Способ, которым TMPyP4 связывается с G4, аналогичен MM41, с кольцом, укладывающимся на внешний G-квартет, а боковые цепи связаны с петлями G4.[59]
При конструировании лигандов для связывания с G-квадруплексами лиганды имеют более высокое сродство к параллельно сложенным G-квадруплексам. Было обнаружено, что лиганды с меньшими боковыми цепями лучше связываются с квадруплексом, потому что лиганды меньшего размера имеют более концентрированные электронная плотность. Кроме того, водородные связи лигандов с меньшими боковыми цепями короче и, следовательно, прочнее. Лиганды с подвижными боковыми цепями, которые способны вращаться вокруг своего центрального хромофора, сильнее связаны с G-квадруплексами, поскольку конформация петель G4 и боковых цепей лиганда может выравниваться.[60]
Квадруплексные методы прогнозирования
Выявление и прогнозирование последовательностей, способных образовывать квадруплексы, является важным инструментом для дальнейшего понимания их роли. Как правило, для поиска возможных внутрицепочечных образующих квадруплекс последовательностей используется простое сопоставление с образцом: d (G3+N1-7грамм3+N1-7грамм3+N1-7грамм3+), где N - любое нуклеотидное основание (включая гуанин ).[61] Это правило широко используется в он-лайн алгоритмы. Хотя правило эффективно идентифицирует сайты образования G-квадруплекса, оно также определяет подмножество несовершенных гомопуриновых зеркальных повторов, способных образовывать триплекс.[62] и образование i-мотива С-цепи.[63] Более того, эти последовательности также обладают способностью образовывать структуры с проскальзыванием и сгибанием, которые являются неявными промежуточными звеньями в образовании обоих квадруплексов.[4] и триплекс ДНК[64] конструкции. В одном исследовании[65] было обнаружено, что наблюдаемое количество на пару оснований (то есть частота) этих мотивов быстро увеличивалось в Eumetazoa для которых доступны полные геномные последовательности. Это указывает на то, что последовательности могут находиться под положительным отбором, возможным благодаря эволюции систем, способных подавлять образование не-B-структур.
Методы изучения G-квадруплексов
Ряд экспериментальных методов был разработан для поддержки вычислительного предсказания G-квадруплексов. Эти методы можно в общих чертах разделить на два класса: биофизические и биохимические.[66]
Биохимические методы
Биохимические методы были использованы для исследования образования G-квадруплекса в контексте более длинной последовательности. В анализе остановки ДНК-полимеразы образование G-квадруплекса в матрице ДНК может действовать как препятствие и вызывать остановку полимеразы, что останавливает удлинение праймера.[67] Диметилсульфат (DMS) с последующим анализом расщепления пиперидина основан на том факте, что образование G-квадруплекса будет запрещать метилирование гуанина N7, вызванное DMS, что приводит к паттерну защиты, наблюдаемому в области G-квадруплекса ДНК после пиперидина. расщепление.[68]
Биофизические методы
Топология структуры G-квадруплекса может быть определена путем мониторинга сигналов положительного или отрицательного кругового дихроизма (CD) на определенных длинах волн.[69] Параллельные G-квадруплексы имеют отрицательные и положительные сигналы CD на 240 и 262 нм, соответственно, тогда как антипараллельные G-квадруплексы размещают эти сигналы на 262 и 295 нм соответственно. Чтобы проверить образование G-квадруплекса, необходимо также провести эксперименты с CD в условиях стабилизации не-G-квадруплекса (Li +) и стабилизации G-квадруплекса (например, K + или с лигандами G-квадруплекса) и сканировать в сторону дальней УФ-области. (180–230 нм). Точно так же термостабильность структуры G-квадруплекса можно определить, наблюдая УФ-сигнал при 295 нм.[70] При плавлении G-квадруплекса УФ-поглощение при 295 нм уменьшается, что приводит к гипохромному сдвигу, который является отличительной особенностью структуры G-квадруплекса. Другой подход к обнаружению G-квадруплексов включает: нанопора -основанные методы. Во-первых, было показано, что биологические нанопоры может обнаруживать G-квадруплексы на основе исключения размера и специфического взаимодействия G-квадруплекса и белковой нанополости.[71] Новый подход сочетает в себе твердотельные нанопоры и ДНК-нанотехнологии для безметочного обнаружения G-квадруплексов, для их картирования на дцДНК и для мониторинга образования G-квадруплексов.[72]
Роль в неврологических расстройствах
G-квадруплексы вовлечены в неврологические расстройства посредством двух основных механизмов. Первый заключается в увеличении G-повторов внутри генов, что приводит к образованию структур G-квадруплексов, которые непосредственно вызывают заболевание, как в случае с геном C9orf72 и боковой амиотрофический склероз (ALS) или лобно-височная деменция (FTD). Второй механизм связан с мутациями, которые влияют на экспрессию связывающих G-квадруплекс белков, как видно на ломкий X ген умственной отсталости 1 (FMR1) ген и Синдром ломкой Х-хромосомы.[73]
Ген C9orf72 кодирует белок C9orf72 который находится по всему мозгу в нейронный цитоплазма и в пресинаптический терминалы.[74] Мутации гена C9orf72 связаны с развитием FTD и ALS.[75] Эти два заболевания имеют причинную связь с GGGGCC (G4C2) повторяется в пределах 1-го интрон гена C9orf72. Нормальные люди обычно имеют от 2 до 8 G4C2 повторяется, но люди с ЛТД или БАС имеют от 500 до нескольких тысяч G4C2 повторяется.[76][77] Было показано, что транскрибируемая РНК этих повторов формирует стабильные G-квадруплексы, причем данные свидетельствуют о том, что G4C2 Повторы в ДНК также обладают способностью образовывать смешанные параллельно-антипараллельные G-квадруплексные структуры.[78][79] Эти РНК-транскрипты, содержащие G4C2 было показано, что повторы связывают и разделяют широкий спектр белков, включая нуклеолин. Нуклеолин участвует в синтезе и созревании рибосомы внутри ядра, и разделение нуклеолина мутировавшими транскриптами РНК нарушает функцию ядрышка и синтез рибосомной РНК.[80]
Белок умственной отсталости Fragile X (FMRP) - это широко экспрессируемый белок, кодируемый геном FMR1, который связывается с вторичными структурами G-квадруплекса в нейронах и участвует в синаптическая пластичность.[81] FMRP действует как негативный регулятор трансляции, и его связывание стабилизирует структуры G-квадруплексов в транскриптах мРНК, ингибируя рибосомы. удлинение мРНК в нейронах дендрит и контроль времени выражения стенограммы.[82][83] Мутации этого гена могут вызвать развитие синдрома ломкой Х-хромосомы, аутизм и другие неврологические расстройства.[84] В частности, синдром ломкой Х-хромосомы вызван увеличением от 50 до более 200 повторов CGG в пределах экзон 13 гена FMR1. Это повторное расширение способствует Метилирование ДНК и другие эпигенетические гетерохроматин модификации FMR1, которые предотвращают транскрипцию гена, что приводит к патологически низким уровням FMRP.[85][86]
Лечебные подходы
Антисмысловые вмешательства и малая молекула лиганды являются общими стратегиями, используемыми для нацеливания на неврологические заболевания, связанные с повторами экспансии G-квадруплекса. Следовательно, эти методы особенно полезны для лечения неврологических заболеваний, которые имеют механизм усиления функции, когда измененный продукт гена имеет новую функцию или новую экспрессию гена; это было обнаружено в C9orf72 (открытая рамка считывания 72 хромосомы 9).[87]
Антисмысловая терапия это процесс, с помощью которого синтезированные нити нуклеиновых кислот используются для прямого и специфического связывания с мРНК, продуцируемой определенным геном, которая инактивирует его. Антисмысловые олигонуклеотиды (ASO) обычно используются для нацеливания на РНК C9orf72 области повторения экспансии G-квадруплекса GGGGCC, что снижает токсичность в клеточных моделях C9orf72.[88][89][90] ASOs ранее использовались для восстановления нормальных фенотипов при других неврологических заболеваниях, которые имеют механизмы усиления функции, с той лишь разницей, что они использовались в отсутствие областей повторов расширения G-квадруплекса.[91][92][93][94]
Другой широко используемый метод - использование низкомолекулярных соединений. лиганды. Их можно использовать для нацеливания на области G-квадруплекса, вызывающие неврологические расстройства. Существует около 1000 различных лигандов G-квадруплексов, в которых они могут взаимодействовать через свои ароматические кольца; это позволяет низкомолекулярным лигандам складываться на плоских концевых тетрадах в областях G-квадруплекса. Недостатком использования низкомолекулярных лигандов в качестве терапевтического метода является то, что специфичностью трудно управлять из-за изменчивости G-квадруплексов в их первичных последовательностях, ориентации, термодинамической стабильности и стехиометрии цепи нуклеиновой кислоты. На данный момент[когда? ] ни один низкомолекулярный лиганд не был способен быть полностью специфичным для одной последовательности G-квадруплекса.[95][96] Однако катионный порфирин, известный как TMPyP4, способен связываться с повторяющейся областью C9orf72 GGGGCC, что заставляет область повторения G-квадруплекса разворачиваться и терять свои взаимодействия с белками, что приводит к потере функциональности.[97] Низкомолекулярные лиганды, состоящие в основном из свинца, также могут нацеливаться на области повторов GGGGCC и в конечном итоге снижают как ассоциированную с повторами трансляцию не-ATG, так и фокусы РНК в нейронных клетках, полученных от пациентов с Боковой амиотрофический склероз (БАС). Это свидетельствует о том, что низкомолекулярные лиганды являются эффективным и действенным процессом нацеливания на области GGGGCC, и что специфичность связывания низкомолекулярных лигандов является достижимой целью для научного сообщества.
Комплексы металлов обладают рядом свойств, которые делают их особенно подходящими в качестве связывающих веществ G4 ДНК и, следовательно, в качестве потенциальных лекарств. Хотя металл играет в значительной степени структурную роль в большинстве связывающих G4, есть также примеры, когда он непосредственно взаимодействует с G4 посредством электростатических взаимодействий или прямой координации с азотистыми основаниями.[98]
Рекомендации
- ^ Капра, Джон А .; Паешке, Катрин; Сингх, Мона; Закиан, Вирджиния А .; Стормо, Гэри Д. (22 июля 2010 г.). «Последовательности G-квадруплекса ДНК эволюционно консервативны и связаны с определенными геномными особенностями в Saccharomyces cerevisiae». PLOS вычислительная биология. 6 (7): e1000861. Bibcode:2010PLSCB ... 6E0861C. Дои:10.1371 / journal.pcbi.1000861. ЧВК 2908698. PMID 20676380.
- ^ Routh ED, Creacy SD, Beerbower PE, Akman SA, Vaughn JP, Smaldino PJ (март 2017 г.). «Подход сродства к G-квадруплексной ДНК для очистки ферментативно активной резолвазы 1 G4». Журнал визуализированных экспериментов. 121 (121). Дои:10.3791/55496. ЧВК 5409278. PMID 28362374.
- ^ а б Ларджи Э, Мергни Дж, Габелика В (2016). «Глава 7. Роль ионов щелочных металлов в структуре и стабильности G-квадруплексной нуклеиновой кислоты». В Astrid S, Helmut S, Roland KO S (ред.). Ионы щелочных металлов: их роль в жизни. Ионы металлов в науках о жизни. 16. Springer. С. 203–258. Дои:10.1007/978-4-319-21756-7_7 (неактивно 11.11.2020).CS1 maint: DOI неактивен по состоянию на ноябрь 2020 г. (связь)
- ^ а б Сандквист В.И., Клуг А. (декабрь 1989 г.). «Теломерная ДНК димеризуется путем образования гуаниновых тетрад между петлями шпильки». Природа. 342 (6251): 825–9. Bibcode:1989Натура.342..825С. Дои:10.1038 / 342825a0. PMID 2601741. S2CID 4357161.
- ^ а б c Сен Д., Гилберт В. (июль 1988 г.). «Формирование параллельных четырехцепочечных комплексов богатыми гуанином мотивами в ДНК и его значение для мейоза». Природа. 334 (6180): 364–6. Bibcode:1988Натура.334..364S. Дои:10.1038 / 334364a0. PMID 3393228. S2CID 4351855.
- ^ а б c d Равал П., Куммарасетти В.Б., Равиндран Р., Кумар Н., Гальдер К., Шарма Р., Мукерджи М., Дас С.К., Чоудхури С. (2006). «Полногеномное предсказание ДНК G4 как регуляторных мотивов: роль в глобальном регулировании Escherichia Coli». Геномные исследования. 16 (5): 644‐655. Дои:10.1101 / гр.4508806. ЧВК 1457047. PMID 16651665.
- ^ а б c d Борман С (28 мая 2007 г.). «Подъем структур нуклеиновых кислот квадруплексов становится перспективной лекарственной мишенью». Новости химии и техники. 85 (22): 12–17. Дои:10.1021 / cen-v085n009.p012a.
- ^ Верма А., Гальдер К., Хальдер Р., Ядав В.К., Равал П., Такур Р.К., Мохд Ф., Шарма А., Чоудхури С. (2008). «Полногеномный вычислительный и экспрессионный анализ выявляет мотивы G-квадруплекса ДНК как консервативные цис-регуляторные элементы у человека и родственных видов». Журнал медицинской химии. 51 (18): 5641‐5649. Дои:10.1021 / jm800448a. PMID 18767830.
- ^ Хан Х, Херли ЛХ (апрель 2000 г.). «G-квадруплекс ДНК: потенциальная мишень для разработки противораковых лекарств». Тенденции в фармакологических науках. 21 (4): 136–42. Дои:10.1016 / s0165-6147 (00) 01457-7. PMID 10740289.
- ^ Бохман М.Л., Паешке К., Закян В.А. (ноябрь 2012 г.). «Вторичные структуры ДНК: стабильность и функция G-квадруплексных структур». Обзоры природы. Генетика. 13 (11): 770–80. Дои:10.1038 / nrg3296. ЧВК 3725559. PMID 23032257.
- ^ Ядав В.К., Абрахам Дж. К., Мани П., Кульшреста Р., Чоудхури С. (2008). «QuadBase: полногеномная база данных ДНК G4 - наличие и сохранение в промоторах человека, шимпанзе, мыши и крысы и 146 микробах». Исследования нуклеиновых кислот. 36 (База данных): D381 ‐ D385. Дои:10.1093 / нар / гкм781. ЧВК 2238983. PMID 17962308.
- ^ Дхапола П., Чоудхури С. (июль 2016 г.). "QuadBase2: веб-сервер для мультиплексного анализа и визуализации гуаниновых квадруплексов". Исследования нуклеиновых кислот. 44 (W1): W277 ‐ W283. Дои:10.1093 / нар / gkw425. ЧВК 4987949. PMID 27185890.
- ^ Родос Д., Липпс Х. Дж. (Октябрь 2015 г.). «G-квадруплексы и их регуляторные роли в биологии». Исследования нуклеиновых кислот. 43 (18): 8627–37. Дои:10.1093 / нар / gkv862. ЧВК 4605312. PMID 26350216.
- ^ Борман С (ноябрь 2009 г.). «Промоторные квадруплексы, свернутые в структуры ДНК в сайтах активации генов, могут быть полезными мишенями для лечения рака». Новости химии и техники. 87 (44): 28–30. Дои:10.1021 / cen-v087n044.p028.
- ^ Геллерт М., Липсетт М.Н., Дэвис Д.Р. (декабрь 1962 г.). «Образование спирали гуаниловой кислотой». Труды Национальной академии наук Соединенных Штатов Америки. 48 (12): 2013–8. Bibcode:1962ПНАС ... 48.2013Г. Дои:10.1073 / пнас.48.12.2013. ЧВК 221115. PMID 13947099.
- ^ Хендерсон Э., Хардин К.С., Уолк С.К., Тиноко I, Блэкберн Э.Х. (декабрь 1987 г.). «Теломерные олигонуклеотиды ДНК образуют новые внутримолекулярные структуры, содержащие пары оснований гуанин-гуанин». Клетка. 51 (6): 899–908. Дои:10.1016/0092-8674(87)90577-0. PMID 3690664. S2CID 37343642.
- ^ Симонссон Т. (апрель 2001 г.). «G-квадруплексные структуры ДНК - вариации на тему». Биологическая химия. 382 (4): 621–8. Дои:10.1515 / BC.2001.073. PMID 11405224. S2CID 43536134.
- ^ Бердж С., Паркинсон Г. Н., Хейзел П., Тодд А. К., Нейдл С. (2006). «Квадруплексная ДНК: последовательность, топология и структура». Исследования нуклеиновых кислот. 34 (19): 5402–15. Дои:10.1093 / нар / gkl655. ЧВК 1636468. PMID 17012276.
- ^ Цао К., Рывкин П., Джонсон Ф. Б. (май 2012 г.). «Вычислительное обнаружение и анализ последовательностей с дуплексным межцепочечным потенциалом образования G-квадруплекса». Методы. 57 (1): 3–10. Дои:10.1016 / j.ymeth.2012.05.002. ЧВК 3701776. PMID 22652626.
- ^ Кудлицкий А.С. (2016). «G-квадруплексы, включающие обе нити геномной ДНК, очень многочисленны и колокализуются с функциональными участками в геноме человека». PLOS ONE. 11 (1): e0146174. Bibcode:2016PLoSO..1146174K. Дои:10.1371 / journal.pone.0146174. ЧВК 4699641. PMID 26727593.
- ^ Мурат П., Баласубраманян С. (апрель 2014 г.). «Существование и последствия G-квадруплексных структур в ДНК». Текущее мнение в области генетики и развития. 25 (25): 22–9. Дои:10.1016 / j.gde.2013.10.012. PMID 24584093.
- ^ Миёси Д., Каримата Х., Сугимото Н. (июнь 2006 г.). «Гидратация регулирует термодинамику образования G-квадруплексов в условиях молекулярного краудинга». Журнал Американского химического общества. 128 (24): 7957–63. Дои:10.1021 / ja061267m. PMID 16771510.
- ^ Чжэн К.В., Чен З., Хао Ю.Х., Тан З. (январь 2010 г.). «Молекулярное скопление создает необходимую среду для образования стабильных G-квадруплексов в длинной двухцепочечной ДНК». Исследования нуклеиновых кислот. 38 (1): 327–38. Дои:10.1093 / nar / gkp898. ЧВК 2800236. PMID 19858105.
- ^ Эндох Т., Роде А.Б., Такахаши С., Катаока Ю., Кувахара М., Сугимото Н. (февраль 2016 г.). «Мониторинг образования G-квадруплекса во время транскрипции в реальном времени». Аналитическая химия. 88 (4): 1984–9. Дои:10.1021 / acs.analchem.5b04396. PMID 26810457.
- ^ Ван Цзянь, Лю Дж.К., Чен З, Чжэн К.В., Чен Си, Хао Й., Тан З. (август 2011 г.). «Образование G-квадруплекса на 3'-конце теломерной ДНК ингибирует его удлинение теломеразой, полимеразой и раскручивание геликазой». Исследования нуклеиновых кислот. 39 (14): 6229–37. Дои:10.1093 / nar / gkr164. ЧВК 3152327. PMID 21441540.
- ^ Schaffitzel C, Berger I, Postberg J, Hanes J, Lipps HJ, Plückthun A (июль 2001 г.). «Созданные in vitro антитела, специфичные для теломерной гуанин-квадруплексной ДНК, реагируют с макронуклеарами Stylonychia lemnae». Труды Национальной академии наук Соединенных Штатов Америки. 98 (15): 8572–7. Bibcode:2001PNAS ... 98.8572S. Дои:10.1073 / pnas.141229498. ЧВК 37477. PMID 11438689.
- ^ Паешке К., Симонссон Т., Постберг Дж., Родос Д., Липпс Х.Д. (октябрь 2005 г.). «Белки, связывающие концы теломер, контролируют образование структур G-квадруплекса ДНК in vivo». Структурная и молекулярная биология природы. 12 (10): 847–54. Дои:10.1038 / nsmb982. PMID 16142245. S2CID 6079323.
- ^ Кар, Анирбан; Джонс, Натан; Арат, Н. Озлем; Фишел, Ричард; Гриффит, Джек Д. (15.06.2018). «Длинно повторяющаяся (TTAGGG) n одноцепочечная ДНК самоконденсируется в компактные бисерные филаменты, стабилизированные образованием G-квадруплекса». Журнал биологической химии. 293 (24): 9473–9485. Дои:10.1074 / jbc.RA118.002158. ISSN 0021-9258. ЧВК 6005428. PMID 29674319.
- ^ Simonsson T, Pecinka P, Kubista M (март 1998 г.). «Образование тетраплекса ДНК в контрольной области c-myc». Исследования нуклеиновых кислот. 26 (5): 1167–72. Дои:10.1093 / nar / 26.5.1167. ЧВК 147388. PMID 9469822.
- ^ Сиддики-Джайн А., Grand CL, Bearss DJ, Hurley LH (сентябрь 2002 г.). «Прямое доказательство наличия G-квадруплекса в промоторной области и его нацеливания с помощью небольшой молекулы для репрессии транскрипции c-MYC». Труды Национальной академии наук Соединенных Штатов Америки. 99 (18): 11593–8. Bibcode:2002PNAS ... 9911593S. Дои:10.1073 / pnas.182256799. ЧВК 129314. PMID 12195017.
- ^ Huppert JL, Balasubramanian S (14 декабря 2006 г.). «G-квадруплексы в промоторах по всему геному человека». Исследования нуклеиновых кислот. 35 (2): 406–13. Дои:10.1093 / нар / gkl1057. ЧВК 1802602. PMID 17169996.
- ^ Дай Дж, Дексхаймер Т.С., Чен Д., Карвер М., Амбрус А., Джонс Р.А., Ян Д. (февраль 2006 г.). «Внутримолекулярная структура G-квадруплекса со смешанными параллельными / антипараллельными G-цепями, образованная в области промотора BCL-2 человека в растворе». Журнал Американского химического общества. 128 (4): 1096–8. Дои:10.1021 / ja055636a. ЧВК 2556172. PMID 16433524.
- ^ а б Фернандо Х., Решка А.П., Хупперт Дж., Ладам С., Ранкин С., Венкитараман А.Р., Нейдл С., Баласубраманиан С. (июнь 2006 г.). «Консервативный квадруплексный мотив, расположенный в сайте активации транскрипции онкогена c-kit человека». Биохимия. 45 (25): 7854–60. Дои:10.1021 / bi0601510. ЧВК 2195898. PMID 16784237.
- ^ Хупперт Дж. Л., Баласубраманиан С. (2005). «Преобладание квадруплексов в геноме человека». Исследования нуклеиновых кислот. 33 (9): 2908–16. Дои:10.1093 / нар / gki609. ЧВК 1140081. PMID 15914667.
- ^ Равал П., Куммарасетти В. Б., Равиндран Дж., Кумар Н., Гальдер К., Шарма Р., Мукерджи М., Дас С. К., Чоудхури С. (май 2006 г.). «Общегеномное предсказание ДНК G4 как регуляторных мотивов: роль в глобальной регуляции Escherichia coli». Геномные исследования. 16 (5): 644–55. Дои:10.1101 / гр.4508806. ЧВК 1457047. PMID 16651665.
- ^ Камат-Леб А., Леб Л.А., Фрай М. (2012). Коттерилл С (ред.). «Белок синдрома Вернера отличается от белка синдрома Блума своей способностью прочно связывать различные структуры ДНК». PLOS ONE. 7 (1): e30189. Bibcode:2012PLoSO ... 730189K. Дои:10.1371 / journal.pone.0030189. ЧВК 3260238. PMID 22272300.
- ^ Вон Дж. П., Creacy SD, Routh ED, Joyner-Butt C, Jenkins GS, Pauli S, Nagamine Y, Akman SA (ноябрь 2005 г.). «Белковый продукт DEXH гена DHX36 является основным источником разрешающей активности тетрамолекулярного квадруплекса G4-ДНК в лизатах клеток HeLa». Журнал биологической химии. 280 (46): 38117–20. Дои:10.1074 / jbc.C500348200. PMID 16150737.
- ^ Чен М.С., Ферре-Д'Амаре, АР (15 августа 2017 г.). «Структурные основы активности геликазы DEAH / RHA». Кристаллы. 7 (8): 253. Дои:10,3390 / крист7080253.
- ^ а б Такур Р.К., Кумар П., Гальдер К., Верма А., Кар А., Родитель Д.Л., Басундра Р., Кумар А., Чоудхури С. (январь 2009 г.). "Взаимодействие супрессора метастазов NM23-H2 с G-квадруплексной ДНК в гиперчувствительном элементе нуклеазы промотора c-MYC, индуцирующем экспрессию c-MYC". Nucleic Acid Reseacrh. 37 (1): 172‐183. Дои:10.1093 / nar / gkn919. ЧВК 2615625. PMID 19033359.
- ^ Борман С (ноябрь 2009 г.). «Промоторные квадруплексы, свернутые структуры ДНК в сайтах активации генов, могут быть полезными мишенями для лечения рака». Новости химии и техники. 87 (44): 28–30. Дои:10.1021 / cen-v087n044.p028.
- ^ а б c Саха Д., Сингх А., Хуссейн Т., Шривастава В., Сенгупта С., Кар А., Дхапола П., Умманни Р., Чоудхури С. (июль 2017 г.). «Эпигенетическое подавление теломеразы человека (hTERT) опосредовано супрессором метастазов NME2 в зависимости от G-квадруплекса». Журнал биологической химии. 292 (37): 15205‐15215. Дои:10.1074 / jbc.M117.792077. ЧВК 5602382. PMID 28717007.
- ^ Мукерджи А.К., Шарма С., Багри С., Кутум Р., Кумар П., Хуссейн А., Сингх П., Саха Д., Кар А., Даш Д., Чоудхури С. (ноябрь 2019 г.). «Фактор связывания теломерных повторов 2 широко связывается с экстра-теломерными G-квадруплексами и регулирует эпигенетический статус нескольких промоторов генов». Журнал биологической химии. 294 (47): 17709–17722. Дои:10.1074 / jbc.RA119.008687. ЧВК 6879327. PMID 31575660.
- ^ Майзелс Н, Грей LT (апрель 2013 г.). Розенберг С.М. (ред.). «Геном G4». PLOS Genetics. 9 (4): e1003468. Дои:10.1371 / journal.pgen.1003468. ЧВК 3630100. PMID 23637633.
- ^ Биффи Дж., Таннахилл Д., Маккафферти Дж., Баласубраманиан С. (март 2013 г.). «Количественная визуализация структур G-квадруплексов ДНК в клетках человека». Химия природы. 5 (3): 182–6. Bibcode:2013НатЧ ... 5..182Б. Дои:10.1038 / nchem.1548. ЧВК 3622242. PMID 23422559.
- ^ Chen MC, Tippana R, Demeshkina NA, Murat P, Balasubramanian S, Myong S, Ferré-D'Amaré AR (июнь 2018 г.). «Структурная основа разворачивания G-квадруплекса с помощью DEAH / RHA геликазы DHX36». Природа. 558 (7710): 465–469. Bibcode:2018Натура.558..465C. Дои:10.1038 / s41586-018-0209-9. ЧВК 6261253. PMID 29899445.
- ^ Рис C., Skordalakes E (2016). «Строение и функции теломерного комплекса CST». Журнал вычислительной и структурной биотехнологии. 14: 161–7. Дои:10.1016 / j.csbj.2016.04.002. ЧВК 4872678. PMID 27239262.
- ^ Neidle S (июль 2016 г.). «Квадруплексные нуклеиновые кислоты как новые терапевтические мишени» (PDF). Журнал медицинской химии. 59 (13): 5987–6011. Дои:10.1021 / acs.jmedchem.5b01835. PMID 26840940.
- ^ Чен Й, Ян Д. (сентябрь 2012 г.). Последовательность, стабильность и структура G-квадруплексов и их взаимодействие с лекарствами. Текущие протоколы в химии нуклеиновых кислот. Глава 17. С. 17.5.1–17.5.17. Дои:10.1002 / 0471142700.nc1705s50. ISBN 978-0471142706. ЧВК 3463244. PMID 22956454.
- ^ Брукс Т.А., Кендрик С., Херли Л. (сентябрь 2010 г.). «Понимание функций G-квадруплекса и i-мотива в промоторах онкогенов». Журнал FEBS. 277 (17): 3459–69. Дои:10.1111 / j.1742-4658.2010.07759.x. ЧВК 2971675. PMID 20670278.
- ^ а б Ou TM, Lin J, Lu YJ, Hou JQ, Tan JH, Chen SH, Li Z, Li YP, Li D, Gu LQ, Huang ZS (август 2011 г.). «Ингибирование пролиферации клеток производным хиндолина (SYUIQ-05) за счет его предпочтительного взаимодействия с промотором c-myc G-quadruplex». Журнал медицинской химии. 54 (16): 5671–9. Дои:10.1021 / jm200062u. PMID 21774525.
- ^ Шарма С., Мукерджи А. К., Рой С. С., Багри С., Лир С., Верма М., Сенгупта А., Кумар М., Нессе Г., Панди Д. П., Чоудхури С. (январь 2020 г.). «Экспрессия теломеразы человека находится под прямым транскрипционным контролем теломер-связывающего фактора TRF2». bioRxiv. Дои:10.1101/2020.01.15.907626. S2CID 214472968.
- ^ Сунь Д., Го К., Руше Дж. Дж., Херли Л. Х. (2005-10-12). «Содействие структурному переходу в тракте полипурин / полипиримидин в проксимальной промоторной области гена VEGF человека за счет присутствия взаимодействующих с калием и G-квадруплекс агентов». Исследования нуклеиновых кислот. 33 (18): 6070–80. Дои:10.1093 / нар / gki917. ЧВК 1266068. PMID 16239639.
- ^ Хуссейн Т., Саха Д., Пурохит Дж., Мукерджи А. К., Шарма С., Сенгупта С., Дхапола П., Маджи Б., Ведагопурам С., Хорикоши Н. Т., Хорикоши Н., Пандита Р. К., Бхаттачарья С., Баджадж А., Рио Дж. Ф., Пандита С. К., Чоудури (Сентябрь 2017 г.). «Регуляция транскрипции CDKN1A (p21 / CIP1 / WAF1) с помощью TRF2 эпигенетически контролируется посредством репрессорного комплекса REST». Научные отчеты. 7 (1): 11541. Bibcode:2017НатСР ... 711541H. Дои:10.1038 / s41598-017-11177-1. ЧВК 5599563. PMID 28912501.
- ^ Де Армонд Р., Вуд С., Сан Д., Херли Л. Х., Эббингаус ЮЗ (декабрь 2005 г.). «Доказательства присутствия области, образующей квадруплекс гуанина в полипуриновом тракте промотора фактора 1альфа, индуцируемого гипоксией». Биохимия. 44 (49): 16341–50. Дои:10.1021 / bi051618u. PMID 16331995.
- ^ Гуо К., Поурпак А., Битц-Роджерс К., Гокхале В., Сан Д., Херли Л.Х. (август 2007 г.). «Формирование псевдосимметричных структур G-квадруплекса и i-мотива в проксимальной области промотора онкогена RET». Журнал Американского химического общества. 129 (33): 10220–8. Дои:10.1021 / ja072185g. ЧВК 2566970. PMID 17672459.
- ^ Цинь И, Резлер Э.М., Гокхале В., Сан Д., Херли Л.Х. (26 ноября 2007 г.). «Характеристика G-квадруплексов в сверхчувствительном элементе дуплексной нуклеазы промотора PDGF-A и модуляции активности промотора PDGF-A с помощью TMPyP4». Исследования нуклеиновых кислот. 35 (22): 7698–713. Дои:10.1093 / нар / гкм538. ЧВК 2190695. PMID 17984069.
- ^ Чилакамарти Ю., Котешвар Д., Джинка С., Вамси Кришна Н., Шридхаран К., Нагеш Н., Гирибабу Л. (ноябрь 2018 г.). «Новый амфифильный G-квадруплекс, связывающий синтетическое производное TMPyP4, и его влияние на пролиферацию раковых клеток и индукцию апоптоза». Биохимия. 57 (46): 6514–6527. Дои:10.1021 / acs.biochem.8b00843. PMID 30369235.
- ^ Ohnmacht SA, Marchetti C, Gunaratnam M, Besser RJ, Haider SM, Di Vita G, Lowe HL, Mellinas-Gomez M, Diocou S, Robson M, Šponer J, Islam B, Pedley RB, Hartley JA, Neidle S (июнь 2015 г. ). «Соединение, связывающееся с G-квадруплексом, проявляющее противоопухолевую активность на модели рака поджелудочной железы in vivo». Научные отчеты. 5: 11385. Bibcode:2015НатСР ... 511385O. Дои:10.1038 / srep11385. ЧВК 4468576. PMID 26077929.
- ^ Сиддики-Джайн А., Grand CL, Bearss DJ, Hurley LH (сентябрь 2002 г.). «Прямое доказательство наличия G-квадруплекса в промоторной области и его нацеливания с помощью небольшой молекулы для репрессии транскрипции c-MYC». Труды Национальной академии наук Соединенных Штатов Америки. 99 (18): 11593–8. Bibcode:2002PNAS ... 9911593S. Дои:10.1073 / pnas.182256799. ЧВК 129314. PMID 12195017.
- ^ Колли Г.В., Промонторио Р., Хампель С.М., Микко М., Нейдл С., Паркинсон Г.Н. (февраль 2012 г.). «Структурная основа для нацеливания теломерного G-квадруплекса с помощью нафталиндиимидных лигандов». Журнал Американского химического общества. 134 (5): 2723–31. Дои:10.1021 / ja2102423. PMID 22280460.
- ^ Тодд А.К., Джонстон М., Нейдл С. (2005). «Широко распространенные предполагаемые мотивы квадруплексной последовательности в ДНК человека». Исследования нуклеиновых кислот. 33 (9): 2901–7. Дои:10.1093 / нар / gki553. ЧВК 1140077. PMID 15914666.
- ^ Франк-Каменецкий М.Д., Миркин С.М. (1995). «Триплексные структуры ДНК». Ежегодный обзор биохимии. 64 (9): 65–95. Дои:10.1146 / annurev.bi.64.070195.000433. PMID 7574496.
- ^ Гуо К., Гокхале В., Херли Л.Х., Сунь Д. (август 2008 г.). «Внутримолекулярно свернутые структуры G-квадруплексов и i-мотивов в проксимальном промоторе гена фактора роста эндотелия сосудов». Исследования нуклеиновых кислот. 36 (14): 4598–608. Дои:10.1093 / нар / gkn380. ЧВК 2504309. PMID 18614607.
- ^ Миркин С.М., Лямичев В.И., Друшляк К.Н., Добрынин В.Н., Филиппов С.А., Франк-Каменецкий М.Д. (1987). «Форма ДНК H требует зеркального повторения гомопурин-гомопиримидин». Природа. 330 (6147): 495–7. Bibcode:1987Натура.330..495М. Дои:10.1038 / 330495a0. PMID 2825028. S2CID 4360764.
- ^ Смит СС (2010). «Эволюционное расширение структурно сложных последовательностей ДНК». Геномика и протеомика рака. 7 (4): 207–15. PMID 20656986.
- ^ Квок, Чун Кит; Меррик, Кэтрин Дж. (2017-10-01). «G-квадруплексы: прогноз, характеристика и биологическое применение». Тенденции в биотехнологии. 35 (10): 997–1013. Дои:10.1016 / j.tibtech.2017.06.012. ISSN 0167-7799. PMID 28755976.
- ^ Han, H .; Hurley, L.H .; Салазар, М. (1999-01-01). «Анализ остановки ДНК-полимеразы для соединений, взаимодействующих с G-квадруплексом». Исследования нуклеиновых кислот. 27 (2): 537–542. Дои:10.1093 / nar / 27.2.537. ISSN 0305-1048. ЧВК 148212. PMID 9862977.
- ^ Солнце, Дээкю; Херли, Лоуренс Х. (2009-10-23), Биохимические методы для характеристики структур G-квадруплексов: EMSA, DMS Footprinting и ДНК-полимеразный стоп-анализ, Методы молекулярной биологии, 608, Humana Press, стр. 65–79, Дои:10.1007/978-1-59745-363-9_5, ISBN 9781588299505, ЧВК 2797547, PMID 20012416
- ^ Парамасиван, Саттанатан; Ружан, Юлиан; Болтон, Филип Х. (2007-12-01). «Круговой дихроизм квадруплексных ДНК: приложения к структуре, катионным эффектам и связыванию лигандов». Методы. Квадруплексная ДНК. 43 (4): 324–331. Дои:10.1016 / j.ymeth.2007.02.009. ISSN 1046-2023. PMID 17967702.
- ^ Мерни, Жан-Луи; Фан, Ань-Туан; Лакруа, Лоран (11 сентября 1998). «После формирования G-квартета методом УФ-спектроскопии». Письма FEBS. 435 (1): 74–78. Дои:10.1016 / s0014-5793 (98) 01043-6. ISSN 0014-5793. PMID 9755862. S2CID 1306129.
- ^ Анна; Флеминг, Аарон М .; Миддлтон, Эрик Дж .; Берроуз, Синтия Дж. (2014-09-15). «Одномолекулярное исследование складок G-квадруплекса теломерной последовательности человека в белковой нанополости». Труды Национальной академии наук. 111 (40): 14325–14331. Bibcode:2014ПНАС..11114325А. Дои:10.1073 / pnas.1415944111. ISSN 0027-8424. ЧВК 4209999. PMID 25225404.
- ^ Бошкович, Филип; Чжу, Дзинбо; Чен, Кайкай; Кейзер, Ульрих Ф. (02.10.2019). «Мониторинг образования G-квадруплексов с ДНК-носителями и твердотельными нанопорами». Нано буквы. 19 (11): 7996–8001. Bibcode:2019NanoL..19.7996B. Дои:10.1021 / acs.nanolett.9b03184. ISSN 1530-6984. PMID 31577148.
- ^ Симона Р., Фратта П., Нейдл С., Паркинсон Г. Н., Исаакс А. М. (июнь 2015 г.). «G-квадруплексы: новые роли в нейродегенеративных заболеваниях и некодирующий транскриптом». Письма FEBS. 589 (14): 1653–68. Дои:10.1016 / j.febslet.2015.05.003. PMID 25979174.
- ^ Открытая рамка считывания 72 хромосомы 9 C9orf72 [Homo sapiens] - Ген - NCBI
- ^ Ратнавалли Е., Брейн С., Доусон К., Ходжес-младший (июнь 2002 г.). «Распространенность лобно-височной деменции». Неврология. 58 (11): 1615–21. Дои:10.1212 / WNL.58.11.1615. PMID 12058088. S2CID 45904851.
- ^ Резерфорд, штат Нью-Джерси, Хекман М.Г., Дехесус-Эрнандес М., Бейкер М.С., Сото-Ортолаза А.И., Райапролу С., Стюарт Х., Фингер Э, Фолькенинг К., Сили У. , Knopman DS, Kretzschmar HA, Neumann M, Caselli RJ, White CL, Mackenzie IR, Petersen RC, Strong MJ, Miller BL, Boeve BF, Uitti RJ, Boylan KB, Wszolek ZK, Graff-Radford NR, Dickson DW, Ross OA , Радемакерс Р. (декабрь 2012 г.). «Длина нормальных аллелей повтора C9ORF72 GGGGCC не влияет на фенотип заболевания». Нейробиология старения. 33 (12): 2950.e5–7. Дои:10.1016 / j.neurobiolaging.2012.07.005. ЧВК 3617405. PMID 22840558.
- ^ Бек Дж., Поултер М., Хенсман Д., Рорер Дж. Д., Махони Си Джей, Адамсон Дж., Кэмпбелл Т., Апхилл Дж., Борг А., Фратта П., Оррелл Р. У., Маласпина А., Роу Дж., Браун Дж., Ходжес Дж., Сайдл К., Полке Дж. М. , Houlden H, Schott JM, Fox NC, Rossor MN, Tabrizi SJ, Isaacs AM, Hardy J, Warren JD, Collinge J, Mead S (март 2013 г.). «Большие экспансии гексануклеотидных повторов C9orf72 наблюдаются при множественных нейродегенеративных синдромах и встречаются чаще, чем ожидалось, в популяции Великобритании». Американский журнал генетики человека. 92 (3): 345–53. Дои:10.1016 / j.ajhg.2013.01.011. ЧВК 3591848. PMID 23434116.
- ^ Fratta P, Mizielinska S, Nicoll AJ, Zloh M, Fisher EM, Parkinson G, Isaacs AM (декабрь 2012 г.). «Гексануклеотидный повтор C9orf72, связанный с боковым амиотрофическим склерозом и лобно-височной деменцией, образует G-квадруплексы РНК». Научные отчеты. 2: 1016. Bibcode:2012НатСР ... 2Э1016Ф. Дои:10.1038 / srep01016. ЧВК 3527825. PMID 23264878.
- ^ Редди К., Замири Б., Стэнли С.И., Макгрегор Р.Б., Пирсон К.Э. (апрель 2013 г.). «Связанный с заболеванием повтор r (GGGGCC) n из гена C9orf72 образует зависимые от длины тракта одно- и многомолекулярные G-квадруплексные структуры РНК». Журнал биологической химии. 288 (14): 9860–6. Дои:10.1074 / jbc.C113.452532. ЧВК 3617286. PMID 23423380.
- ^ Haeusler AR, Donnelly CJ, Periz G, Simko EA, Shaw PG, Kim MS, Maragakis NJ, Troncoso JC, Pandey A, Sattler R, Rothstein JD, Wang J (март 2014 г.). «Структуры нуклеотидных повторов C9orf72 инициируют молекулярные каскады болезней». Природа. 507 (7491): 195–200. Bibcode:2014Натура.507..195H. Дои:10.1038 / природа13124. ЧВК 4046618. PMID 24598541.
- ^ Дарнелл, Дж. К., Дженсен, К. Б., Джин, П., Браун, В., Уоррен, С. Т., Дарнелл. Р. Б. (ноябрь 2001 г.). "Fragile X Mental Retardation Protein Targets G Quartet mRNAs important for Neuronal Function". Клетка. 107 (4): 489–499. Дои:10.1016 / S0092-8674 (01) 00566-9. PMID 11719189. S2CID 8203054.CS1 maint: несколько имен: список авторов (связь)
- ^ Семан С., О'Доннелл В. Т., Рид М., Паттон С., Поль Дж., Уоррен С. Т. (декабрь 2003 г.). «Фосфорилирование влияет на состояние трансляции FMRP-ассоциированных полирибосом». Молекулярная генетика человека. 12 (24): 3295–305. Дои:10,1093 / hmg / ddg350. PMID 14570712.
- ^ Фэлинг М., Мровка Р., Стиге А., Киршнер К.М., Бенко Э., Ферстера Б., Перссон ПБ, Тиле Б.Дж., Мейер Дж.С., Шольц Х. (февраль 2009 г.). «Трансляционная регуляция гомолога-1 achaete-scute человека с помощью ломкого X-белка умственной отсталости». Журнал биологической химии. 284 (7): 4255–66. Дои:10.1074 / jbc.M807354200. PMID 19097999.
- ^ "Хрупкая умственная отсталость X" Сборник человеческих генов
- ^ Пьеретти, М., Чжан, Ф., Фу, Ю., Уоррен, С. Т., Остра, Б. А., Каски, К. Т., Нельсон, Д. Л. (август 1991 г.). «Отсутствие экспрессии гена FMR-1 при синдроме ломкой Х-хромосомы». Клетка. 66 (4): 816–822. Дои:10.1016 / 0092-8674 (91) 90125-И. PMID 1878973. S2CID 31455523.CS1 maint: несколько имен: список авторов (связь)
- ^ Сатклифф Дж. С., Нельсон Д. Л., Чжан Ф., Пьеретти М., Каски К. Т., Сакс Д., Уоррен С. Т. (сентябрь 1992 г.). «Метилирование ДНК подавляет транскрипцию FMR-1 при синдроме ломкой Х-хромосомы». Молекулярная генетика человека. 1 (6): 397–400. Дои:10,1093 / чмг / 1,6,397. PMID 1301913.
- ^ Mizielinska S, Isaacs AM (октябрь 2014 г.). "C9orf72 боковой амиотрофический склероз и лобно-височная деменция: усиление или потеря функции?". Текущее мнение в неврологии. 27 (5): 515–23. Дои:10.1097 / WCO.0000000000000130. ЧВК 4165481. PMID 25188012.
- ^ Доннелли С.Дж., Чжан П.В., Фам Д.Т., Хойслер А.Р., Хойслер А.Р., Мистри Н.А., Виденски С., Дейли Э.Л., Пот Э.М., Гувер Б., Штраф Д.М., Марагакис Н., Тиенари П.Дж., Петручелли Л., Трейнор Б. , Беннетт С.Ф., Блэкшоу С., Саттлер Р., Ротштейн Д.Д. (октябрь 2013 г.). «Токсичность РНК от экспансии ALS / FTD C9ORF72 смягчается антисмысловым вмешательством». Нейрон. 80 (2): 415–28. Дои:10.1016 / j.neuron.2013.10.015. ЧВК 4098943. PMID 24139042.
- ^ Lagier-Tourenne C, Baughn M, Rigo F, Sun S, Liu P, Li HR, Jiang J, Watt AT, Chun S, Katz M, Qiu J, Sun Y, Ling SC, Zhu Q, Polymenidou M, Drenner K, Artates JW, McAlonis-Downes M, Markmiller S, Hutt KR, Pizzo DP, Cady J, Harms MB, Baloh RH, Vandenberg SR, Yeo GW, Fu XD, Bennett CF, Cleveland DW, Ravits J (ноябрь 2013 г.). «Направленная деградация смысловых и антисмысловых фокусов РНК C9orf72 как терапия для лечения БАС и лобно-височной дегенерации». Труды Национальной академии наук Соединенных Штатов Америки. 110 (47): E4530–9. Bibcode:2013PNAS..110E4530L. Дои:10.1073 / pnas.1318835110. ЧВК 3839752. PMID 24170860.
- ^ Сарин Д., О'Рурк Дж. Г., Мира П., Мухаммад А. К., Грант С., Симпкинсон М., Белл С., Кармона С., Орнелас Л., Сахабиан А., Гендрон Т., Петручелли Л., Бон М., Равиц Дж., Хармс МБ, Риго Ф, Беннетт С.Ф., Отис Т.С., Свендсен С.Н., Бало Р.Х. (октябрь 2013 г.). «Нацеливание на фокусы РНК в двигательных нейронах, происходящих от ИПСК от пациентов с БАС, с увеличением числа повторов C9ORF72». Научная трансляционная медицина. 5 (208): 208ra149. Дои:10.1126 / scitranslmed.3007529. ЧВК 4090945. PMID 24154603.
- ^ Wheeler TM, Leger AJ, Pandey SK, MacLeod AR, Nakamori M, Cheng SH, Wentworth BM, Bennett CF, Thornton CA (август 2012 г.). «Нацеливание на ядерную РНК для коррекции миотонической дистрофии in vivo». Природа. 488 (7409): 111–5. Bibcode:2012Натура.488..111Вт. Дои:10.1038 / природа11362. ЧВК 4221572. PMID 22859208.
- ^ Ли Дж. Э., Беннетт С.Ф., Купер Т.А. (март 2012 г.). «Опосредованная РНКазой Н деградация токсической РНК при миотонической дистрофии 1 типа». Труды Национальной академии наук Соединенных Штатов Америки. 109 (11): 4221–6. Bibcode:2012ПНАС..109.4221Л. Дои:10.1073 / pnas.1117019109. ЧВК 3306674. PMID 22371589.
- ^ Кэрролл Дж. Б., Варби СК, Саутвелл А. Л., Доти С. Н., Гринли С., Скотт Н., Хунг Г., Беннетт С. Ф., Фрейер С. М., Хайден М. Р. (декабрь 2011 г.). «Мощные и селективные антисмысловые олигонуклеотиды, нацеленные на однонуклеотидные полиморфизмы в гене болезни Хантингтона / аллель-специфическое молчание мутантного хантингтина». Молекулярная терапия. 19 (12): 2178–85. Дои:10.1038 / мт.2011.201. ЧВК 3242664. PMID 21971427.
- ^ Gagnon KT, Pendergraff HM, Deleavey GF, Swayze EE, Potier P, Randolph J, Roesch EB, Chattopadhyaya J, Damha MJ, Bennett CF, Montaillier C, Lemaitre M, Corey DR (ноябрь 2010 г.). «Аллель-селективное ингибирование экспрессии мутантного хантингтина с антисмысловыми олигонуклеотидами, нацеленными на увеличенный повтор CAG». Биохимия. 49 (47): 10166–78. Дои:10.1021 / bi101208k. ЧВК 2991413. PMID 21028906.
- ^ Кэмпбелл Н.Х., Патель М. и др. (2009). Селективное распознавание лигандов петель G-квадруплекса. Biochem. 48. 1675-1680.
- ^ Онмахт, С.А., Нейдл, С. (2014). Открытие целевых препаратов с низкомолекулярными квадруплексами. Биоорг. Med. Chem. Lett. 24. 2602-2612.
- ^ Замири Б., Редди К. и др. al. (2014). Порфирин TMPyP4 искажает структуры G-квадруплекса РНК ассоциированного с заболеванием r (GGGGCC) n-повтора гена C9orf72 и блокирует взаимодействия РНК-связывающих белков. J. Biol. Chem. 289. 4653-4659.
- ^ Вилар Р (2018). «Глава 12. Квадруплексы нуклеиновых кислот и металло-препараты». В Sigel A, Sigel H, Freisinger E, Sigel RK (ред.). Металло-препараты: разработка и действие противоопухолевых средств. Ионы металлов в науках о жизни. 18. С. 325–349. Дои:10.1515/9783110470734-018. ISBN 9783110470734. PMID 29394031.
дальнейшее чтение
- Рен Дж, Ван Дж, Хан Л., Ван Е, Ван Дж (октябрь 2011 г.). «Кинетическая прививка G-квадруплексов на наноструктуры ДНК для кодирования структуры и функций с помощью ДНК-машины». Химические коммуникации. 47 (38): 10563–5. Дои:10.1039 / c1cc13973h. PMID 21858307.
- Джонсон Дж. Э., Смит Дж. С., Козак М. Л., Джонсон Ф. Б. (август 2008 г.). «In vivo veritas: использование дрожжей для исследования биологических функций G-квадруплексов». Биохимия. 90 (8): 1250–63. Дои:10.1016 / j.biochi.2008.02.013. ЧВК 2585026. PMID 18331848.
- Хупперт Дж. Л., Баласубраманиан С. (2005). «Преобладание квадруплексов в геноме человека». Исследования нуклеиновых кислот. 33 (9): 2908–16. Дои:10.1093 / нар / gki609. ЧВК 1140081. PMID 15914667.
- Тодд А.К., Джонстон М., Нейдл С. (2005). «Широко распространенные предполагаемые мотивы квадруплексной последовательности в ДНК человека». Исследования нуклеиновых кислот. 33 (9): 2901–7. Дои:10.1093 / нар / gki553. ЧВК 1140077. PMID 15914666.
- Бердж С., Паркинсон Г. Н., Хейзел П., Тодд А. К., Нейдл С. (2006). «Квадруплексная ДНК: последовательность, топология и структура». Исследования нуклеиновых кислот. 34 (19): 5402–15. Дои:10.1093 / нар / gkl655. ЧВК 1636468. PMID 17012276.
- Сиддики-Джайн А., Grand CL, Bearss DJ, Hurley LH (сентябрь 2002 г.). «Прямое доказательство наличия G-квадруплекса в промоторной области и его нацеливания с помощью небольшой молекулы для репрессии транскрипции c-MYC». Труды Национальной академии наук Соединенных Штатов Америки. 99 (18): 11593–8. Bibcode:2002PNAS ... 9911593S. Дои:10.1073 / pnas.182256799. ЧВК 129314. PMID 12195017.
- Равал П., Куммарасетти В. Б., Равиндран Дж., Кумар Н., Гальдер К., Шарма Р., Мукерджи М., Дас С. К., Чоудхури С. (май 2006 г.). «Общегеномное предсказание ДНК G4 как регуляторных мотивов: роль в глобальной регуляции Escherichia coli». Геномные исследования. 16 (5): 644–55. Дои:10.1101 / гр.4508806. ЧВК 1457047. PMID 16651665.
- Хоу Х, Го В, Ся Ф, Не ФК, Донг Х, Тянь И, Вэнь Л, Ван Л, Цао Л, Ян Y, Сюэ Дж, Сон Й, Ван И, Лю Д., Цзян Л. (июнь 2009 г.) "Биомиметический калий-чувствительный наноканал: G-квадруплексное переключение конформации ДНК в синтетической нанопоре". Журнал Американского химического общества. 131 (22): 7800–5. Дои:10.1021 / ja901574c. PMID 19435350.
- Neidle & Balasubramanian, ed. (2006). Квадруплексные нуклеиновые кислоты. ISBN 978-0-85404-374-3. Архивировано из оригинал 30 сентября 2007 г.
- Роуленд Дж. Б., Барнетт К., Дюпон Дж. И., Акурати Дж., Ле В. Х., Льюис Е. А. (декабрь 2013 г.). «Влияние пиридильных заместителей на термодинамику связывания порфирина с G-квадруплексом ДНК». Биоорганическая и медицинская химия. 21 (23): 7515–22. Дои:10.1016 / j.bmc.2013.09.036. PMID 24148836.
внешняя ссылка
- Группа Nanopore и Aptamer Biosensor {Группа NAB}
Квадруплексные сайты
- G-Quadruplex Мир - сайт для обсуждения публикаций и другой информации, интересной для тех, кто работает в области G-квадруплексов
- Греглист - база данных, в которой перечислены возможные гены, регулируемые G-квадруплексом
- База данных по информации Quadruplex: QuadBase от IGIB
- GRSDB - база данных G-квадруплексов вблизи сайтов процессинга РНК.
- GRS_UTRdb - база данных G-квадруплексов в UTR.
- Сайт ресурсов G-quadruplex
- Инструмент поиска не-B-мотивов в не-B-БД - веб-сервер для прогнозирования мотивов, образующих G-квадруплекс, и других мотивов, не относящихся к B-ДНК, на основе последовательностей ДНК пользователей.
Инструменты для предсказания мотивов G-квадруплекса
- QGRS Mapper: веб-приложение для прогнозирования G-квадруплексов в нуклеотидных последовательностях и генах NCBI из группы Багги.
- Quadfinder: инструмент для прогнозирования и анализа мотивов G-квадруплекса в последовательностях ДНК / РНК от группы Маити, IGIB, Дели, Индия[постоянная мертвая ссылка ]
- [1] G4Hunter из группы Мергни, но пользователю необходимо запустить код на R.
- [2] pqsfinder: исчерпывающий и устойчивый к несовершенству инструмент поиска потенциальных последовательностей, образующих квадруплекс в R.
- [3] pqsfinder: инструмент онлайн-поиска с использованием последней версии пакета R / Bioconductor