(ацил-белок-носитель) S-малонилтрансфераза - (acyl-carrier-protein) S-malonyltransferase
| Малонил-КоА: ACP трансацилаза | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Идентификаторы | |||||||||
| Номер ЕС | 2.3.1.39 | ||||||||
| Количество CAS | 37257-17-3 | ||||||||
| Базы данных | |||||||||
| IntEnz | Просмотр IntEnz | ||||||||
| БРЕНДА | BRENDA запись | ||||||||
| ExPASy | Просмотр NiceZyme | ||||||||
| КЕГГ | Запись в KEGG | ||||||||
| MetaCyc | метаболический путь | ||||||||
| ПРИАМ | профиль | ||||||||
| PDB структуры | RCSB PDB PDBe PDBsum | ||||||||
| Генная онтология | AmiGO / QuickGO | ||||||||
| |||||||||
В энзимология, а [ацил-белок-носитель] S-малонилтрансфераза (EC 2.3.1.39 ) является фермент который катализирует то химическая реакция
- малонил-КоА + белок-носитель ацила ⇌ КоА + малонил- [белок-носитель ацила]
Таким образом, два субстраты этого фермента малонил-КоА и белок-носитель ацила, а его два товары находятся CoA и белок-носитель малонилацила. Этот фермент принадлежит к семейству трансферазы особенно те ацилтрансферазы переносящие группы, отличные от аминоацильных групп. Этот фермент участвует в биосинтез жирных кислот.
Номенклатура
В систематическое название этого класса ферментов является малонил-КоА: [белок-ацил-носитель] S-малонилтрансфераза. Другие широко используемые названия включают трансацилазу белка-носителя малонил-кофермента А-ацила,
- [белок-носитель ацила] малонилтрансфераза,
- FabD,
- малонилтрансацилаза,
- малонилтрансфераза,
- трансацилаза белка-носителя малонил-КоА-ацила,
- малонил-КоА: АСР трансацилаза,
- малонил-КоА: трансацилаза AcpM,
- малонил-КоА: трансацилаза белка-носителя ацила,
- МАТ и
- MCAT.
Структура
Кристаллические структуры FabD из Кишечная палочка [1] и Streptomyces coelicolor [2] известны и позволяют лучше понять каталитический механизм FabD. В E.Coli, FabD в первую очередь участвует в пути FAS. Однако в Streptomyces coelicolorFabD участвует в путях FAS и поликетидсинтазы. В обоих случаях структуры и активные сайты очень похожи.

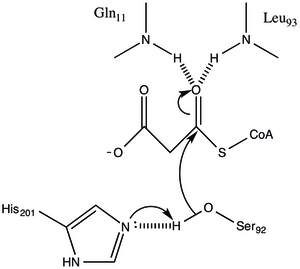
Белок имеет архитектуру типа α / β, но складка уникальна. активный центр, полученный из расположения каталитического Ser92, содержит типичный нуклеофильный изгиб, наблюдаемый в α / β гидролазах.[1] Серин 92 связан водородными связями с His 201 аналогично различным сериновым гирдолазам. однако вместо карбоновой кислоты, обычно присутствующей в каталитических триадах, карбонил основной цепи Gln 250 служит акцептором водородной связи при взаимодействии с His 201.[1] Два других остатка, Arg-117 и Glu-11, также расположены в активном сайте, но их функция не ясна.
Функция
В жирная кислота синтетический путь является основным путем производства мембранных фосфолипид ацильные цепи у бактерий и растений.[3] Последовательность реакции осуществляется серией отдельных растворимых белков, каждый из которых кодируется отдельным геном, а промежуточные продукты пути перемещаются между ферментами.[3] Малони-КоА: ACP-трансацилаза (FabD) является одним из таких индивидуальных растворимых белков и катализирует следующую реакцию:
- малонил-КоА + белок-носитель ацила ⇌ КоА + малонил- [белок-носитель ацила]
Передача малонатный в ацильный белок-носитель (ACP) превращает ацильные группы в тиоэфирные формы, которые характерны для ацильных промежуточных продуктов в синтезе жирных кислот и которые строго необходимы для реакций конденсации, катализируемых β-кетоацил-ACP синтетазой.[4]
Механизм
Малонил-КоА: ACP-трансацилаза использует кинетический механизм пинг-понга со связанным сложным эфиром малоновой кислоты в качестве промежуточного ацила, присоединенного к остатку серина, находящемуся в пентапептиде GHSLG.[5] Сначала FabD связывает малонил-КоА, затем малониловый фрагмент переносится на активный сайт Ser 92, и КоА высвобождается из фермента. Затем АСР связывается, и малонильный фрагмент переносится на концевой сульфгидрил простетической группы АСР. Эта реакция легко обратима.[3][6]

Промышленная значимость
Среди всех известных метаболических путей в живых системах биосинтез жирных кислот дает наиболее энергоемкие продукты.[7] В результате производные жирных кислот микробного происхождения становятся многообещающей возобновляемой энергетической альтернативой транспортному топливу, полученному из ископаемого топлива. Недавно Khosla et al.[7] разработали процедуру восстановления синтазы жирных кислот E.Coli с использованием очищенных белковых компонентов (включая FabD) и сообщили о подробном кинетическом анализе этой восстановленной системы in vitro.[8] Их открытие обеспечивает новую основу для оценки масштабов и ограничений использования E.Coli в качестве биокатализатора для производства дизельного топлива.
Клиническая значимость
FabD как цель для открытия антибактериальных лекарств: предстоящая область
Биосинтез жирных кислот осуществляется повсеместно Синтаза жирных кислот.[9] Синтаза жирных кислот пути делятся на две отдельные молекулярные формы: Тип I и Тип II.[10] В Типе I, Синтаза жирных кислот (обнаружен у людей и других млекопитающих) представляет собой один большой полипептид, состоящий из нескольких отдельных доменов.[11] С другой стороны, каждая ферментативная активность (Реакция конденсации, Редукционная реакция, Реакция обезвоживания ) обнаруживается как дискретный белок в системах типа II.[12] Различие в организации активного центра и преобладание систем FAS типа II у бактерий делают ферменты этого пути привлекательными мишенями для антибактериальных препаратов.[9][12]
FabD (ацил-белок-носитель S-малонилтрансфераза) является разумной мишенью, учитывая, что доступна кристаллическая структура с высоким разрешением.[9] Однако ни о каких ингибиторах FabD не сообщалось в литературе и обзорных статьях по этой теме.[9] Простая структура и кислотность малоната, по-видимому, позволяют несколько подходов к синтезу производных (действующих как потенциальные ингибиторы), которые сохраняют характер молекулы.
Второй подход к использованию FabD в качестве мишени для лекарственного средства часто упоминается в литературе: FabD может предоставить полезную метку для определения местоположения генов fab, поскольку ген FabD обычно соседствует по крайней мере с одним другим геном fab.[13] Однако (по состоянию на 2015 год) ни одно потенциальное лекарство не пыталось использовать эту функцию.
Рекомендации
- ^ а б c d Serre L, Verbree EC, Dauter Z, Stuitje AR, Derewenda ZS (июнь 1995 г.). «Малонил-КоА Escherichia coli: трансацилаза белка-носителя ацила с разрешением 1,5А. Кристаллическая структура компонента синтазы жирных кислот». Журнал биологической химии. 270 (22): 12961–4. Дои:10.1074 / jbc.270.22.12961. PMID 7768883.
- ^ Китинг-Клей А.Т., Шелат А.А., Сэвидж Д.Ф., Цай С.К., Мирке Л.Дж., О'Коннелл Д.Д., Хосла С., Страуд Р.М. (февраль 2003 г.). «Катализ, специфичность и сайт стыковки ACP Streptomyces coelicolor malonyl-CoA: ACP transacylase». Структура. 11 (2): 147–54. Дои:10.1016 / s0969-2126 (03) 00004-2. PMID 12575934.
- ^ а б c d White SW, Zheng J, Zhang YM (июнь 2005 г.). «Структурная биология биосинтеза жирных кислот II типа». Ежегодный обзор биохимии. 74 (1): 791–831. Дои:10.1146 / annurev.biochem.74.082803.133524. PMID 15952903.
- ^ Ruch FE, Vagelos PR (декабрь 1973 г.). «Выделение и общие свойства трансацилазы малонил-кофермента А-ацил-переносчика белка Escherichia coli». Журнал биологической химии. 248 (23): 8086–94. PMID 4584822.
- ^ Ruch FE, Vagelos PR (декабрь 1973 г.). «Изоляция и общие свойства кишечная палочка трансацилаза белка-носителя малонил-кофермента А-ацила ". Журнал биологической химии. 248 (23): 8086–94. PMID 4584822.
- ^ Офнер К., Шульц Х., Д'Арси А., Дейл Г.Е. (июнь 2006 г.). «Картирование активного сайта трансацилазы белка-носителя малонил-CoA-ацила Escherichia coli (FabD) с помощью кристаллографии белка». Acta Crystallographica Раздел D. 62 (Pt 6): 613–618. Дои:10.1107 / S0907444906009474. PMID 16699188.
- ^ а б Лю Т., Хосла С. (2010). «Генная инженерия Escherichia coli для производства биотоплива». Ежегодный обзор генетики. 44: 53–69. Дои:10.1146 / annurev-genet-102209-163440. PMID 20822440.
- ^ Ю X, Лю Т., Чжу Ф., Хосла С. (ноябрь 2011 г.). «Восстановление in vitro и стационарный анализ синтазы жирных кислот из Escherichia coli». Труды Национальной академии наук Соединенных Штатов Америки. 108 (46): 18643–8. Bibcode:2011PNAS..10818643Y. Дои:10.1073 / pnas.1110852108. ЧВК 3219124. PMID 22042840.
- ^ а б c d Payne DJ, Warren PV, Holmes DJ, Ji Y, Lonsdale JT (май 2001 г.). «Биосинтез жирных кислот бактерий: управляемая геномом цель для открытия антибактериальных препаратов». Открытие наркотиков сегодня. 6 (10): 537–544. Дои:10.1016 / S1359-6446 (01) 01774-3. PMID 11369293.
- ^ Rock CO, Cronan JE (июль 1996 г.). «Escherichia coli как модель для регуляции биосинтеза диссоциируемых (тип II) жирных кислот». Biochimica et Biophysica Acta (BBA) - липиды и липидный метаболизм. 1302 (1): 1–16. Дои:10.1016/0005-2760(96)00056-2. PMID 8695652.
- ^ Чирала С.С., Хуанг В.Й., Джаякумар А., Сакаи К., Вакил С.Дж. (май 1997 г.). «Синтаза жирных кислот животных: функциональное картирование, клонирование и экспрессия составляющих домена I активности». Труды Национальной академии наук Соединенных Штатов Америки. 94 (11): 5588–93. Bibcode:1997PNAS ... 94.5588C. Дои:10.1073 / пнас.94.11.5588. ЧВК 20822. PMID 9159116.
- ^ а б Цай Дж. Т., О В., Ларсон Т. Дж., Яковски С., Рок Колорадо (апрель 1992 г.). «Выделение и характеристика гена бета-кетоацил-ацил-носителя белка синтазы III (fabH) из Escherichia coli K-12». Журнал биологической химии. 267 (10): 6807–14. PMID 1551888.
- ^ Кэмпбелл Дж. У., Кронан Дж. Э. (2001). «Биосинтез бактериальных жирных кислот: цели для открытия антибактериальных препаратов». Ежегодный обзор микробиологии. 55: 305–32. Дои:10.1146 / annurev.micro.55.1.305. PMID 11544358.
дальнейшее чтение
- Альбертс AW, Majerus PW, Vagelos PR (1969). «Трансацилаза белка-носителя ацетил-СоА ацила». Методы Энзимол. Методы в энзимологии. 14: 50–53. Дои:10.1016 / S0076-6879 (69) 14009-4. ISBN 978-0-12-181871-5.
- Прескотт DJ, Вагелос PR (1972). «Ацильный белок-носитель». Достижения в энзимологии и смежных областях молекулярной биологии. Достижения в энзимологии и смежных областях молекулярной биологии. 36. С. 269–311. Дои:10.1002 / 9780470122815.ch8. ISBN 9780470122815. PMID 4561013.
- Уильямсон И.П., Вакил С.Дж. (май 1966 г.). «Исследования механизма синтеза жирных кислот. XVII. Получение и общие свойства трансацилаз ацетилкофермента А и малонилкофермента А-ацил-носителя белка». Журнал биологической химии. 241 (10): 2326–32. PMID 5330116.
- Джоши В.К., Вакил С.Дж. (апрель 1971 г.). «Исследования механизма синтеза жирных кислот. XXVI. Очистка и свойства малонил-кофермента А - трансацилазы белка-носителя ацила Escherichia coli». Архивы биохимии и биофизики. 143 (2): 493–505. Дои:10.1016/0003-9861(71)90234-7. PMID 4934182.
- Кремер Л., Нампотири К.М., Лешин С., Довер Л.Г., Грэм С., Беттс Дж., Бреннан П.Дж., Минникин Д.Е., Лохт С., Бесра Г.С. (июль 2001 г.). «Биохимическая характеристика белка-носителя ацила (AcpM) и малонил-КоА: трансацилаза AcpM (mtFabD), двух основных компонентов синтазы жирных кислот II Mycobacterium tuberculosis». Журнал биологической химии. 276 (30): 27967–74. Дои:10.1074 / jbc.M103687200. PMID 11373295.
- Китинг-Клей А.Т., Шелат А.А., Сэвидж Д.Ф., Цай С.К., Мирке Л.Дж., О'Коннелл Д.Д., Хосла С., Страуд Р.М. (февраль 2003 г.). «Катализ, специфичность и сайт стыковки АСР Streptomyces coelicolor малонил-КоА: АСР трансацилаза». Структура. 11 (2): 147–54. Дои:10.1016 / S0969-2126 (03) 00004-2. PMID 12575934.
- Szafranska AE, Hitchman TS, Cox RJ, Crosby J, Simpson TJ (февраль 2002 г.). «Кинетический и механистический анализ трансацилазы малонил-КоА: АСР из Streptomyces coelicolor указывает на единственный каталитически компетентный сериновый нуклеофил в активном центре». Биохимия. 41 (5): 1421–7. Дои:10.1021 / bi012001p. PMID 11814333.