Соматический эмбриогенез - Somatic embryogenesis
Соматический эмбриогенез это искусственный процесс, в котором растение или эмбрион получают из одного Соматическая клетка.[1] Соматические зародыши образуются из растительных клеток, которые обычно не участвуют в развитии зародышей, то есть из обычной растительной ткани. Вокруг соматического зародыша не образуется эндосперм или семенная оболочка.
Клетки, полученные из компетентной исходной ткани, культивируют для образования недифференцированной массы клеток, называемой мозоль. Регуляторы роста растений в среде для культивирования ткани можно манипулировать, чтобы вызвать образование каллуса, а затем изменить, чтобы заставить эмбрионы формировать каллус. Соотношение разных регуляторы роста растений необходимое для образования каллуса или зародыша зависит от типа растения.[2] Соматические эмбрионы в основном производятся in vitro и для лабораторных целей, используя твердые или жидкие питательные среды которые содержат регуляторы роста растений (PGR). Основные используемые ГРР: ауксины но может содержать цитокинин в меньшем количестве.[3] Побеги и корни монополярны, в то время как соматические зародыши биполярны, что позволяет им формировать целое растение без культивирования на нескольких типах сред. Соматический эмбриогенез служил моделью для понимания физиологических и биохимических событий, происходящих во время процессов развития растений, а также одним из компонентов биотехнологического прогресса.[4] Первые документы о соматическом эмбриогенезе были получены Steward et al. в 1958 г. и Рейнерт в 1959 г. с суспензионными культурами клеток моркови.[5][6]
Прямой и непрямой эмбриогенез
Было описано, что соматический эмбриогенез происходит двумя способами: прямо или косвенно.[7]
Прямой эмбриогенез
происходит, когда эмбрионы запускаются непосредственно из ткани эксплантата, создавая идентичный клон.
Непрямой эмбриогенез
происходит, когда эксплантаты продуцируют недифференцированные или частично дифференцированные клетки (часто называемые каллусом), которые затем сохраняются или дифференцируются в ткани растения, такие как лист, стебель или корни. 2,4-дихлорфеноксиуксусная кислота (2,4-D), 6-бензиламинопурин (BAP) и Гибберелловая кислота (GA) был использован для развития непрямых соматических зародышей клубники (Fragaria ananassa) [8]
Регенерация растений соматическим эмбриогенезом
Регенерация растений посредством соматического эмбриогенеза происходит в пять этапов: инициирование эмбриогенных культур, пролиферация эмбриогенных культур, преждевременное созревание соматических зародышей, созревание соматических зародышей и развитие растений на неспецифической среде. Инициация и пролиферация происходят в среде, богатой ауксином, который вызывает дифференцировку. локализованных меристематические клетки. В ауксин обычно используется 2,4-Д. После перехода на средний уровень с низким или нулевым ауксин эти клетки могут затем развиться в зрелые эмбрионы. Прорастание соматического зародыша может произойти только тогда, когда он достаточно зрел, чтобы иметь функциональные возможности. корень и стрелять вершины [3]
Факторы, влияющие
Факторы и механизмы, контролирующие дифференцировку клеток соматических эмбрионов, относительно неоднозначны. Было показано, что некоторые соединения, выделяемые культурами тканей растений и обнаруживаемые в питательной среде, необходимы для координации деления клеток и морфологических изменений.[9] Эти соединения были идентифицированы Chung et al.[10] как различные полисахариды, аминокислоты, регуляторы роста, витамины, низкомолекулярные соединения и полипептиды. Несколько сигнальные молекулы Было обнаружено, что они влияют на формирование соматических эмбрионов или контролируют их, и включают внеклеточные белки, арабиногалактановые белки и липохитоолигосахариды. Температура и освещение также могут влиять на созревание соматического эмбриона.
Приложения
Применения этого процесса включают: клональное размножение из генетически однородного растительного материала; устранение вирусы; предоставление исходной ткани для генетических трансформация; создание целых растений из отдельных клеток, называемых протопласты; разработка технологии синтетических семян.[1]
Использование соматического эмбриогенеза
- Растение трансформации[11]
- Масса распространение[12]
Развитие процедур соматического эмбриогенеза привело к исследованиям запасных белков семян (SSP) древесные растения для древесных пород, имеющих промышленное значение, т.е. в основном голосеменные, включая белая ель. В этой области исследований SSP используются в качестве маркеров для определения эмбриогенного потенциала и способности эмбриогенной системы производить соматический эмбрион, биохимически похожий на его. зиготический аналог (Флинн и др., 1991, Бирдмор и др., 1997).[13][14]
Grossnickle et al. (1992)[15] сравнил интерьерную ель саженцы с посадками во время развития питомника и с помощью программы оценки качества поголовья непосредственно перед высаживанием в поле. Высота всхода рассады, корневая шейка диаметр и сухая масса у проростков увеличивались быстрее, чем у зародышей, в течение первой половины первого вегетационного периода, но после этого рост побегов был одинаковым для всех растений. К концу вегетационного периода проростки были на 70% выше, чем зародыши, имели больший диаметр корневой шейки и большую сухую массу побегов. В начале вегетационного периода у проростков сухой вес корней увеличивался быстрее, чем у прививок.
Во время осенней акклиматизации картина увеличения индекса высвобождения покоя и повышения устойчивости к замораживанию была сходной как у проростков, так и у посадок. При осенней акклиматизации способность к росту корней снижалась, затем увеличивалась, причем у проростков это увеличение было больше.
Оценка качества стада непосредственно перед посадкой показала, что: у посадок было больше эффективности водопользования с уменьшением предрассветного водного потенциала побегов по сравнению с сеянцами; саженцы и зародыши имели одинаковую способность движения воды как при высокой, так и при низкой температуре корней; чистый фотосинтез и проводимость хвои при низких температурах корней были выше у проростков, чем у закваски; и у проростков был больший рост корней, чем у зародышей при температуре корня 22 ° C, но рост корней у всех растений был низким при температуре корня 7,5 ° C.
Рост и выживаемость сеянцев и приростов внутренней ели 313B Styroblock после высадки на участке лесовосстановления определяли Grossnickle and Major (1992).[16] Как для проростков, так и для зародышей, осмотический потенциал при насыщении (ψсидел) и точки потери тургора (ψкончик) увеличилась с минимального значения -1,82 и -2,22 МПа, соответственно, непосредственно перед посадкой до сезонного максимума -1,09 и -1,21 МПа, соответственно, во время активного удлинения побегов. После этого всходы и посадки (ψсидел) и (ψкончик) снизилась до -2,00 и -2,45 МПа соответственно в конце вегетационного периода, что совпало с устойчивым снижением температуры участков и прекращением роста высоты. В целом у проростков и приростов ψсидел и ψкончик значений в течение вегетационного периода, а также имели аналогичные сдвиги в сезонных моделях максимального модуля упругости, симпатической фракции и относительного содержания воды на потеря тургора точка.
Гроссникл и майор (1992)[16] обнаружили, что хвоя однолетних и текущих годов как у сеянцев, так и у привитых растений имела одинаковое снижение проводимости игл с увеличением давление газа дефицит. Модели поверхности отклика нетто-фотосинтеза хвои текущего года (Pп) реакция на дефицит давления пара (VPD) и фотосинтетически активную радиацию (PAR) показали, что у эмблинга на 15% больше Pп при VPD менее 3,0 кПа и PAR более 1000 мкмоль · м−2s−1. Хвоя саженцев и посевов годовалого и текущего года показала схожие модели эффективности использования воды.
Темпы роста побегов у сеянцев и посадок в течение вегетационного периода также были сходными. У сеянцев была более крупная система побегов как во время посадки, так и в конце вегетационного периода. У проростков также было более сильное развитие корней, чем у зародышей в течение вегетационного периода, но соотношение корня: побеги для двух типов подвоя было одинаковым в конце вегетационного периода, когда выживаемость сеянцев и заквасок составляла 96% и 99% соответственно.
Карты слежения и судьбы
Понимание формирования соматического эмбриона путем установления морфологических и молекулярных маркеров важно для построения карты судьбы. Карта судьбы - это фундамент для дальнейших исследований и экспериментов. Существуют два метода построения карты судьбы: синхронное деление клеток и покадровое отслеживание. Последний обычно работает более стабильно из-за химических веществ, изменяющих клеточный цикл, и центрифугирования, участвующих в синхронном делении клеток.[17]
Покрытосеменные
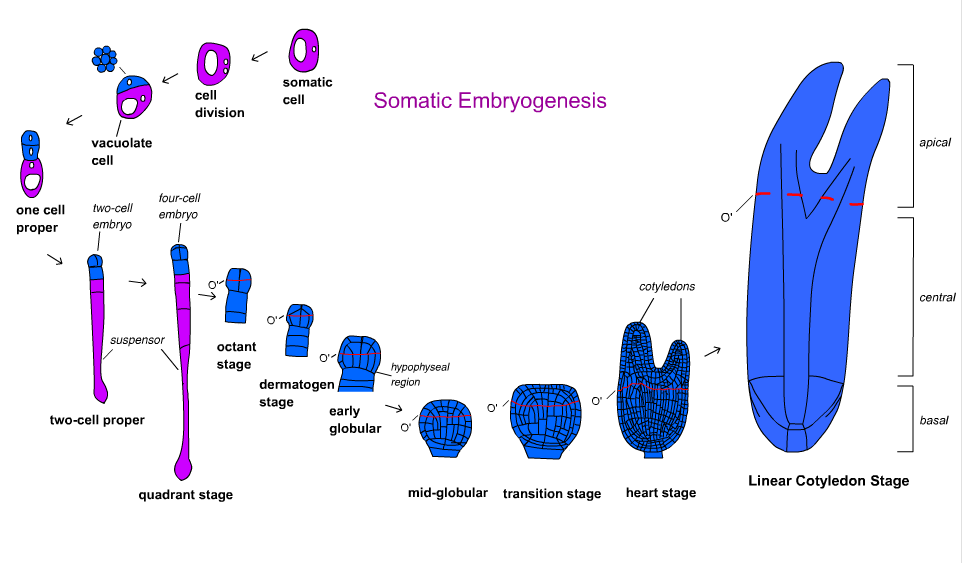
Развитие эмбриона у покрытосеменных растений делится на несколько этапов. Зигота разделена асимметрично, образуя небольшую апикальную клетку и большую базальную клетку. Организационный паттерн формируется на стадии шаровидного тела, а затем эмбрион переходит в стадию семядоли.[18] Развитие зародыша отличается у однодольных и двудольных. Двудольные проходят через шаровидную, сердцевидную и торпедную стадии, а однодольные проходят через шаровидную, щитковидную и жесткоптилярную стадии.[19]
Многие системы культивирования индуцируют и поддерживают соматический эмбриогенез путем непрерывного воздействия 2,4-дихлорфеноксиуксусная кислота. Абсцизовая кислота сообщалось, что он индуцирует соматический эмбриогенез у проростков. После мозоль формирование, культивирование на среде с низким содержанием ауксина или без гормонов будет способствовать соматическому росту зародыша и формированию корней. В однодольные, эмбриогенная способность обычно ограничивается тканями эмбриогенного или меристематического происхождения. Соматические клетки однодольных быстро дифференцируются, а затем теряют митотическую и морфогенную способность. Различия в чувствительности к ауксину при росте эмбриогенного каллуса между разными генотипами одного и того же вида показывают, насколько разнообразными могут быть ответы на ауксин.[20]
Морковь Daucus carota был первым и наиболее изученным видом в отношении путей развития и молекулярных механизмов.[17] Покадровое отслеживание Toonen et al. (1994) показали, что морфология компетентных клеток может варьироваться в зависимости от формы и плотности цитоплазмы. Из эмбриональной суспензии было идентифицировано пять типов клеток: сферические клетки с богатой цитоплазмой, сферические вакуолизированные, овальные вакуолизированные, удлиненные вакуолизированные и клетки неправильной формы. Каждый тип клеток размножается с определенной геометрической симметрией. Они превратились в симметричные, асимметричные кластеры клеток аберрантной формы, которые в конечном итоге сформировали эмбрионы с разной частотой.[21] Это указывает на то, что полярность организованного роста не всегда существует в соматическом эмбриогенезе.[17]
Голосеменные
Развитие эмбриона в голосеменные происходит в три фазы. Проэмбриогенез включает все стадии, предшествующие суспензор удлинение. Ранний эмбриогенез включает все стадии после удлинения суспензора, но до развития корневой меристемы. Поздний эмбриогенез включает развитие меристем корня и побега.[18]Замедленное отслеживание в Ель обыкновенная Picea abies показали, что ни одиночные богатые цитоплазмой клетки, ни вакуолизированные клетки не развиваются в эмбрионы. Проэмбриогенные массы (PEM), промежуточное звено между неорганизованными клетками и эмбрионом, состоящим из богатых цитоплазмой клеток рядом с вакуолизированной клеткой, стимулируются с помощью ауксин и цитокинин. Постепенное удаление ауксина и цитокинина и введение абсцизовая кислота (ABA) позволит сформироваться эмбриону.[17] Использование соматического эмбриогенеза было рассмотрено для массового производства вегетативно размножаемых клонов хвойных деревьев и криоконсервации гермоплазма. Однако использование этой технологии для лесовосстановления и выращивания хвойных деревьев находится в зачаточном состоянии.[22][23]
Смотрите также
- Эмбриогенез растений
- Каллус (клеточная биология)
- Культура тканей растений
- Гормон растения
- Спасение эмбрионов
- Гипергидричность
- Мурашиге и Скуг средний
Рекомендации
- ^ а б Саху, Джоти Пракаш (11.06.2018). «Органогенез и соматический эмбриогенез». Дои:10.13140 / rg.2.2.26278.57928. Цитировать журнал требует
| журнал =(помощь) - ^ http://www.accessexcellence.org/LC/ST/st2bgplant.html Культура тканей растений
- ^ а б E.F. George et al. (ред.), Размножение растений тканевой культурой, 3-е издание, 335-354.
- ^ Кирос-Фигероа, Ф. Р., Рохас-Эррера, Р., Галаз-Авалос, Р. М., и Лойола-Варгас, В. М. 2006. Производство эмбрионов посредством соматического эмбриогенеза можно использовать для изучения дифференцировки клеток у растений. Растительная клеточная ткань. Орг. Культ. 86: 285–301.
- ^ Стюард, Ф. К., Мэйпс, М. О. и Смит, Дж. (1958). Рост и организованное развитие культивируемых клеток. I. Рост и деление свободно подвешенных клеток. Являюсь. J. Bot. 45, 693-703.
- ^ Reinert J (1959) Uber die kontrolle der morphogenese und die индукция от adventivembryonen и gewebekulturen aus karotten. Планта 53: 318–333
- ^ Sharp et al. (1980). В: Обзоры садоводства, Vol. 2. (Джаник, Дж., Ред.). AVI Publishing Co, Вестпорт, Коннектикут, США, стр. 268.
- ^ Вагмаре, С. Г., Павар, К. Р., и Табе, Р. 2017. Соматический эмбриогенез у земляники (Fragaria ananassa) var. Камароза. Глобальный журнал биологии и биотехнологии 6 (2): 309 - 313.
- ^ Уоррен, Г.С., Фаулер, М.В. 1981. Физиологические взаимодействия на начальных этапах эмбриогенеза в культурах Daucus carota L. New Phytol 87: 481-486.
- ^ Чанг, В., Педерсен, Х., Чин, К. К.. 1992. Повышенное производство соматических эмбрионов кондиционированной средой в клеточных суспензионных культурах Daucus carota. Biotechnol Lett 14: 837-840.
- ^ Клэпхэм; и другие. (2000). «Перенос генов путем бомбардировки частицами в эмбриогенные культуры Picea abies и производство трансгенных проростков». Скандинавский журнал исследований леса. 15 (2): 151–160. Дои:10.1080/028275800750014957. S2CID 84397445.
- ^ Хименес В.М., Гевара Э., Эррера Дж. И Бангерт Ф. 2001. Уровни эндогенных гормонов в привыкших к нуцеллару Citrus мозоль на начальных этапах регенерации. Реплика растительных клеток 20: 92–100.
- ^ Flinn, B.S .; Робертс, Д.Р .; Webb, D.T .; Саттон, Британская Колумбия 1991. Изменения запасного белка во время зиготического эмбриогенеза у внутренней ели. Tree Physiol. 8: 71–81. (Цитируется по Beardmore et al. 1997).
- ^ Beardmore, T.L .; Wetzel, S .; Реган, С. 1997. Запасные белки семян тополя. Гл. 17, стр. 131–142 в Klopfenstein, N.B .; Chun, Y.W .; Kim, M.S .; Ахуджа, М.Р. (ред.), Диллон, М.К .; Carman, R.C .; Эскью, Л. (Tech. Eds.) 1997. Микроразмножение, генная инженерия и молекулярная биология Populus. USDA, для. Serv., Rocky Mountain Res. Sta., Fort Collins CO, Gen. Tech. Реп. RM-GTR-297.
- ^ Grossnickle, S.C .; Робертс, Д.Р .; Major, J.E .; Folk, R.S .; Webster, F.B .; Саттон, Б.С.С. 1992. Интеграция соматического эмбриогенеза в оперативное лесоводство: сравнение зародышей и сеянцев ели во время производства 1 + 0 стада. п. 106–113 в Лэндис, Т. Д. (Техническая координация). Proc. Ассоциация питомников межгорных лесов, август 1991 г., Парк-Сити, Юта. USDA, для. Серв., Скалистая гора. За. Диапазон Exp. Sta., Fort Collins CO, Gen. Tech. Реп. RM-211.
- ^ а б Grossnickle, S.C .; Major, J.E. 1992. Внутренние сеянцы ели по сравнению с зародышами, полученными в результате соматического эмбриогенеза. 2. Физиологическая реакция и морфологическое развитие на участке лесовосстановления. п. 98 (сокращ.) в Коломбо, С.Дж .; Hogan, G .; Уирн В. (Составители и ред.), Proc. 12-я Северная Америка., For. Биол. Семинар: Роль физиологии и генетики в исследованиях и мониторинге лесных экосистем, Sault Ste. Мари О.Н., август 1992 г. Онт. Мин. Nat. Res., Ont. За. Res. Instit. И For. Кан., Онт. Область, край.
- ^ а б c d Ян, Сиянь и Чжан, Сяньлун (2010) «Регулирование соматического эмбриогенеза у высших растений», Critical Reviews in Plant Sciences, 29: 1, 36 - 57
- ^ а б Фон Арнольд С., Сабала И., Божков П., Дьячок Дж. И Филонова Л. (2002) Пути развития соматического эмбриогенеза. Растительная клетка Tiss Org. Культ. 69: 233–249
- ^ Хименес В.М., Томас С. (2005) Участие растительных гормонов в детерминации и прогрессировании соматического эмбриогенеза. в: Mujib A, Šamaj J (eds) Соматический эмбриогенез. Springer, Berlin, стр. 103–118.
- ^ Фехер, Аттила. Почему соматические клетки растений начинают формировать зародыши? В: Муджид, Абдул и Самадж, Йозеф. ред. Соматический эмбриогенез. Монографии по растительной клетке, Springer; Берлин / Гейдельберг, 2005, т. 2, стр. 85-101.
- ^ Тоонен, М.А. Дж., Хендрикс, Т., Шмидт, Э. Д. Л., Верховен, Х. А., ван Каммен, А. и Де Врис, С. С. 1994. Описание единичных клеток, образующих соматические эмбрионы, в суспензионных культурах моркови с использованием видео отслеживания клеток. Planta 194: 565–572.
- ^ Häggman H .; Vuosku J .; Сарджала Т .; Jokela A .; Ниеми К. Соматический эмбриогенез видов сосны: от функциональной геномики до плантационного лесоводства. Копать землю. Растительная клетка Monogr. 2: 119–140; 2006 г.
- ^ Муллин; Ли, ред. (2013). ЛУЧШАЯ ПРАКТИКА ДЕРЕВОДСТВА В ЕВРОПЕ. Скогфорск. ISBN 978-91-977649-6-4.
{kind=link}