Технология сайт-специфической рекомбиназы - Site-specific recombinase technology
эта статья может быть слишком техническим для большинства читателей, чтобы понять. Пожалуйста помогите улучшить это к Сделайте это понятным для неспециалистов, не снимая технических деталей. (Май 2019) (Узнайте, как и когда удалить этот шаблон сообщения) |
Сайт-специфические рекомбиназные технологии находятся геномная инженерия инструменты, которые зависят от ферменты рекомбиназы для замены целевых участков ДНК.
История
В конце 1980-х гг. Нацеливание генов у мышей эмбриональные стволовые клетки (ESC) сделали возможным передачу мутаций в зародышевую линию мыши и стали новым вариантом для изучения генетической основы регуляторных сетей, существующих в геноме. Тем не менее, классический нацеливание на гены оказался ограниченным несколькими способами, поскольку функции гена были необратимо разрушены маркером, который должен был быть введен для отбора рекомбинантных ESC. Эти первые шаги привели к появлению животных, у которых мутация присутствовала во всех клетках тела с самого начала, что привело к сложным фенотипам и / или ранней летальности. Существовала явная потребность в методах ограничения этих мутаций определенными точками развития и конкретными типами клеток. Эта мечта стала реальностью, когда группы в США смогли ввести бактериофаги и дрожжевые системы сайт-специфической рекомбинации (SSR-) в клетки млекопитающих, а также в мыши.[1][2][3]
Классификация, свойства и специальные приложения
Общие стратегии генной инженерии требуют постоянной модификации целевого генома. Для этого необходимо приложить немало усилий для разработки маршрутов доставки трансгенов. Хотя для биотехнологических целей случайная интеграция все еще является обычным явлением, она может привести к непредсказуемой экспрессии генов из-за переменного числа копий трансгена, отсутствия контроля над сайтами интеграции и связанных мутаций. Молекулярные требования в области стволовых клеток намного строже. Вот, гомологичная рекомбинация (HR) может, в принципе, обеспечивать специфичность процесса интеграции, но для эукариот это скомпрометировано чрезвычайно низкой эффективностью. Хотя мегануклеазы, эффекторные нуклеазы типа цинковых пальцев и активаторов транскрипции (ZFN и TALEN) являются реальными инструментами, поддерживающими HR, именно доступность сайт-специфичных рекомбиназ (SSR) запустила рациональное построение клеточных линий с предсказуемыми свойствами. В настоящее время обе технологии, HR и SSR, могут быть объединены в высокоэффективные «технологии tag-and-exchange».[4]
Много сайт-специфическая рекомбинация системы были идентифицированы для выполнения этих перестроек ДНК для различных целей, но почти все они принадлежат к одному из двух семейств, тирозин-рекомбиназ (YR) и сериновых рекомбиназ (SR), в зависимости от их механизм. Эти два семейства могут опосредовать до трех типов перестроек ДНК (интеграция, вырезание / разрешение и инверсия) по различным маршрутам реакции в зависимости от их происхождения и архитектуры.[5]
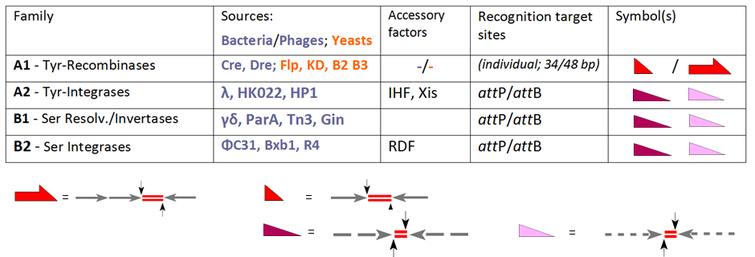
Член-основатель семьи YR - лямбда-интеграза, закодировано бактериофаг λ, позволяя интегрировать ДНК фага в бактериальный геном. Общей чертой этого класса является консервативный нуклеофил тирозина, атакующий расщепляющийся ДНК-фосфат с образованием 3'-фосфотирозиновой связи. Ранние члены семейства SR тесно связаны резольваза / ДНК-инвертазы из бактериальных транспозонов Tn3 и γδ, которые полагаются на каталитический серин, ответственный за атаку ножничного фосфата с образованием 5'-фосфосериновой связи. Эти неоспоримые факты, однако, были скомпрометированы из-за большой путаницы, когда на сцену вышли другие члены, например рекомбиназы YR. Cre и Flp (способные к интеграции, вырезанию / разрешению, а также к инверсии), которые, тем не менее, приветствовались как новые члены «семейства интегразы». Обратными примерами являются PhiC31 и родственные им SR, которые первоначально были введены как резольваза / инвертазы, хотя при отсутствии вспомогательных факторов их единственной функцией является интеграция. В настоящее время стандартная активность каждого фермента определяет его классификацию, оставляя за собой общий термин «рекомбиназа» для членов семейства, которые, по сути, включают все три пути, INT, RES и INV:
Наша таблица расширяет выбор обычных систем SSR и группирует их в соответствии с их характеристиками. Все эти ферменты рекомбинируют два сайта-мишени, которые либо идентичны (подсемейство A1), либо различаются (ферменты фагового происхождения в A2, B1 и B2).[6] Тогда как для A1 эти сайты имеют индивидуальные обозначения ("FRT"в случае Flp-рекомбиназы, loxP для Cre-рекомбиназы), термины "attП "и"attB "(сайты прикрепления на фаговой и бактериальной части соответственно) действительны в остальных случаях. В случае подсемейства A1 мы имеем дело с короткими (обычно 34 п.н.) сайтами, состоящими из двух (почти) идентичных плеч по 13 п.н. (стрелки), фланкирующие спейсер из 8 п.н. (кроссоверная область, обозначенная дублетами красной линии).[7] Обратите внимание, что для Flp существует альтернативный сайт из 48 пар оснований, доступный с тремя ветвями, каждое из которых вмещает единицу Flp (так называемый «протомер»). attP- и attB-сайты следуют аналогичным архитектурным правилам, но здесь руки показывают лишь частичную идентичность (обозначенную пунктирными линиями) и различаются в обоих случаях. Эти особенности объясняют существенные различия:
- рекомбинация двух идентичных сайтов эдукта приводит к сайтам продуктов с одинаковым составом, хотя они содержат ответвления от обоих субстратов; эти преобразования обратимы;
- в случае attP x attКроссоверы B-рекомбинации могут происходить только между этими дополнительными партнерами в процессах, которые приводят к двум различным продуктам (attP x attB → attR + attL) необратимым образом.
Чтобы упростить эту главу, следующие реализации будут сосредоточены на двух рекомбиназах (Flp и Cre) и только на одной интегразе (PhiC31), поскольку их спектр охватывает инструменты, которые в настоящее время в основном используются для направленных модификаций генома. Это будет сделано в рамках следующего обзора.

Пути реакции
Режим интеграции / разрешения и инверсии (INT / RES и INV) зависит от ориентации сайтов-мишеней рекомбиназы (RTS) среди этих пар attP и attB. В разделе C в упрощенной форме указывается, как рекомбиназа-опосредованная замена кассет (RMCE) может быть достигнуто путем синхронных двойных взаимных кроссоверов (а не интеграции с последующим разрешением).[8][9]
Тир-рекомбиназы обратимы, а сер-интеграза - однонаправленна. Следует отметить, что обратимая интеграция / разрешение Flp (Tyr-рекомбиназы) модулируется 48 п.н. (вместо минимальных 34 п.н.) FRT версии: дополнительное плечо из 13 п.н. служит «траекторией посадки» Flp, способствующей формированию синаптического комплекса, как в контексте функций Flp-INT, так и Flp-RMCE (см. соответствующие ситуации равновесия). Хотя едва ли возможно предотвратить (управляемую энтропией) реверсию интеграции в разделе A для Cre и трудно достичь для Flp, RMCE может быть завершен, если донорская плазмида предоставляется в избытке из-за бимолекулярного характера обеих прямых - и обратная реакция. Позирует как FRT сайтов обратным образом приведет к равновесию обеих ориентаций вставки (зеленая стрелка). В отличие от Flp, Ser-интеграза PhiC31 (нижние изображения) приводит к однонаправленной интеграции, по крайней мере, в отсутствие фактора рекомбиназной направленности (RDF-).[10] По сравнению с Flp-RMCE, который требует двух разных («гетероспецифических») FRT-спейсерные мутанты, партнер реакции (attБ) первой реагирующей attР-сайт поражается произвольно, так что нет возможности контролировать направление, в котором донорская кассета входит в цель (см. Альтернативные продукты). Также отличается от Flp-RMCE, несколько различных мишеней RMCE не могут быть установлены параллельно из-за отсутствия гетероспецифических (не перекрестно взаимодействующих) attП/attКомбинации B.
Cre рекомбиназа
Cre рекомбиназа (Cre) может рекомбинировать определенные последовательности ДНК без необходимости кофакторы. Фермент распознает последовательности ДНК из 34 пар оснований, называемых loxP («локус кроссовера в фаге P1»). В зависимости от ориентации сайтов-мишеней относительно друг друга Cre будет интегрировать / вырезать или инвертировать последовательности ДНК. После иссечения (называемого «расслоением» в случае кольцевого субстрата) определенной области ДНК нормальная экспрессия гена значительно нарушается или прекращается.[11]
Из-за выраженной разрешающей способности Cre одним из первых его применений было удаление лохР-фланкированные («флоксовые») гены, приводящие к нокауту клеточно-специфического гена такого флоксованного гена после того, как Cre становится экспрессированным в интересующей ткани. Современные технологии включают методы, которые позволяют как пространственный, так и временной контроль активности Cre. Общий метод, облегчающий пространственный контроль генетических изменений, включает выбор тканеспецифичного промоутер для управления выражением Cre. Размещение Cre под контролем такого промотора приводит к локальной тканеспецифической экспрессии. Например, Леоне и др. поместили единицу транскрипции под контроль регуляторных последовательностей миелин ген протеолипидного белка (PLP), приводящий к индуцированному удалению целевых последовательностей генов в олигодендроциты и Шванновские клетки.[12] Специфический фрагмент ДНК, распознаваемый Cre, остается интактным в клетках, которые не экспрессируют ген PLP; это, в свою очередь, облегчает эмпирическое наблюдение локальных эффектов изменений генома в миелиновой оболочке, окружающей нервные волокна в Центральная нервная система (ЦНС) и периферическая нервная система (ПНС).[13] Селективная экспрессия Cre была достигнута также во многих других типах клеток и тканях.
Чтобы контролировать временную активность реакции вырезания, формы Cre, которые используют различные лиганд связывающие домены были разработаны. Одна успешная стратегия индукции специфической временной активности Cre включает слияние фермента с мутантным лиганд-связывающим доменом человека. рецептор эстрогена (ERt). После введения тамоксифен (эстроген антагонист рецепторов ), конструкция Cre-ERt способна проникать в ядро и вызывать целевую мутацию. ERt связывает тамоксифен с большей аффинностью, чем эндогенный эстрогены, что позволяет Cre-ERt оставаться цитоплазматический у животных, не получавших тамоксифен. Временной контроль активности SSR тамоксифеном позволяет индуцировать генетические изменения позже в эмбриогенез и / или во взрослых тканях.[12] Это позволяет исследователям избежать эмбриональной летальности, продолжая исследовать функцию целевых генов.
Недавнее расширение этих общих концепций привело к созданию «Cre-zoo», то есть коллекций сотен линий мышей, для которых определенные гены могут быть удалены с помощью целевой экспрессии Cre.[3]

Рекомбиназа flp
У своего естественного хозяина (S. cerevisiae) Flp /FRT система обеспечивает репликацию «плазмиды 2μ» путем инверсии сегмента, который обрамлен двумя идентичными, но противоположно ориентированными FRT сайты (активность "flippase"). Эта инверсия изменяет относительную ориентацию вилок репликации внутри плазмиды, обеспечивая «катящийся круг» - усиление кольцевой 2-микронной сущности до того, как мультимерные промежуточные соединения разделятся с высвобождением множественных мономерных продуктов. В то время как минимум 34 п.н. FRT сайты поддерживают удаление / разрешение в той же степени, что и аналог лохP-сайты для Cre, естественных, более протяженных вариантов FRT размером 48 п.н., обеспечивают более высокую степень интеграции, преодолевая при этом определенные беспорядочные взаимодействия, как описано для фаговых ферментов, таких как Cre- [5] и PhiC31.[6] Дополнительным преимуществом является то, что можно применять простые правила для создания гетероспецифических FRT сайты, которые пересекаются с равными партнерами, но не с диким типом FRTс. Эти факты позволили с 1994 года развивать и непрерывно совершенствовать рекомбиназа-опосредованная замена кассет (RMCE-) стратегии, позволяющие чистую замену целевой кассеты на поступающую донорскую кассету.[6]
Основанный на технологии RMCE, в рамках программы EUCOMM (Европейский условный мутагенез мышей), основанной на уже существующих Cre- и / или Flp-, был разработан конкретный ресурс предварительно охарактеризованных штаммов ES, который поддается дальнейшей разработке. установки на основе "FlExing" (Flp-опосредованное иссечение / инверсия),[6] с участием эксцизии и инверсии. Этот проект, инициированный в 2005 году, в первую очередь был сфокусирован на мутагенезе насыщения, чтобы обеспечить полную функциональную аннотацию генома мыши (координируемый Международным консорциумом нокаут-мышей, IKMC) с конечной целью, чтобы все гены белка были мутированы посредством улавливания и нацеливания генов у мышей. ES клетки.[14] Эти усилия знаменуют собой вершину различных стратегий «метки и обмена», которые посвящены метке отдельного геномного сайта, так что «метка» может служить адресом для введения новой (или изменения существующей) генетической информации. Шаг пометки как таковой могут обращаться к определенным классам сайтов интеграции, используя предпочтения интеграции ретровирусов или даже интеграций, специфичных для сайта, таких как PhiC31, оба из которых действуют по существу однонаправленным образом.
Традиционные трудоемкие процедуры «метки и обмена» основывались на двух последовательных стадиях гомологичной рекомбинации (HR-), первая из которых («HR1») предназначена для введения метки, состоящей из гена маркера отбора. Затем для замены маркера на «GOI» использовали «HR2». В первой реакции («нокаут» -) ген был помечен селектируемым маркером, обычно путем вставки кассеты hygtk ([+/-]) обеспечение устойчивости к G418. На следующем этапе «нокаута» помеченная геномная последовательность была заменена гомологичными геномными последовательностями с определенными мутациями. Затем можно выделить клоны клеток по их устойчивости к ганцикловиру из-за потери гена HSV-tk, т.е. («отрицательный выбор»). Эта обычная двухэтапная процедура «теги и обмен» [15] можно было бы оптимизировать после появления RMCE, который мог бы взять на себя и повысить эффективность этапа врезки.
PhiC31 интеграза
Без особых сомнений, Сер интегрирует являются текущими инструментами выбора для интеграции трансгенов в ограниченное число хорошо изученных геномных акцепторных сайтов, которые в основном (но не всегда) имитируют фаг attP в том, что они привлекают attB-содержащий донорный вектор. В настоящее время наиболее заметным членом является PhiC31-INT с доказанным потенциалом в контексте геномов человека и мыши.
В отличие от вышеупомянутых рекомбиназ Tyr, PhiC31-INT как таковой действует однонаправленно, прочно фиксируя донорный вектор на геномно заякоренной мишени. Очевидным преимуществом этой системы является то, что она может полагаться на неизмененные, собственные attP (акцептор) и attB донорские сайты. Дополнительные преимущества (вместе с некоторыми осложнениями) могут возникнуть из-за того, что геномы мыши и человека сами по себе содержат ограниченное количество эндогенных мишеней (так называемые "attP-псевдозиты "). Имеющаяся информация свидетельствует о том, что значительные требования к последовательности ДНК позволяют интегразе распознавать меньше сайтов, чем системы интеграции на основе ретровирусов или даже транспозаз, открывая свою карьеру в качестве превосходного носителя для транспорта и вставки в ряд хорошо установленных геномных сайтов , некоторые из которых обладают так называемыми «безопасными» свойствами.[10]
Используя факт конкретного (attP x attБ) пути рекомбинации, RMCE становится возможным без потребности в синтетических, гетероспецифических att-места. Однако это очевидное преимущество достигается за счет определенных недостатков, таких как отсутствие контроля над видом или направленностью входящей (донорной) кассеты.[6] Дальнейшие ограничения наложены тем фактом, что необратимость не допускает стандартных мультиплексирование-RMCE установки, включающие "последовательные реакции RMCE", то есть повторные замены кассет в данной геномной локус.
Перспективы и перспективы
Аннотации геномов человека и мыши привели к идентификации> 20 000 генов, кодирующих белок, и> 3 000 генов некодирующих РНК, которые направляют развитие организма от оплодотворения через эмбриогенез до взрослой жизни. Хотя отмечен значительный прогресс, актуальность редких вариантов генов остается центральной темой исследований.
В качестве одной из наиболее важных платформ для работы с функциями генов позвоночных в крупном масштабе были созданы полногеномные генетические ресурсы мутантных мышиных ES-клеток. С этой целью в Европе и Северной Америке были созданы четыре международные программы, направленные на насыщающий мутагенез генома мыши (EUCOMM, KOMP, NorCOMM и TIGM). Эти репозитории ES-клеток, координируемые Международным консорциумом мышей-нокаутов (IKSC), доступны для обмена между международными исследовательскими подразделениями. Настоящие ресурсы включают мутации в 11 539 уникальных генах, 4 414 из которых являются условными.[14]
Соответствующие технологии сейчас достигли уровня, позволяющего распространить их на другие виды млекопитающих и стволовые клетки человека, в первую очередь на те, которые имеют ИПС (индуцированный плюрипотент) статус.
Смотрите также
- Сайт-специфическая рекомбинация
- Обмен кассет, опосредованный рекомбиназой
- Cre рекомбиназа
- Cre-Lox рекомбинация
- Рекомбинация FLP-FRT
- Генетическая рекомбинация
- Гомологичная рекомбинация
использованная литература
- ^ Зауэр, Брайан; Хендерсон, Нэнси (1988). «Сайт-специфическая рекомбинация ДНК в клетках млекопитающих с помощью Cre-рекомбиназы бактериофага P1». Труды Национальной академии наук Соединенных Штатов Америки. 85 (14): 5166–70. Bibcode:1988ПНАС ... 85.5166С. Дои:10.1073 / pnas.85.14.5166. JSTOR 32380. ЧВК 281709. PMID 2839833.
- ^ О'Горман, Стивен; Фокс, Дэниел Т .; Уол, Джеффри М. (1991). «Активация гена, опосредованная рекомбиназой, и сайт-специфическая интеграция в клетках млекопитающих». Наука. 251 (4999): 1351–5. Bibcode:1991Научный ... 251.1351O. Дои:10.1126 / science.1900642. JSTOR 2875533. PMID 1900642.
- ^ а б Раевский, Клаус (2007). «От мечты к реальности». Европейский журнал иммунологии. 37: S134–7. Дои:10.1002 / eji.200737819. PMID 17972357.
- ^ Бранда, Кэтрин С .; Димеки, Сьюзен М. (2004). «Говоря о революции. Влияние сайт-специфичных рекомбиназ на генетический анализ мышей». Клетка развития. 6 (1): 7–28. Дои:10.1016 / S1534-5807 (03) 00399-X. PMID 14723844.
- ^ а б Nern, A .; Pfeiffer, B.D .; Свобода, К .; Рубин, Г. М. (2011). «Множество новых сайт-специфичных рекомбиназ для использования при манипулировании геномами животных». Труды Национальной академии наук. 108 (34): 14198–203. Bibcode:2011ПНАС..10814198Н. Дои:10.1073 / pnas.1111704108. ЧВК 3161616. PMID 21831835.
- ^ а б c d е ж Туран, С .; Боде, Дж. (2011). «Сайт-специфичные рекомбиназы: от геномных модификаций на основе меток и мишеней до модификаций на основе меток и обмена». Журнал FASEB. 25 (12): 4088–107. Дои:10.1096 / fj.11-186940. PMID 21891781.
- ^ Боде, Юрген; Шлейк, Томас; Ибер, Микаэла; Шубелер, Дирк; Зайблер, Йост; Снежков, Евгений; Николаев, Лев (2000). «Набор инструментов трансгенетиков: новые методы целевой модификации геномов эукариот». Биологическая химия. 381 (9–10): 801–13. Дои:10.1515 / BC.2000.103. PMID 11076013.
- ^ Lyznik, Leszek A .; Митчелл, Джон С .; Хираяма, Линн; Ходжес, Томас К. (1993). «Активность дрожжевой рекомбиназы FLP в протопластах кукурузы и риса». Исследования нуклеиновых кислот. 21 (4): 969–75. Дои:10.1093 / nar / 21.4.969. ЧВК 309231. PMID 8451196.
- ^ Lauth, M .; Spreafico, F; Dethleffsen, K; Мейер, М (2002). «Стабильный и эффективный обмен кассет в неизбираемых условиях путем комбинированного использования двух сайт-специфичных рекомбиназ». Исследования нуклеиновых кислот. 30 (21): 115e. Дои:10.1093 / нар / gnf114. ЧВК 135837. PMID 12409474.
- ^ а б Каров, Мариса; Калос, Мишель П (2011). «Терапевтический потенциал интегразы phiC31 как системы генной терапии». Мнение эксперта по биологической терапии. 11 (10): 1287–96. Дои:10.1517/14712598.2011.601293. PMID 21736536.
- ^ Умар, Андре; Цяо, Цзюньхуа; Джосток, Томас; Ли, Цзяньдун; Боде, Юрген (2006). «Рекомендуемый метод использования хромосом: системы обмена кассет на основе RMCE в биотехнологии клеток животных». Цитотехнология. 50 (1–3): 93–108. Дои:10.1007 / s10616-006-6550-0. ЧВК 3476001. PMID 19003073.
- ^ а б Леоне, Дино П.; Genoud, S.Téphane; Атанасоски, Сузана; Граузенбургер, Рейнхард; Бергер, Филипп; Мецгер, Даниэль; МакКлин, Венди Б; Шамбон, Пьер; Сутер, Ули (2003). «Тамоксифен-индуцируемые глии-специфичные мыши Cre для соматического мутагенеза в олигодендроцитах и шванновских клетках». Молекулярная и клеточная неврология. 22 (4): 430–40. Дои:10.1016 / S1044-7431 (03) 00029-0. PMID 12727441.
- ^ Koenning, M .; Jackson, S .; Hay, C.M .; Faux, C .; Килпатрик, Т. Дж .; Willingham, M .; Эмери, Б. (2012). «Регуляторный фактор гена миелина необходим для поддержания идентичности миелина и зрелых олигодендроцитов в ЦНС взрослого». Журнал неврологии. 32 (36): 12528–42. Дои:10.1523 / JNEUROSCI.1069-12.2012. ЧВК 3752083. PMID 22956843.
- ^ а б Брэдли, Аллан; Анастасиадис, Константинос; Аяди, Абделькадер; Бэтти, Джеймс Ф .; Белл, Синди; Бирлинг, Мари-Кристин; Боттомли, Джоанна; Браун, Стив Д .; и другие. (2012). "Ресурс функции генов млекопитающих: Международный консорциум нокаут-мышей". Геном млекопитающих. 23 (9–10): 580–6. Дои:10.1007 / s00335-012-9422-2. ЧВК 3463800. PMID 22968824.
- ^ Аскью, Дж. Роджер; Дётчман, Томас; Лингрел, Джерри Б. (1993). «Сайт-ориентированные точечные мутации в эмбриональных стволовых клетках: стратегия меток и обмена генов». Молекулярная и клеточная биология. 13 (7): 4115–24. Дои:10.1128 / MCB.13.7.4115. ЧВК 359961. PMID 8391633.
внешние ссылки
- http://www.knockoutmouse.org/
- Emes, RD; Goodstadt, L; Winter, EE; Понтинг, CP (2003). «Сравнение геномов человека и мыши закладывает основы зоологии генома». Хум Мол Генет. 12 (7): 701–9. Дои:10.1093 / hmg / ddg078. PMID 12651866.