Nassellaria - Nassellaria
| Nassellaria | |
|---|---|
 | |
| Научная классификация | |
| Домен: | |
| (без рейтинга): | |
| (без рейтинга): | |
| Тип: | |
| Подтип: | |
| Учебный класс: | |
| Заказ: | Nassellaria Эренберг, 1875 |
| Семьи | |
Nassellaria это приказ Ризария принадлежность к классу Радиолярии.[1] Для организмов этого отряда характерно сшивание скелета с конусом или кольцом.
Вступление
Насселлария - это орден Радиолярии под классом Поликистина. Эти организмы представляют собой одноклеточный эукариотический гетеротрофный планктон, как правило, с кремнистым конусообразным скелетом. Наиболее распространенной группой радиолярий являются поликистиновые радиолярии, которые делятся на две подгруппы: спумелларии и насселларии. И спумелларии, и насселларии обычны. черт -формирование микрофоссилий и важны для стратиграфического датирования, так как самые старые радиолярии имеют докембрийский возраст. Насселларии появляются в летописи окаменелостей намного позже, чем их другие поликистиновые родственники, спумелларии. spumellarians можно увидеть еще в докембрии, тогда как nassellarians не начинают появляться до Каменноугольный. Считается, что разнообразие видов насселларийцев увеличивалось с начала Кайнозойский.[2]
Насселларианцы были и остаются одними из самых замечательных и эстетически интересных протистов как живых, так и в летописи окаменелостей. Они играют важную экологическую роль, и сделали это с Докембрийский. Мы еще многого не знаем об этих протисты, и открытие большего потребует постоянных междисциплинарных усилий многих ученых.
Морфология
В голотип морфология Насселларианского ордена, описанного Андерсоном[2] и Болтовский и др.[3] имеет яйцевидную центральную капсулу (часть клетки, содержащую один или несколько ядра, Тела Гольджи, митохондрии, лизосомы, и другие тела, важные для клеточной функции), расположенные внутри пористого конического скелета, состоящего из кремнезем. Центральная капсула обычно расположена ближе к апикальный конец скелета. В базальный конец обычно имеет форму полого круга, что придает скелету конический вид. Экстракапсулум окружает центральную капсулу, но все еще внутри скелета. Это в первую очередь альвеолы, наполненные газом пузырьковые структуры, регулирующие плавучесть организма. Альвеолы соединены сетью ризоподий по всей экстракапсуле. Фузулы соединяют интракапсулум (центральную капсулу) с экстракапсулой. Аксоподии и дополнительные фузулы выходят из базального отверстия и проходят через поры скелета. Они будут выступать в качестве кормовых аппаратов для населярии.
- скелеты

Anthocyrtium hispidum

Антоцирт гроссулярия

Артостробус sp.
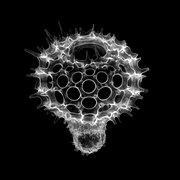
Калоциклома sp.

Калоциклы sp.

Eucyrtidium sp.

Eusyringium sipho

Лихноканома sp.







Lithochytris pyramidalis

Lychnocanium ventricosum

Подоциртис принцепс

Podocyrtis mitra

Podocyrtis papalis
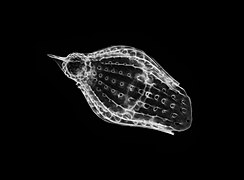
Podocyrtis papalis

Феокотилиса фикус sp.







Выделяют три основных типа скелетов населярий.[2] Первый образуется из соединения трех удлиненных спикулы в центральной точке. Три спикулы расположены в одной плоскости на расстоянии 120 градусов друг от друга. Это обычно называется «штативной» конструкцией. Второй тип скелета, распространенный у насселларий, - это уже упомянутый конический пористый скелет. Третий тип - кольцо, окружающее решетчатую оболочку. Эти три типа скелетов могут быть смешаны и сопоставлены для создания семи категорий скелетов у населлариан.[2] Даже простейшие классификации морфологии насселлярий сложны из-за большого разнообразия морфологий в отряде насселлярий даже по сравнению с их сестринской группой, спумелларий. Врожденная природа насселларийной геометрии допускает разнообразие, что может быть одним из факторов, ведущих к быстрой диверсификации насселларий по сравнению со спумелларианами во время Кайнозойский.[2]
Многие насселлярии сегментированы, что означает, что есть круговые скелетные подразделения, которые существуют экваториально вдоль конуса скелета. Эти разделения называются стриктурами.[2] Стриктуры делят организм на сегменты, обычно придавая ему более выпуклую форму. Некоторые насселларии также имеют шипы на апикальном конце или вдоль боковых сторон конуса. Шипы вдоль базального отверстия организма называются стопами.[2]
Экология
Дом многих насселларианцев динофлагеллята симбионты в рамках своих тестов.[3] Насселлариан обеспечивает аммоний и углекислый газ для динофлагелляты, в то время как динофлагеллята обеспечивает слизистую оболочку носовой части, полезную для охоты и защиты от вредных захватчиков.[2] Имеются данные о малых субъединицах рибосомных ДНК анализ этих симбионтов динофлагеллят, которые в симбиозе динофлагеллят с радиоляриями эволюционировали независимо от других симбиозов динофлагеллят (например, фораминиферы ). Анализ малых субъединиц рибосомной ДНК также показывает доказательства того, что коэволюции динофлагеллят и радиолярий не происходило.[4]
Насселларийское питание в основном осуществляется за счет продления псевдоподия из базального отверстия. Осевой выступ очень длинный псевдоподиум которые выходят из базальной апертуры и окружены более короткими концевыми выступами, которые образуют конический конус вокруг осевого выступа. Питание сегментированных насселларийцев осуществляется в три фазы: 1) вытягивание, 2) захват и 3) втягивание.[5] Геометрия nasselarians весьма полезна для этого механизма питания, поскольку пища проходит через базальное отверстие, которое намного больше, чем поры в остальной части скелета, именно так большинство spumellarians должны транспортировать пищу.
Вторая разновидность насселларийцев питается вообще без аксиальной проекции. Вместо этого эти организмы образуют широкий конус, который они выбрасывают за собой из базального отверстия, как рыболовную сеть. Три фазы питания, используемые сегментированными насселлариями, также очевидны у этих насселларий.[5] Эти «рыболовные сети» насселларии гораздо легче увидеть кормящихся в дикой природе, и они представляют собой довольно захватывающее зрелище.
Населляры питаются другим планктоном, таким как мелкие водоросли, бактерии, диатомовые водоросли и мелкий зоопланктон.[2][3][5]
История окаменелостей
Рано Палеозой В ископаемых радиоляриях доминируют Spumellaria до тех пор, пока Каменноугольный период, в течение которого фауна насселларий пережила резкое увеличение разнообразия. Насселларийское и спумеллярное разнообразие было относительно схожим с Мезозойский, с уменьшением разнообразия после массовое вымирание событий и рост разнообразия спумелларий и насселлярий во время Четвертичный.[2]
Симбиоз между водорослями и радиоляриями часто наблюдается у сохранившийся видов, но эволюция и время этого симбиоза в настоящее время неизвестны, так как симбиотические водоросли не оставляют после себя твердых скелетов для окаменения. Ответить на этот вопрос можно, используя изотопный анализ, так как водорослевые симбионты преимущественно поглощают углерод-12, поэтому симбионтсодержащие известковые организмы, такие как Фораминиферы обогатиться углерод-13 по сравнению с известковыми организмами, не несущими симбионтов.[2] Если ископаемые свидетельства могут быть собраны для анализа изотопного соотношения, временное окно для появления водорослевых симбионтов с насселлариями и / или спумеллариями должно быть ограничено геологическим временем.
Рекомендации
- ^ "Radiolaria.org> Миоцен". radiolaria.org.
- ^ а б c d е ж грамм час я j k Андерсон, О. Р. (1983). Радиолярии. Springer Science & Business Media.
- ^ а б c Болтовской, Деметрио; Андерсон, О. Роджер; Корреа, Нэнси М. (2017). Справочник протистов. Спрингер, Чам. С. 731–763. Дои:10.1007/978-3-319-28149-0_19. ISBN 9783319281476.
- ^ Gast, R.J .; Кэрон, Д. А. (1996-11-01). «Молекулярная филогения симбиотических динофлагеллат из планктонных фораминифер и радиолярий».. Молекулярная биология и эволюция. 13 (9): 1192–1197. Дои:10.1093 / oxfordjournals.molbev.a025684. ISSN 0737-4038. PMID 8896371.
- ^ а б c Мацуока, Ацуши (01.09.2007). «Механизмы питания живых радиолярий: новый взгляд на морские экосистемы прошлого». Швейцарский журнал наук о Земле. 100 (2): 273–279. Дои:10.1007 / s00015-007-1228-у. ISSN 1661-8726.
| Этот Радиолярий -связанная статья является заглушка. Вы можете помочь Википедии расширяя это. |