Вирус с отрицательной цепью РНК - Negative-strand RNA virus
| Негарнавирикота | |
|---|---|
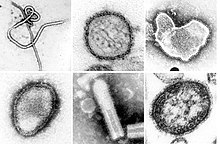 | |
| Монтаж просвечивающие электронные микрофотографии некоторых вирусов в типе Негарнавирикота. Не в масштабе. Виды слева направо, сверху вниз: Заир эболавирус, Ортоантавирус Sin Nombre, Ортопневмовирус человека, Hendra henipavirus, неопознанный рабдовирус, Морбилливирус кори. | |
| Классификация вирусов | |
| (без рейтинга): | Вирус |
| Область: | Рибовирия |
| Королевство: | Орторнавиры |
| Тип: | Негарнавирикота |
| Субналоги | |
| Синонимы[1][2] | |
| |
Вирусы с отрицательной цепью РНК (-ssRNA вирусы) являются группой связанных вирусы который имеет отрицательный смысл, одноцепочечные геномы из рибонуклеиновая кислота. У них есть геномы, которые действуют как дополнительные цепи, из которых информационная РНК (мРНК) синтезируется вирусным ферментом РНК-зависимая РНК-полимераза (RdRp). Во время репликации вирусного генома RdRp синтезирует антигеном с положительным смыслом, который он использует в качестве матрицы для создания геномной РНК с отрицательным смыслом. Вирусы с РНК с отрицательной цепью также обладают рядом других характеристик: большинство из них содержат вирусный конверт окружающий капсид, в который заключен вирусный геном, геномы -ssRNA вируса обычно линейны, и их геном обычно сегментируется.
Вирусы с отрицательной цепью РНК составляют филюм Негарнавирикота, в королевстве Орторнавиры и царство Рибовирия. Они произошли от общего предка, который был двухцепочечная РНК (дцРНК) вируса, и они считаются родственными клада реовирусы, которые представляют собой вирусы дцРНК. Внутри типа есть две основные ветви, которые образуют два подтипа: Гапловирикотина, члены которого в основном несегментированы и которые кодируют RdRp, который синтезирует кэпы на мРНК, и Полипловирикотина, члены которого сегментированы и кодируют RdRp, хватает кепки из мРНК хозяина. Всего распознается шесть классов этого типа.
Вирусы с отрицательной цепью РНК тесно связаны с членистоногие и могут быть неофициально разделены на тех, которые зависят от членистоногих в передаче, и тех, которые произошли от вирусов членистоногих, но теперь могут воспроизводиться у позвоночных без помощи членистоногих. Известные вирусы -ssRNA, переносимые членистоногими, включают: Вирус лихорадки Рифт-Валли и вирус пятнистого увядания томатов. Известные вирусы -ssRNA позвоночных включают Вирус Эбола, хантавирусы, вирусы гриппа, то Вирус лихорадки Ласса, а вирус бешенства.
Этимология
Негарнавирикота берет первую часть своего названия от латинский нега, что означает отрицательный, средняя часть rna относится к РНК, и последняя часть, -вирикота, суффикс, используемый для типа вируса. Подтип Гапловирикотина берет первую часть своего имени, Haplo, из Древнегреческий ἁπλός, что означает простой, и -вирикотина суффикс, используемый для подтипа вируса. Подтип Полипловирикотина следует той же схеме, Полипло взяты из древнегреческого πολύπλοκος, что означает комплекс.[1]
Характеристики
Геном

Все вирусы в Негарнавирикота представляют собой вирусы с одноцепочечной РНК с отрицательным смыслом (-ssRNA). У них есть геномы, состоящие из РНК, которые являются одноцепочечными, а не двухцепочечными. Их геномы имеют отрицательный смысл, что означает, что информационная РНК (мРНК) может быть синтезирована непосредственно из генома вирусным ферментом РНК-зависимой РНК-полимеразой (RdRp), также называемой РНК-репликазой, которая кодируется всеми вирусами -ssRNA. Исключение вирусов из рода Тенуивирус и некоторые в семье Chuviridae все -ssRNA вирусы имеют линейные, а не кольцевые геномы, и эти геномы могут быть сегментированными или несегментированными.[1][3][4] Все геномы -ssРНК содержат концевые инвертированные повторы, которые представляют собой палиндромные нуклеотидные последовательности на каждом конце генома.[5]
Репликация и транскрипция
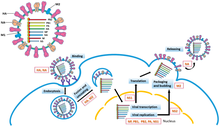
Репликация геномов -ssRNA выполняется с помощью RdRp, который инициирует репликацию путем связывания с лидерной последовательностью на 3'-конце (обычно произносится как «три первичных конца») генома. Затем RdRp использует геном с отрицательным смыслом в качестве матрицы для синтеза антигенома с положительным смыслом. При репликации антигенома RdRp сначала связывается с концевой последовательностью на 3'-конце антигенома. После этого RdRp игнорирует все сигналы транскрипции на антигеноме и синтезирует копию генома, используя антигеном в качестве матрицы.[6] Репликация выполняется, пока геном находится внутри нуклеокапсида, а RdRp раскрывает капсид и перемещается по геному во время репликации. По мере того как новые нуклеотидные последовательности синтезируются с помощью RdRp, капсидные белки собираются и инкапсидируют вновь реплицируемую вирусную РНК.[2]
Транскрибирование мРНК из генома следует по той же схеме направленности, что и создание антигенома. В лидерной последовательности RdRp синтезирует 5-концевую (обычно произносится как «пять простых концов») трифосфат-лидерную РНК и либо, в случае подтипа Гапловирикотина, закрывает 5'-конец или, в случае подтипа Полипловирикотина, хватает кепку от мРНК хозяина и прикрепляет ее к вирусной мРНК, так что мРНК может быть переведено клеткой-хозяином рибосомы.[7][8][9]
После кэпирования мРНК RdRp инициирует транскрипцию в стартовый кодон а затем прекращает транскрипцию при достижении стоп-кодон. В конце транскрипции RdRp синтезирует полиадиновый хвост (polyA tail) состоящий из сотен аденины на 3-конце мРНК, что может быть сделано заикание на последовательности урацилы. После конструирования хвоста полиА мРНК высвобождается с помощью RdRp. В геномах, которые кодируют более одной транскрибируемой части, RdRp может продолжить сканирование до следующего стартового кодона, чтобы продолжить транскрипцию.[7][10]
Некоторые -ssRNA вирусы амбисенс, что означает, что и отрицательная геномная цепь, и положительный антигеном по отдельности кодируют разные белки. Чтобы транскрибировать вирусы амбисенса, выполняются два цикла транскрипции: во-первых, мРНК продуцируется непосредственно из генома; во-вторых, мРНК создается из антигенома. Все вирусы амбисенса содержат петля для шпильки структура, чтобы остановить транскрипцию после того, как мРНК белка была транскрибирована.[11]
Морфология

Вирусы с отрицательной цепью РНК содержат рибонуклеопротеин комплекс, состоящий из генома и RdRp, прикрепленного к каждому сегменту генома, окруженного капсидом.[12] Капсид состоит из белков, складчатая структура которых содержит пять альфа-спиралей в N-концевой доли (мотив 5-H) и трех альфа-спиралей в C-терминал мочка (мотив 3-H). Внутри капсида геном зажат между этими двумя мотивами.[2] Исключая семью Аспивириды, -ssRNA вирусы содержат внешнюю вирусный конверт, тип липид мембрана, окружающая капсид. Форма вирусной частицы, называемой вирионом, -ssRNA вирусов может быть нитевидной, плеоморфной, сферической или трубчатой.[13]
Эволюция
Сегментация генома является характерной чертой многих вирусов -ssRNA, а вирусы -ssRNA варьируются от геномов с одним сегментом, что типично для членов отряда. Mononegavirales, в геномы с десятью сегментами, как в случае Тилапия тилапинвирус.[5][14] Нет четкой тенденции с течением времени, которая определяет количество сегментов, и сегментация генома среди -ssRNA вирусов, по-видимому, является гибким признаком, поскольку она развивалась независимо во многих случаях. Большинство членов подтипа Гапловирикотина несегментированы, тогда как сегментация универсальна в Полипловирикотина.[2][5]
Филогенетика

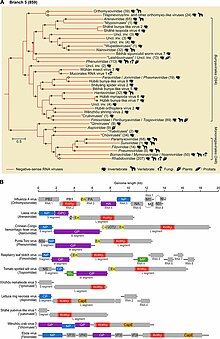
Филогенетический анализ, основанный на RdRp, показывает, что вирусы -ssRNA происходят от общего предка и что они, вероятно, являются сестринской кладой реовирусы, которые представляют собой вирусы дцРНК. Внутри этого типа есть две четкие ветви, относящиеся к двум подтипам, в зависимости от того, синтезирует ли RdRp кэп на вирусной мРНК или хватает кепку от мРНК хозяина и прикрепляет этот кэп к мРНК вируса.[1][3]
В пределах филума -ssRNA вирусы, которые инфицируют членистоногих, по-видимому, являются базальными и являются предками всех других -ssRNA вирусов. Членистоногие часто живут вместе большими группами, что позволяет легко передавать вирусы. Со временем это привело к тому, что -ssRNA-вирусы членистоногих приобрели высокий уровень разнообразия. Хотя членистоногие являются хозяевами большого количества вирусов, существуют разногласия по поводу степени межвидовой передачи вирусов -ssRNA членистоногих среди членистоногих.[4][5]
Вирусы -ssRNA растений и позвоночных обычно генетически связаны с вирусами, инфицированными членистоногими. Более того, большинство -ssRNA вирусов за пределами членистоногих обнаруживается у видов, которые взаимодействуют с членистоногими. Таким образом, членистоногие служат как ключевыми хозяевами, так и векторами передачи вирусов -ssRNA. С точки зрения передачи, -ssRNA-вирусы, не относящиеся к членистоногим, можно разделить на те, которые передаются через членистоногие, и вирусы, которые могут циркулировать среди позвоночных без помощи членистоногих. Вторая группа, вероятно, произошла от первой, приспособившись к передаче только позвоночными.[5]
Классификация
Негарнавирикота принадлежит королевству Орторнавиры, который охватывает все РНК-вирусы, кодирующие RdRp, и область Рибовирия, который включает Орторнавиры а также все вирусы, кодирующие обратная транскриптаза в королевстве Парарнавиры. Негарнавирикота содержит два подтипа, которые объединяют шесть классов, пять из которых являются монотипными вплоть до низших таксонов:[2][9][15]
- Подтип: Гапловирикотина, который содержит вирусы -ssRNA, которые кодируют RdRp, который синтезирует структуру cap на вирусной мРНК и которые обычно имеют несегментированные геномы
- Учебный класс: Chunquiviricetes
- Заказ: Мувиралес
- Семья: Qinviridae
- Род: Иньвирус
- Семья: Qinviridae
- Заказ: Мувиралес
- Учебный класс: Milneviricetes
- Заказ: Serpentovirales
- Семья: Аспвириды
- Род: Офиовирус
- Семья: Аспвириды
- Заказ: Serpentovirales
- Учебный класс: Monjiviricetes
- Учебный класс: Yunchangviricetes
- Заказ: Goujianvirales
- Семья: Yueviridae
- Род: Ююевирус
- Семья: Yueviridae
- Заказ: Goujianvirales
- Учебный класс: Chunquiviricetes
- Подтип: Полипловирикотина, который содержит вирусы -ssRNA, которые кодируют RdRp, который берет кэп с мРНК хозяина для использования в качестве кэпа на вирусной мРНК и которые имеют сегментированные геномы
- Учебный класс: Ellioviricetes
- Заказ: Буньявиралес
- Учебный класс: Insthoviricetes
- Заказ: Articulavirales
- Учебный класс: Ellioviricetes
Вирусы с отрицательной цепью РНК классифицируются как Группа V в Балтиморская классификация система, которая группирует вирусы на основе их способа производства мРНК и которая часто используется вместе со стандартной систематикой вирусов, основанной на истории эволюции. Следовательно, Группа V и Негарнавирикота являются синонимами.[1]
Болезнь
Вирусы с отрицательной цепью РНК вызывают множество широко известных заболеваний. Многие из них передаются членистоногими, включая Вирус лихорадки Рифт-Валли и вирус пятнистого увядания томатов.[16][17] Среди позвоночных летучие мыши и грызуны являются распространенными переносчиками многих вирусов, включая Вирус Эбола и вирус бешенства передается летучими мышами и другими позвоночными животными,[18][19] и Вирус лихорадки Ласса и хантавирусы, передается грызунами.[20][21] Вирусы гриппа распространены среди птиц и млекопитающих.[22] Специфические для человека вирусы -ssRNA включают вирус кори и вирус паротита.[23][24]
История
Многие заболевания, вызываемые -ssRNA-вирусами, известны на протяжении всей истории, включая хантавирусную инфекцию, корь и бешенство.[25][26][27] В современной истории некоторые, такие как Эбола и грипп вызвали вспышки смертельных заболеваний.[28][29] В вирус везикулярного стоматита, впервые выделенный в 1925 году и один из первых вирусов животных, подлежащих изучению, поскольку его можно было хорошо изучить в клеточные культуры, был идентифицирован как вирус -ssРНК, который был уникальным в то время, потому что другие обнаруженные РНК-вирусы имели положительный смысл.[30][31] В начале 21 века болезнь крупного рогатого скота чума крупного рогатого скота, вызванная вирусом -ssRNA чумы крупного рогатого скота, стала второй болезнью, которую необходимо ликвидировать после оспа, вызванный ДНК-вирусом.[32]
В 21 веке вирусная метагеномика стало обычным явлением для выявления вирусов в окружающей среде. Для вирусов -ssRNA это позволило идентифицировать большое количество вирусов беспозвоночных, и особенно членистоногих, что помогло получить представление об эволюционной истории вирусов -ssRNA. На основании филогенетического анализа RdRp, показывающего, что вирусы -ssRNA произошли от общего предка, Негарнавирикота и его два подтипа были созданы в 2018 году, и он был помещен в недавно созданную область Рибовирия.[1][33]
Галерея












Примечания
- ^ В вирус гепатита D часто называют вирусом, но более конкретно его можно описать как вирусоид -подобная патогенная цепь -ssРНК. Исключен из Негарнавирикота потому что, хотя это -ssRNA, она не кодирует RdRp, что является объединяющим признаком вирусов в Орторнавиры.
Рекомендации
- ^ а б c d е ж Wolf Y, Krupovic M, Zhang YZ, Maes P, Dolji V, Koonin EV (21 августа 2017 г.). «Мегатаксономия вирусов с отрицательной РНК» (docx). Международный комитет по таксономии вирусов (ICTV). Получено 6 августа 2020.
- ^ а б c d е Луо М., Террел-младший, Mcmanus SA (30 июля 2020 г.). «Структура нуклеокапсида вируса с отрицательной цепью РНК». вирусы. 12 (8): 835. Дои:10.3390 / v12080835. Получено 6 августа 2020.
- ^ а б Вольф Ю.И., Казлаускас Д., Иранзо Дж., Люсия-Санз А., Кун Дж. Х., Крупович М., Доля В. В., Кунинг Е. В. (27 ноября 2018 г.). «Происхождение и эволюция глобального РНК-вирома». мБио. 9 (6): e02329-18. Дои:10,1128 / мБио.02329-18. ЧВК 6282212. PMID 30482837. Получено 6 августа 2020.
- ^ а б Käfer S, Paraskevopoulou S, Zirkel F, Wieseke N, Donath A, Petersen M, Jones TC, Liu S, Zhou X, Middendorf M, Junglen S, Misof B, Drosten C (12 декабря 2019 г.). «Переоценка разнообразия вирусов с отрицательной цепью РНК у насекомых». PLoS Pathog. 15 (12): e1008224. Дои:10.1371 / journal.ppat.1008224. ЧВК 6932829. PMID 31830128. Получено 6 августа 2020.
- ^ а б c d е Ли СХ, Ши М., Тиан Дж.Х., Линь ХД, Кан Й.Дж., Чен ЛДж, Цинь ХС, Сюй Дж., Холмс Э.К., Чжан Й. «Беспрецедентное геномное разнообразие РНК-вирусов у членистоногих свидетельствует о происхождении РНК-вирусов с отрицательным смыслом». eLife (4): e05378. Дои:10.7554 / eLife.05378. ЧВК 4384744. PMID 25633976. Получено 6 августа 2020.
- ^ «Репликация вируса с отрицательной цепью РНК». ViralZone. Швейцарский институт биоинформатики. Получено 6 августа 2020.
- ^ а б «Транскрипция вируса с отрицательной цепью РНК». ViralZone. Швейцарский институт биоинформатики. Получено 6 августа 2020.
- ^ "Кепка схватки". ViralZone. Швейцарский институт биоинформатики. Получено 6 августа 2020.
- ^ а б Kuhn JH, Wolf YI, Krupovic M, Zhang YZ, Maes P, Dolja VV, Koonin EV (февраль 2019 г.). «Классифицируйте вирусы - выгода стоит затраченных усилий». Природа. 566 (7744): 318–320. Дои:10.1038 / d41586-019-00599-8. PMID 30787460. Получено 6 августа 2020.
- ^ «Заикание полимеразы вируса с отрицательной цепью РНК». ViralZone. Швейцарский институт биоинформатики. Получено 6 августа 2020.
- ^ «Транскрипция Ambisense в вирусах с отрицательной цепью РНК». ViralZone. Швейцарский институт биоинформатики. Получено 6 августа 2020.
- ^ Чжоу Х, Сунь И, Го И, Лу З (сентябрь 2013 г.). «Структурная перспектива образования рибонуклеопротеинового комплекса в одноцепочечных РНК-вирусах с отрицательным смыслом». Тенденции Microbiol. 21 (9): 475–484. Дои:10.1016 / j.tim.2013.07.006. PMID 23953596.
- ^ Фермин, Г. (2018). Вирусы: молекулярная биология, взаимодействие с хозяевами и применение в биотехнологии. Эльзевир. п. 19-27, 43. Дои:10.1016 / B978-0-12-811257-1.00002-4. ISBN 9780128112571.
- ^ Бахарах Э., Мишра Н., Бриз Т., Зоди М.С., Кембу Цофак Дж. Э., Замостиано Р., Берковиц А., Нг Дж., Нитидо А., Корвело А., Туссен, Северная Каролина, Абель Нильсен СК, Хорниг М., Дель Посо Дж, Блум Т., Фергюсон Х , Эльдар А., Липкин В.И. (5 апреля 2016 г.). «Характеристика нового ортомиксоподобного вируса, вызывающего массовое отмирание тилапии». мБио. 7 (2): e00431-16. Дои:10,1128 / мBio.00431-16. ЧВК 4959514. PMID 27048802. Получено 6 августа 2020.
- ^ «Таксономия вирусов: выпуск 2019 г.». talk.ictvonline.org. Международный комитет по таксономии вирусов. Получено 6 августа 2020.
- ^ Хартман А. (июнь 2017 г.). "Лихорадка Рифт-Валли". Clin Lab Med. 37 (2): 285–301. Дои:10.1016 / j.cll.2017.01.004. ЧВК 5458783. PMID 28457351.
- ^ Scholthof KB, Adkins S, Czosnek H, Palukaitis P, Jacquot E, Hohn T., Hohn B, Saunders K, Candresse T., Ahlquist P, Hemenway C, Foster GD (декабрь 2011 г.). «Топ-10 вирусов растений в молекулярной патологии растений». Мол Растение Патол. 12 (9): 938–954. Дои:10.1111 / j.1364-3703.2011.00752.x. ЧВК 6640423. PMID 22017770. Получено 6 августа 2020.
- ^ Муньос-Фонтела С, МакЭлрой АК (2017). «Болезнь, вызванная вирусом Эбола, у людей: патофизиология и иммунитет». Curr Top Microbiol Immunol. 411: 141–169. Дои:10.1007/82_2017_11. ЧВК 7122202. PMID 28653186. Получено 6 августа 2020.
- ^ Фишер CR, Streicker DG, Schnell MJ (апрель 2018 г.). «Распространение и эволюция вируса бешенства: покорение новых рубежей». Нат Рев Микробиол. 16 (4): 241–255. Дои:10.1038 / nrmicro.2018.11. ЧВК 6899062. PMID 29479072.
- ^ Юн NE, Уокер DH (9 октября 2012 г.). «Патогенез лихорадки Ласса». Вирусы. 4 (10): 2031–2048. Дои:10.3390 / v4102031. ЧВК 3497040. PMID 23202452. Получено 6 августа 2020.
- ^ Авшич-Жупанч Т, Саксида А, Корва М (апрель 2019). «Хантавирусные инфекции». Clin Microbiol Инфекция. 21S: e6 – e16. Дои:10.1111/1469-0691.12291. PMID 24750436. Получено 6 августа 2020.
- ^ Боркенхаген Л.К., Салман М.Д., Ма М.Дж., Грей Г.К. (ноябрь 2019 г.). «Инфекции вируса гриппа животных у людей: комментарий». Int J Infect Dis. 88: 113–119. Дои:10.1016 / j.ijid.2019.08.002. PMID 31401200. Получено 6 августа 2020.
- ^ «Передача кори». cdc.gov. Центры по контролю и профилактике заболеваний (CDC). 5 февраля 2018 г.. Получено 6 августа 2020.
- ^ Рубин С., Экхаус М., Ренник Л.Дж., Бэмфорд К.Г., Дюпрекс В.П. (январь 2015 г.). «Молекулярная биология, патогенез и патология вируса паротита». Джей Патол. 235 (2): 242–252. Дои:10.1002 / путь.4445. ЧВК 4268314. PMID 25229387. Получено 6 августа 2020.
- ^ Цзян Х, Чжэн Х, Ван Л., Ду Х, Ван П, Бай Х (2017). «Хантавирусная инфекция: глобальная зоонозная проблема». Вирол Син. 32 (1): 32–43. Дои:10.1007 / с12250-016-3899-х. ЧВК 6598904. PMID 28120221.
- ^ «История кори». cdc.gov. Центры по контролю и профилактике заболеваний (CDC). 5 февраля 2018 г.. Получено 6 августа 2020.
- ^ Веласко-Вилла А, Маулдин М.Р., Ши М., Эскобар Л.Е., Галлардо-Ромеро Н.Ф., Дэймон И., Олсон В.А., Штрейкер Д.Г., Эмерсон Г. (октябрь 2017 г.). «История бешенства в Западном полушарии». Противовирусный Res. 146: 221–232. Дои:10.1016 / j.antiviral.2017.03.013. ЧВК 5620125. PMID 28365457. Получено 6 августа 2020.
- ^ Завилинская Б, Кош-Вненчак М (2014). «Общее введение в биологию и болезнь вируса Эбола» (PDF). Folia Med Cracov. 54 (3): 57–65. PMID 25694096. Получено 6 августа 2020.
- ^ Krammer F, Smith G, Fouchier R, Peiris M, Kedzierska K, Doherty PC, Palese P, Shaw ML, Treanor J, Webster RG, García-Sastre A (28 июня 2018 г.). «Грипп». Праймеры Nat Rev Dis. 4 (1): 3. Дои:10.1038 / с41572-018-0002-у. ЧВК 7097467. PMID 29955068. Получено 6 августа 2020.
- ^ «Вирус везикулярного стоматита» (PDF). Информационный центр здоровья свиней. Центр продовольственной безопасности и общественного здравоохранения, Колледж ветеринарной медицины, Университет штата Айова. Ноябрь 2015. Получено 6 августа 2020.
- ^ Колаковский Д. (апрель 2015 г.). «Краткая предвзятая история РНК-вирусов». РНК. 21 (4): 667–669. Дои:10.1261 / rna.049916.115. ЧВК 4371325. PMID 25780183. Получено 6 августа 2020.
- ^ Гринвуд B (12 мая 2014 г.). «Вклад вакцинации в глобальное здоровье: прошлое, настоящее и будущее». Philos Trans R Soc Lond B Biol Sci. 369 (1645): 20130433. Дои:10.1098 / rstb.2013.0433. ЧВК 4024226. PMID 24821919. Получено 6 августа 2020.
- ^ «История таксономии ICTV: Negarnaviricota». Международный комитет по таксономии вирусов (ICTV). Получено 6 августа 2020.
дальнейшее чтение
- Уорд, К. У. (1993). «Прогресс в направлении более высокой таксономии вирусов». Исследования в вирусологии. 144 (6): 419–53. Дои:10.1016 / S0923-2516 (06) 80059-2. ЧВК 7135741. PMID 8140287.