Перекрывающийся ген - Overlapping gene
An перекрывающийся ген (или же OLG)[1] это ген чьи выразимые нуклеотидная последовательность частично перекрывается с экспрессируемой нуклеотидной последовательностью другого гена.[2] Таким образом, нуклеотидная последовательность может вносить вклад в функцию одного или нескольких генные продукты. Надпечатка относится к типу перекрытия, при котором вся или часть последовательности одного гена читается в альтернативном рамка чтения из другого гена одновременно локус. Надпечатка была выдвинута как механизм de novo появление новых генов из существующих последовательностей, либо более старых генов, либо ранее некодирование области генома.[3] Гены с надпечаткой - особенно общие черты геномный организация вирусов, вероятно, значительно увеличит количество потенциально экспрессируемых генов из небольшого набора вирусной генетической информации.
Классификация
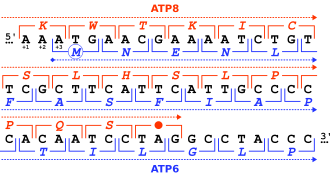
Гены могут перекрываться множеством способов и могут быть классифицированы по их положению относительно друг друга.[2][5][6][7][8]
- Однонаправленный или же тандем перекрытие: 3' конец одного гена перекрывается с 5' конец другого гена на той же цепи. Это расположение может быть обозначено обозначением → → где стрелки указывают рамку считывания от начала до конца.
- Сходящийся или же заканчивается в перекрытие: 3' концы двух генов перекрываются на противоположных цепях. Это можно записать как → ←.
- Расходящийся или же хвост на перекрытие: 5' концы двух генов перекрываются на противоположных цепях. Это можно записать как ← →.
Перекрывающиеся гены также можно классифицировать по фазы, которые описывают их относительные рамки чтения:[2][5][6][7][8]
- Синфазное перекрытие происходит, когда общие последовательности используют одну и ту же рамку считывания. Это также известно как «фаза 0». Однонаправленные гены с перекрытием фазы 0 не считаются отдельными генами, а скорее как альтернативные стартовые площадки того же гена.
- Противофазные перекрытия происходит, когда общие последовательности используют разные рамки считывания. Это может происходить в «фазе 1» или «фазе 2», в зависимости от того, смещены ли рамки считывания на 1 или 2 нуклеотида. Потому что кодон имеет длину три нуклеотида, смещение в три нуклеотида является синфазной рамкой фазы 0.
Эволюция
Перекрывающиеся гены особенно часто встречаются в быстро развивающихся геномах, таких как геномы вирусы, бактерии, и митохондрии. Они могут возникать тремя способами:[9]
- Путем расширения существующего открытая рамка чтения (ORF) ниже по течению в смежный ген из-за потери стоп-кодон;
- Путем расширения существующей открытой рамки считывания до смежного гена из-за потери кодон инициации;
- Путем создания новой ORF внутри существующей за счет точечная мутация.
Использование одной и той же нуклеотидной последовательности для кодирования нескольких генов может обеспечить: эволюционный преимущество за счет сокращения геном размер и возможность транскрипционный и переводной совместное регулирование перекрывающихся генов.[6][10][11][12] Перекрытия генов вводят новые эволюционные ограничения на последовательности перекрывающихся областей.[8][13]
Истоки новых генов

В 1977 г. Пьер-Поль Грассе предположил, что один из генов в паре мог возникнуть de novo мутациями для введения новых ORF в альтернативные рамки считывания; он описал механизм как надпечатка.[15]:231 Позже это было подтверждено Сусуму Оно, который идентифицировал ген-кандидат, который мог возникнуть в результате этого механизма.[16] Некоторые гены de novo, возникающие таким образом, могут не перекрываться, но субфункционализировать следующий дупликация гена[3], способствуя распространению сиротские гены. Можно определить, какой член перекрывающейся пары генов младше биоинформатически либо более ограниченным филогенетический распространение, или менее оптимизированным использование кодонов.[14][17][18] Младшие члены пары, как правило, имеют более высокие внутреннее структурное расстройство чем более старые члены, но и более старые члены также более неупорядочены, чем другие белки, по-видимому, как способ облегчить возросшие эволюционные ограничения, создаваемые перекрытием.[17] Перекрытия чаще возникают в белках, которые уже имеют высокий уровень нарушения.[17]
Таксономическое распределение

Перекрывающиеся гены встречаются во всех области жизни, правда, с разными частотами. Они особенно распространены в популярный геномы.
Вирусы

Существование перекрывающихся генов было впервые обнаружено у вирусов; первый когда-либо секвенированный ДНК-геном бактериофаг ΦX174, содержит несколько примеров.[19] Еще один пример OLG - открытие в октябре 2020 г. ORF3d ген найден в Вирус COVID-19 геном это может повлиять на Пандемия SARS-CoV-2.[1][21] Перекрывающиеся гены особенно часто встречаются у популярный геномы.[14] Некоторые исследования связывают это наблюдение с селективное давление к небольшим размерам генома, опосредованным физическими ограничениями упаковки генома в вирусный капсид, особенно один из икосаэдр геометрия.[22] Однако другие исследования оспаривают этот вывод и утверждают, что распределение перекрытий в вирусных геномах с большей вероятностью отражает наложение как эволюционное происхождение перекрывающихся вирусных генов.[23] Печать поверх - распространенный источник de novo гены в вирусах.[18]
Исследования вирусных генов с надпечаткой показывают, что их белковые продукты, как правило, являются дополнительными белками, которые не являются существенный к распространению вирусов, но способствуют патогенность. Белки с наложенной печатью часто имеют необычные аминокислота распределений и высоких уровней внутренних беспорядок.[24] В некоторых случаях белки с надпечаткой действительно имеют четко определенные, но новые трехмерные структуры;[25] одним из примеров является Супрессор молчания РНК p19 нашел в Томбусвирусы, в котором есть как роман белковая складка и новый режим привязки в распознавании миРНК.[18][20][26]
Прокариоты
Оценки перекрытия генов в бактериальный геномы обычно обнаруживают, что около одной трети бактериальных генов перекрываются, хотя обычно только несколькими парами оснований.[6][27][28] Большинство исследований перекрытия бактериальных геномов обнаруживают доказательства того, что перекрытие выполняет функцию в генная регуляция, позволяя перекрывающимся генам быть транскрипционно и переводно совместно регулируется.[6][12] В прокариотических геномах наиболее распространены однонаправленные перекрытия, возможно, из-за тенденции соседних прокариотических генов к общей ориентации.[6][8][5] Среди однонаправленных перекрытий длинные перекрытия чаще считываются со смещением на один нуклеотид в рамке считывания (т.е. фаза 1), а короткие перекрытия чаще считываются в фазе 2.[28][29] Длинные перекрытия более 60 пар оснований чаще встречаются для конвергентных генов; однако предполагаемое длительное перекрытие имеет очень высокие показатели неправильная аннотация.[30] Надежно подтвержденные примеры длинных перекрытий в бактериальных геномах редки; в хорошо изученных модельный организм кишечная палочка только четыре пары генов хорошо подтверждены как имеющие длинные перекрытия с наложением.[31]
Эукариоты
По сравнению с прокариотическими геномами, эукариотические геномы часто плохо аннотированы, и поэтому идентификация подлинных перекрытий является относительно сложной задачей.[18] Однако примеры подтвержденного перекрытия генов были задокументированы у множества эукариотических организмов, включая млекопитающих, таких как мыши и люди.[32][33][34][35] Эукариоты отличаются от прокариот распределением типов перекрытия: в то время как однонаправленные (т. Е. Одинаковые нити) перекрытия наиболее распространены у прокариот, перекрытия противоположных или антипараллельных цепей чаще встречаются у эукариот. Среди перекрытий противоположных прядей наиболее распространена сходящаяся ориентация.[33] Большинство исследований перекрытия эукариотических генов показало, что перекрывающиеся гены в значительной степени подвержены геномной реорганизации даже у близкородственных видов, и, таким образом, перекрытие не всегда хорошо сохраняется.[34][36] Перекрытие со старыми или менее таксономически ограниченными генами также является общей чертой генов, которые, вероятно, возникли. de novo в данной эукариотической линии.[34][37][38]
Рекомендации
- ^ а б Нельсон, Чейз В.; и другие. (1 октября 2020 г.). «Динамично развивающийся новый перекрывающийся ген как фактор пандемии SARS-CoV-2». eLife. 9. Дои:10.7554 / eLife.59633. ЧВК 7655111. PMID 33001029. Получено 11 ноября 2020.
- ^ а б c Ю. Фукуда, М. Томита и Т. Вашио (1999). «Сравнительное исследование перекрывающихся генов в геномах Mycoplasma genitalium и Mycoplasma pneumoniae". Нуклеиновые кислоты Res. 27 (8): 1847–1853. Дои:10.1093 / nar / 27.8.1847. ЧВК 148392. PMID 10101192.
- ^ а б Киз, ПК; Гиббс, А (15 октября 1992 г.). «Истоки генов:« большой взрыв »или непрерывное творчество?». Труды Национальной академии наук Соединенных Штатов Америки. 89 (20): 9489–93. Bibcode:1992PNAS ... 89.9489K. Дои:10.1073 / пнас.89.20.9489. ЧВК 50157. PMID 1329098.
- ^ Андерсон С., Банкир А.Т., Баррелл Б.Г., де Брюин М.Х., Колсон А.Р., Друин Дж., Эперон И.К., Нирлих Д.П., Роу Б.А., Сэнгер Ф., Шрайер П.Х., Смит А.Дж., Стаден Р., Янг И.Г. (апрель 1981 г.) «Последовательность и организация митохондриального генома человека». Природа. 290 (5806): 457–465. Bibcode:1981Натура.290..457A. Дои:10.1038 / 290457a0. PMID 7219534. S2CID 4355527.
- ^ а б c Фукуда, Йоко; Накаяма, Йоичи; Томита, Масару (декабрь 2003 г.). «О динамике перекрывающихся генов в бактериальных геномах». Ген. 323: 181–187. Дои:10.1016 / j.gene.2003.09.021. PMID 14659892.
- ^ а б c d е ж Джонсон З., Чисхолм С (2004). «Свойства перекрывающихся генов сохраняются в микробных геномах». Genome Res. 14 (11): 2268–72. Дои:10.1101 / гр.2433104. ЧВК 525685. PMID 15520290.
- ^ а б Normark S .; Bergstrom S .; Эдлунд Т .; Grundstrom T .; Jaurin B .; Линдберг Ф.П .; Ольссон О. (1983). «Перекрывающиеся гены». Ежегодный обзор генетики. 17: 499–525. Дои:10.1146 / annurev.ge.17.120183.002435. PMID 6198955.
- ^ а б c d Рогозин, Игорь Б .; Спиридонов, Алексей Н .; Сорокин, Александр В .; Волк, Юрий I .; Джордан, И. Кинг; Татусов, Роман Л .; Кунин, Евгений В. (май 2002 г.). «Очищающий и направленный отбор в перекрывающихся генах прокариот». Тенденции в генетике. 18 (5): 228–232. Дои:10.1016 / S0168-9525 (02) 02649-5. PMID 12047938.
- ^ Кракауэр, Дэвид К. (июнь 2000 г.). «Стабильность и эволюция перекрывающихся генов». Эволюция. 54 (3): 731–739. Дои:10.1111 / j.0014-3820.2000.tb00075.x. PMID 10937248. S2CID 8818055.
- ^ Делай, Луис; ДеЛуна, Александр; Ласкано, Антонио; Бесерра, Артуро (2008). «Происхождение нового гена путем наложения на Escherichia coli». BMC Эволюционная биология. 8 (1): 31. Дои:10.1186/1471-2148-8-31. ЧВК 2268670. PMID 18226237.
- ^ Саха, Дейя; Поддер, Сумита; Панда, Аруп; Гош, Тапаш Чандра (май 2016 г.). «Перекрывающиеся гены: значительный геномный коррелят скорости роста прокариот». Ген. 582 (2): 143–147. Дои:10.1016 / j.gene.2016.02.002. PMID 26853049.
- ^ а б Ло, Инцинь; Баттистуцци, Фабия; Лин, Куи; Гибас, Синтия (29 ноября 2013 г.). «Эволюционная динамика перекрывающихся генов у сальмонелл». PLOS ONE. 8 (11): e81016. Дои:10.1371 / journal.pone.0081016. ЧВК 3843671. PMID 24312259.
- ^ Wei, X .; Чжан, Дж. (31 декабря 2014 г.). «Простой метод оценки силы естественного отбора по перекрывающимся генам». Геномная биология и эволюция. 7 (1): 381–390. Дои:10.1093 / gbe / evu294. ЧВК 4316641. PMID 25552532.
- ^ а б c Павеси, Анджело; Магиоркинис, Гкикас; Карлин, Дэвид Г .; Вилке, Клаус О. (15 августа 2013 г.). «Вирусные белки, полученные De Novo путем наложения печати, могут быть идентифицированы по использованию кодонов: применение в« генетическом питомнике »дельтаретровирусов». PLOS вычислительная биология. 9 (8): e1003162. Дои:10.1371 / journal.pcbi.1003162. ЧВК 3744397. PMID 23966842.
- ^ Грассе, Пьер-Поль (1977). Эволюция живых организмов: доказательства новой теории трансформации. Академическая пресса. ISBN 9781483274096.
- ^ Оно, С. (апрель 1984 г.). «Рождение уникального фермента из альтернативной рамки считывания ранее существовавшей, внутренне повторяющейся кодирующей последовательности». Труды Национальной академии наук Соединенных Штатов Америки. 81 (8): 2421–5. Bibcode:1984PNAS ... 81.2421O. Дои:10.1073 / пнас.81.8.2421. ЧВК 345072. PMID 6585807.
- ^ а б c Уиллис, Сара; Масел, Джоанна (19 июля 2018 г.). «Рождение гена способствует структурному нарушению, кодируемому перекрывающимися генами». Генетика. 210 (1): 303–313. Дои:10.1534 / генетика.118.301249. ЧВК 6116962. PMID 30026186.
- ^ а б c d Sabath, N .; Вагнер, А .; Карлин, Д. (19 июля 2012 г.). "Эволюция вирусных белков, созданных De Novo путем наложения печати". Молекулярная биология и эволюция. 29 (12): 3767–3780. Дои:10.1093 / molbev / mss179. ЧВК 3494269. PMID 22821011.
- ^ а б Sanger, F .; Воздух, Г. М .; Barrell, B.G .; Brown, N.L .; Coulson, A.R .; Fiddes, J.C .; Hutchison, C.A .; Slocombe, P.M .; Смит, М. (1977). «Нуклеотидная последовательность ДНК бактериофага ΦX174». Природа. 265 (5596): 687–95. Bibcode:1977Натура.265..687С. Дои:10.1038 / 265687a0. PMID 870828. S2CID 4206886.
- ^ а б Е, Кэцюн; Малинина, Люси; Патель, Диншоу Дж. (3 декабря 2003 г.). «Распознавание малой интерферирующей РНК вирусным супрессором молчания РНК». Природа. 426 (6968): 874–878. Bibcode:2003Натура.426..874л. Дои:10.1038 / природа02213. ЧВК 4694583. PMID 14661029.
- ^ Докрил, Питер (11 ноября 2020 г.). «Ученые только что обнаружили загадочно спрятанный« ген в гене »SARS-CoV-2». ScienceAlert. Получено 11 ноября 2020.
- ^ Chirico, N .; Вианелли, А .; Белшоу, Р. (7 июля 2010 г.). «Почему у вирусов перекрываются гены». Труды Королевского общества B: биологические науки. 277 (1701): 3809–3817. Дои:10.1098 / rspb.2010.1052. ЧВК 2992710. PMID 20610432.
- ^ Брандес, Надав; Линиал, Михал (21 мая 2016 г.). «Перекрытие генов и ограничения размера в вирусном мире». Биология Директ. 11 (1): 26. Дои:10.1186 / s13062-016-0128-3. ЧВК 4875738. PMID 27209091.
- ^ Rancurel, C .; Хосрави, М .; Дункер, А. К .; Romero, P.R .; Карлин, Д. (29 июля 2009 г.). «Перекрывающиеся гены производят белки с необычными свойствами последовательности и дают представление о создании белков De Novo». Журнал вирусологии. 83 (20): 10719–10736. Дои:10.1128 / JVI.00595-09. ЧВК 2753099. PMID 19640978.
- ^ Аброй, Ааре (1 декабря 2015 г.). «Основанный на белковом домене взгляд на отношения виросферы и хозяина». Биохимия. 119: 231–243. Дои:10.1016 / j.biochi.2015.08.008. PMID 26296474.
- ^ Варгасон, Джеффри М; Ситтья, Дьёрдь; Бургьян, Йожеф; Холл, Трэйси М. Танака (декабрь 2003 г.). «Выборочное распознавание миРНК подавителем молчания РНК». Клетка. 115 (7): 799–811. Дои:10.1016 / S0092-8674 (03) 00984-X. PMID 14697199. S2CID 12993441.
- ^ Хювет, Максим; Штумпф, Майкл PH (1 января 2014 г.). «Перекрывающиеся гены: окно эволюции генов». BMC Genomics. 15 (1): 721. Дои:10.1186/1471-2164-15-721. ISSN 1471-2164. ЧВК 4161906. PMID 25159814.
- ^ а б Кок, Питер Дж. А .; Уитворт, Дэвид Э. (19 марта 2007 г.). «Эволюция перекрытий генов: относительное смещение рамки считывания в генах двухкомпонентной системы прокариот». Журнал молекулярной эволюции. 64 (4): 457–462. Дои:10.1007 / s00239-006-0180-1. PMID 17479344. S2CID 21612308.
- ^ Fonseca, M. M .; Харрис, Д. Дж .; Посада, Д. (5 ноября 2013 г.). «Происхождение и распределение по длине однонаправленных генов перекрытия прокариот». G3: гены, геномы, генетика. 4 (1): 19–27. Дои:10.1534 / g3.113.005652. ЧВК 3887535. PMID 24192837.
- ^ Палледжа, Альберт; Харрингтон, Иоган Д.; Борк, Пер (2008). «Большие перекрытия генов в геномах прокариот: результат функциональных ограничений или ошибочных прогнозов?». BMC Genomics. 9 (1): 335. Дои:10.1186/1471-2164-9-335. ЧВК 2478687. PMID 18627618.
- ^ Фелльнер, Леа; Симон, Свенья; Шерлинг, Кристиан; Уиттинг, Майкл; Шобер, Штеффен; Полте, Кристина; Шмитт-Копплин, Филипп; Keim, Daniel A .; Шерер, Зигфрид; Нойгауз, Клаус (18 декабря 2015 г.). «Доказательства недавнего происхождения бактериального белка, кодирующего перекрывающийся орфанный ген, путем эволюционного наложения». BMC Эволюционная биология. 15 (1): 283. Дои:10.1186 / s12862-015-0558-z. ЧВК 4683798. PMID 26677845.
- ^ McLysaght, Aoife; Герцони, Даниэле (31 августа 2015 г.). «Новые гены из некодирующей последовательности: роль генов de novo, кодирующих белок, в эволюционных инновациях эукариот». Философские труды Королевского общества B: биологические науки. 370 (1678): 20140332. Дои:10.1098 / rstb.2014.0332. ЧВК 4571571. PMID 26323763.
- ^ а б К. Санна, В. Ли и Л. Чжан (2008). «Перекрывающиеся гены в геномах человека и мыши». BMC Genomics. 9 (169): 169. Дои:10.1186/1471-2164-9-169. ЧВК 2335118. PMID 18410680.
- ^ а б c Макаловска, Изабела; Линь Цзяо-Фэн; Эрнандес, Криситина (2007). «Рождение и смерть перекрытия генов у позвоночных». BMC Эволюционная биология. 7 (1): 193. Дои:10.1186/1471-2148-7-193. ЧВК 2151771. PMID 17939861.
- ^ Веерамачанени, В. (1 февраля 2004 г.). «Перекрывающиеся гены млекопитающих: сравнительная перспектива». Геномные исследования. 14 (2): 280–286. Дои:10.1101 / гр.1590904. ЧВК 327103. PMID 14762064.
- ^ Бехура, Сусанта К.; Северсон, Дэвид В. (2013). «Перекрывающиеся гены Aedes aegypti: эволюционные последствия по сравнению с ортологами Anopheles gambiae и других насекомых». BMC Эволюционная биология. 13 (1): 124. Дои:10.1186/1471-2148-13-124. ЧВК 3689595. PMID 23777277.
- ^ Мерфи, Дэниел Н .; McLysaght, Aoife; Кармель, Лиран (21 ноября 2012 г.). "Происхождение генов, кодирующих белок, у мышиных грызунов De Novo". PLOS ONE. 7 (11): e48650. Bibcode:2012PLoSO ... 748650M. Дои:10.1371 / journal.pone.0048650. ЧВК 3504067. PMID 23185269.
- ^ Ноулз, Д. Г .; McLysaght, A. (2 сентября 2009 г.). «Недавнее происхождение de novo генов, кодирующих белок человека». Геномные исследования. 19 (10): 1752–1759. Дои:10.1101 / гр.095026.109. ЧВК 2765279. PMID 19726446.