Тиоредоксинредуктаза - Thioredoxin reductase
| Тиоредоксин-дисульфид редуктаза | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Кристаллическая структура человека тиоредоксинредуктаза 1; рендеринг на основе PDB: 2OHV. | |||||||||
| Идентификаторы | |||||||||
| Номер ЕС | 1.8.1.9 | ||||||||
| Количество CAS | 9074-14-0 | ||||||||
| Базы данных | |||||||||
| IntEnz | Просмотр IntEnz | ||||||||
| БРЕНДА | BRENDA запись | ||||||||
| ExPASy | Просмотр NiceZyme | ||||||||
| КЕГГ | Запись в KEGG | ||||||||
| MetaCyc | метаболический путь | ||||||||
| ПРИАМ | профиль | ||||||||
| PDB структуры | RCSB PDB PDBe PDBsum | ||||||||
| Генная онтология | AmiGO / QuickGO | ||||||||
| |||||||||
| Тиоредоксинредуктаза | |
|---|---|
| Идентификаторы | |
| Символ | ? |
| ИнтерПро | IPR005982 |
| PROSITE | PS00573 |
| SCOP2 | 1zof / Объем / СУПФАМ |
Тиоредоксинредуктазы (TR, TrxR) (ЕС 1.8.1.9 ) - единственные известные ферменты, снижающие тиоредоксин (Trx).[1] Были идентифицированы два класса тиоредоксинредуктазы: один класс у бактерий и некоторых эукариот и один у животных. У бактерий TrxR также катализирует восстановление глутаредоксиноподобных белков, известных как NrdH.[2][3][4] Оба класса флавопротеины которые функционируют как гомодимеры. Каждый мономер содержит FAD протезная группа, а НАДФН связывающий домен и активный сайт, содержащий редокс-активный дисульфидная связь.[5]
Сотовая роль
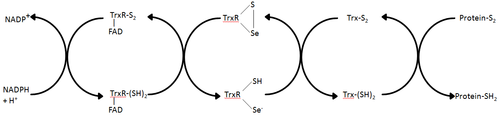
Тиоредоксинредуктаза - единственный известный фермент, который катализирует восстановление тиоредоксина.[1] и, следовательно, является центральным компонентом тиоредоксиновой системы. Вместе с тиоредоксином (Trx) и НАДФН наиболее общее описание этой системы - это метод образования восстановленных дисульфидных связей в клетках. Электроны отбираются от НАДФН через TrxR и переносятся в активный центр Trx, который продолжает восстанавливать дисульфиды белка или другие субстраты.[6] Система Trx существует во всех живых клетках и имеет эволюционную историю, связанную с ДНК как генетическим материалом, защитой от окислительного повреждения из-за метаболизма кислорода и редокс-сигнализацией с использованием таких молекул, как перекись водорода и оксид азота.[7][8]
Разнообразие
Два класса тиоредоксинредуктазы развились независимо:
- Тип с высокой молекулярной массой (MW = ~ 55,000), содержащий селеноцистеин остаток в его активном центре был идентифицирован у высших эукариот, включая человека. Этот TxR связан с глутатионредуктаза, трипанотионредуктаза, редуктаза ртути и липоамиддегидрогеназа.[5]
- Тип с низкой молекулярной массой (MW = ~ 35000) был идентифицирован у архей, бактерий и других эукарий.[5]
Эти два класса TrxR имеют только ~ 20% идентичности последовательностей в участке первичной последовательности, где они могут быть надежно выровнены.[5] Чистая реакция обоих классов TrxR идентична, но механизм действия каждого различен.[9]
Люди экспрессируют три изофермента тиоредоксинредуктазы: тиоредоксинредуктаза 1 (TrxR1, цитозольный), тиоредоксинредуктаза 2 (TrxR2, митохондриальная), тиоредоксинредуктаза 3 (TrxR3, тестис-специфичная).[10] Каждый изозим кодируется отдельным геном:
|
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Структура
Кишечная палочка
В Кишечная палочка ThxR существует два связывающих домена, один для FAD и еще один для НАДФН. Связь между этими двумя доменами представляет собой двухцепочечный антипараллельный β-лист.[11] Каждый домен в отдельности очень похож на аналогичные домены в глутатионредуктаза, и липоамиддегидрогеназа но их относительная ориентация этих доменов в ThxR повернута на 66 градусов.[11] Это становится важным в механизме действия фермента, который описан ниже. ThxR гомодимеризуется с границей раздела между двумя мономерами, образованными тремя альфа-спирали и две петли.[11] Каждый мономер может отдельно связывать молекулу тиоредоксин.

Структура Кишечная палочка Тиоредоксин, связанный с димером ThxR

Структура Кишечная палочка ThxR с маркированными простетическими группами FAD и NADPH


Млекопитающее
Структура TrxR млекопитающих похожа на Кишечная палочка. Он содержит FAD и НАДФН связывающий домен и интерфейс между двумя мономерными субъединицами. В ThxR млекопитающих есть вставка в FAD связывающий домен между двумя альфа-спиралями, который образует небольшую пару бета-цепей.[12] Активный дисульфид в ферменте расположен на одной из этих спиралей, и, таким образом, активная дисульфидная связь находится в FAD домен, а не НАДФН домен как в Кишечная палочка и другие прокариоты.[12]

Структура простетических групп ThxR FAD и NADPH человека

Механизм
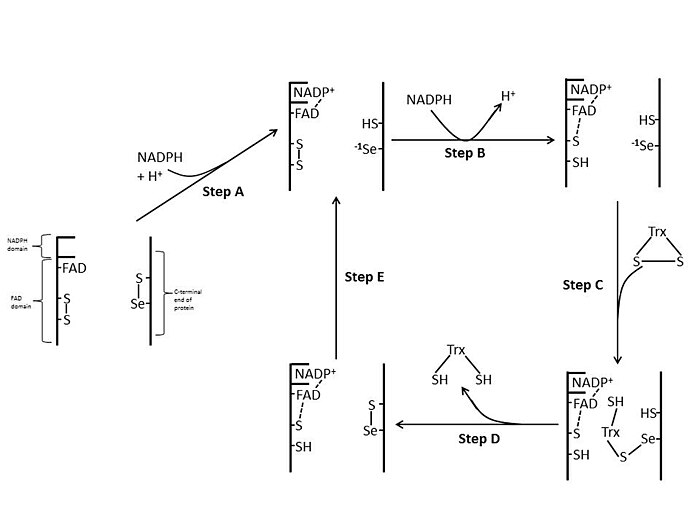
Кишечная палочка
В Кишечная палочка ThxR, пространственная ориентация доменов FAD и NADPH такова, что окислительно-восстановительные кольца FAD и NADPH не находятся в непосредственной близости друг от друга.[1] Когда домен FAD Кишечная палочка повернут на 66 градусов, при этом домен NADPH остается фиксированным, две простетические группы переходят в тесный контакт, позволяя электронам переходить от NADPH к FAD, а затем к дисульфидной связи активного центра.[1][15] Консервативные остатки активного сайта в E. coli представляют собой -Cys-Ala-Thr-Cys-.[1]
Млекопитающее
TrxR млекопитающих имеют гораздо более высокую гомологию последовательности с глутатионредуктазой, чем Кишечная палочка.[1] Остатки Cys активного центра в домене FAD и связанный домен NADPH находятся в непосредственной близости, что исключает необходимость поворота на 66 градусов для переноса электрона, обнаруженного в Кишечная палочка. Дополнительной особенностью механизма у млекопитающих является наличие остатка селеноцистеина на С-конце белка, который необходим для каталитической активности. Консервативные остатки в активном сайте млекопитающих представляют собой -Cys-Val-Asn-Val-Gly-Cys-.[1]
Клиническое значение
Лечение рака
Поскольку активность этого фермента важна для роста и выживания клеток, он является хорошей мишенью для противоопухолевой терапии. Кроме того, фермент активируется при нескольких типах рака, включая злокачественная мезотелиома.[16][17] Например, мотексафин гадолиний (MGd) - это новый химиотерапевтический агент, который избирательно воздействует на опухолевые клетки, что приводит к их гибели и апоптозу за счет ингибирования тиоредоксинредуктазы и рибонуклеотидредуктаза.
Кардиомиопатия
Дилатационная кардиомиопатия (DCM ) является распространенным диагнозом в случаях хроническая сердечная недостаточность. Тиоредоксинредуктазы являются важными белками для регулирования клеточного окислительно-восстановительного баланса и смягчения повреждений, вызванных: активные формы кислорода генерируется через окислительного фосфорилирования в митохондрии. Инактивация митохондриального TrxR2 у мышей приводит к истончению стенок желудочков сердца и неонатальной смерти.[10] Кроме того, две мутации в гене TrxR2 обнаружены у пациентов с диагнозом DCM, а не в контрольной популяции. Предполагается, что патологическим воздействием этих мутаций является нарушение способности контролировать окислительное повреждение в сердечные миоциты.[18]
Антибиотик
Недавно было проведено несколько исследований, показывающих, что низкомолекулярная тиоредоксинредуктаза может быть мишенью для новых антибиотиков (таких как ауранофин или эбселен.[19]) Это особенно актуально для Mycobacterium Haemophilum, и может использоваться для бактерий, устойчивых к антибиотикам.[20]
Рекомендации
- ^ а б c d е ж грамм Mustacich D, Powis G (февраль 2000 г.). «Тиоредоксинредуктаза». Биохимический журнал. 346, Чт 1 (1): 1–8. Дои:10.1042/0264-6021:3460001. ЧВК 1220815. PMID 10657232.
- ^ Джордан А., Аслунд Ф., Понтис Э., Райхард П., Холмгрен А. (июль 1997 г.). «Характеристика Escherichia coli NrdH. Глутаредоксин-подобный белок с тиоредоксиноподобным профилем активности». Журнал биологической химии. 272 (29): 18044–50. Дои:10.1074 / jbc.272.29.18044. PMID 9218434.
- ^ Phulera S, Mande SC (июнь 2013 г.). «Кристаллическая структура Mycobacterium tuberculosis NrdH при 0,87 Å указывает на возможный режим ее активности». Биохимия. 52 (23): 4056–65. Дои:10.1021 / bi400191z. PMID 23675692.
- ^ Фулера С., Акиф М., Сардесай А.А., Манде С.К. (01.01.2014). «Редокс-белки Mycobacterium tuberculosis». Журнал Индийского института науки. 94 (1): 127–138. ISSN 0970-4140.
- ^ а б c d Хирт Р.П., Мюллер С., Эмбли TM, Кумбс Г.Х. (июль 2002 г.). «Разнообразие и эволюция тиоредоксинредуктазы: новые перспективы». Тенденции в паразитологии. 18 (7): 302–8. Дои:10.1016 / S1471-4922 (02) 02293-6. PMID 12379950.
- ^ а б Холмгрен А., Лу Дж. (Май 2010 г.). «Тиоредоксин и тиоредоксинредуктаза: текущие исследования с особым вниманием к болезням человека». Сообщения о биохимических и биофизических исследованиях. 396 (1): 120–4. Дои:10.1016 / j.bbrc.2010.03.083. PMID 20494123.
- ^ Мейер Y, Бьюкенен BB, Vignols F, Reichheld JP (2009). «Тиоредоксины и глутаредоксины: объединяющие элементы в окислительно-восстановительной биологии». Ежегодный обзор генетики. 43: 335–67. Дои:10.1146 / annurev-genet-102108-134201. PMID 19691428.
- ^ Lillig CH, Holmgren A (январь 2007 г.). «Тиоредоксин и родственные ему молекулы - от биологии до здоровья и болезней». Антиоксиданты и редокс-сигналы. 9 (1): 25–47. Дои:10.1089 / ars.2007.9.25. PMID 17115886.
- ^ Арскотт Л.Д., Громер С., Ширмер Р.Х., Беккер К., Уильямс С.Х. (апрель 1997 г.). «Механизм тиоредоксинредуктазы из плаценты человека подобен механизмам липоамиддегидрогеназы и глутатионредуктазы и отличается от механизма тиоредоксинредуктазы из Escherichia coli». Труды Национальной академии наук Соединенных Штатов Америки. 94 (8): 3621–6. Дои:10.1073 / пнас.94.8.3621. ЧВК 20490. PMID 9108027.
- ^ а б Конрад М., Якупоглу С., Морено С.Г., Липпл С., Банджак А., Шнайдер М., Бек Х., Хацопулос А.К., Джаст У, Синоватц Ф., Шмаль В., Чиен К.Р., Вурст В., Борнкамм Г.В., Бриельмайер М. (ноябрь 2004 г.) «Важная роль митохондриальной тиоредоксинредуктазы в кроветворении, развитии сердца и сердечной функции». Молекулярная и клеточная биология. 24 (21): 9414–23. Дои:10.1128 / MCB.24.21.9414-9423.2004. ЧВК 522221. PMID 15485910.
- ^ а б c Уильямс CH (октябрь 1995 г.). «Механизм и структура тиоредоксинредуктазы Escherichia coli». Журнал FASEB. 9 (13): 1267–76. Дои:10.1096 / fasebj.9.13.7557016. HDL:2027.42/154540. PMID 7557016. S2CID 26055087.
- ^ а б Сандалова Т., Чжун Л., Линдквист Ю., Хольмгрен А., Шнайдер Г. (август 2001 г.). «Трехмерная структура тиоредоксинредуктазы млекопитающих: значение для механизма и эволюции селеноцистеин-зависимого фермента». Труды Национальной академии наук Соединенных Штатов Америки. 98 (17): 9533–8. Дои:10.1073 / pnas.171178698. ЧВК 55487. PMID 11481439.
- ^ Чжун Л., Арнер Э.С., Холмгрен А. (май 2000 г.). «Структура и механизм тиоредоксинредуктазы млекопитающих: активный центр представляет собой окислительно-восстановительный селенолтиол / селененилсульфид, образованный из консервативной последовательности цистеин-селеноцистеин». Труды Национальной академии наук Соединенных Штатов Америки. 97 (11): 5854–9. Дои:10.1073 / pnas.100114897. ЧВК 18523. PMID 10801974.
- ^ Беккер К., Херольд-Менде С., Парк Дж. Дж., Лоу Дж., Ширмер Р. Х. (август 2001 г.). «Человеческая тиоредоксинредуктаза эффективно ингибируется комплексами (2,2 ': 6', 2 '' -терпиридин) платины (II). Возможные последствия для новой противоопухолевой стратегии». Журнал медицинской химии. 44 (17): 2784–92. Дои:10.1021 / jm001014i. PMID 11495589.
- ^ Леннон Б.В., Уильямс СН (август 1997 г.). «Восстановительная полуреакция тиоредоксинредуктазы из Escherichia coli». Биохимия. 36 (31): 9464–77. Дои:10.1021 / bi970307j. PMID 9235991.
- ^ Nilsonne G, Sun X, Nyström C, Rundlöf AK, Potamitou Fernandes A, Björnstedt M, Dobra K (сентябрь 2006 г.). «Селенит вызывает апоптоз в клетках саркоматоидной злокачественной мезотелиомы через окислительный стресс». Свободная радикальная биология и медицина. 41 (6): 874–85. Дои:10.1016 / j.freeradbiomed.2006.04.031. PMID 16934670.
- ^ Kahlos K, Soini Y, Säily M, Koistinen P, Kakko S, Pääkkö P, Holmgren A, Kinnula VL (май 2001 г.). «Повышение тиоредоксина и тиоредоксинредуктазы при злокачественной мезотелиоме плевры человека». Международный журнал рака. 95 (3): 198–204. Дои:10.1002 / 1097-0215 (20010520) 95: 3 <198 :: AID-IJC1034> 3.0.CO; 2-F. PMID 11307155.
- ^ Sibbing D, Pfeufer A, Perisic T, Mannes AM, Fritz-Wolf K, Unwin S, Sinner MF, Gieger C, Gloeckner CJ, Wichmann HE, Kremmer E, Schäfer Z, Walch A, Hinterseer M, Näbauer M, Kääb S, Кастрати А., Шёмиг А., Мейтингер Т., Борнкамм Г. В., Конрад М., фон Бекерат Н. (май 2011 г.). «Мутации в митохондриальном гене тиоредоксинредуктазы TXNRD2 вызывают дилатационную кардиомиопатию». Европейский журнал сердца. 32 (9): 1121–33. Дои:10.1093 / eurheartj / ehq507. PMID 21247928.
- ^ Marshall AC, Kidd SE, Lamont-Friedrich SJ, Arentz G, Hoffmann P, Coad BR, Bruning JB (март 2019 г.). «Тиоредоксинредуктаза Aspergillus fumigatus». Противомикробные препараты и химиотерапия. 63 (3). Дои:10.1128 / AAC.02281-18. ЧВК 6395915. PMID 30642940.
- ^ Харбут М.Б., Вильчез С., Луо Х, Хенслер М.Э., Го Х, Ян Б. и др. (Апрель 2015 г.). «Ауранофин оказывает бактерицидное действие широкого спектра действия, воздействуя на тиоловый окислительно-восстановительный гомеостаз». Труды Национальной академии наук Соединенных Штатов Америки. 112 (14): 4453–8. Дои:10.1073 / pnas.1504022112. ЧВК 4394260. PMID 25831516.
внешняя ссылка
- Тиоредоксин + Редуктаза + (НАДФН) в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)