Коэффициент Редфилда - Redfield ratio
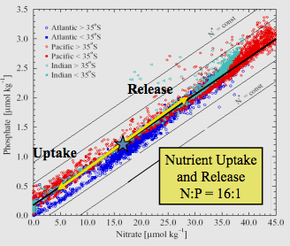
Коэффициент Редфилда или же Стехиометрия Редфилда постоянное атомное соотношение углерод, азот и фосфор найдено в морском фитопланктон и повсюду в глубоких океанах.
Термин назван в честь американского океанограф Альфред С. Редфилд который в 1934 году впервые описал относительно постоянное соотношение питательных веществ в образцах морской биомассы, собранных во время нескольких рейсов на борту исследовательского судна. Атлантида, и эмпирически установлено, что соотношение составляет C: N: P = 106: 16: 1.[1] Хотя отклонения от канонического соотношения 106: 16: 1 были обнаружены в зависимости от видов фитопланктона и области исследования, соотношение Редфилда остается важным ориентиром для океанографов, изучающих ограничение питательных веществ. В документе 2014 года, обобщающем большой набор данных измерений питательных веществ во всех основных регионах океана за период с 1970 по 2010 год, сообщается, что глобальное медианное значение C: N: P составляет 163: 22: 1.[2]
Открытие
| Часть серии о |
| Цикл углерода |
|---|
 |
В своей статье 1934 года Альфред Редфилд проанализировал нитрат и фосфат данные для Атлантический, Индийский, Тихий океан океаны и Баренцево море.[1] Как Гарвард физиолог, Редфилд участвовал в нескольких рейсах на борту исследовательского судна. Атлантида, анализируя данные о содержании C, N и P в морском планктоне, и ссылаясь на данные, собранные другими исследователями еще в 1898 году.
Анализ эмпирических данных, проведенный Редфилдом, позволил ему обнаружить, что во всех трех океанах и в Баренцевом море атомное соотношение N: P в морской воде составляет около 20: 1 (позже исправлено до 16: 1), и оно очень похоже на среднее значение. N: P фитопланктона.
Для объяснения этого явления Редфилд первоначально предложил два взаимоисключающих механизма:
I) N: P в планктоне имеет тенденцию к N: P составу морской воды. В частности, виды фитопланктона с разными потребностями в азоте и фосфоре конкурируют в одной и той же среде и отражают питательный состав морской воды.[1]
II) Равновесие между морской водой и пулами питательных веществ планктона поддерживается за счет механизмов биотической обратной связи.[1] [3] Редфилд предложил сценарий, подобный термостату, в котором действия фиксаторов азота и денитрификаторов поддерживают соотношение нитратов и фосфатов в морской воде, близкое к требованиям протоплазмы.[4] Учитывая, что в то время было мало что известно о составе «протоплазмы» или основном составе фитопланктона, Редфилд не пытался объяснить, почему его соотношение N: P должно быть примерно 16: 1.
В 1958 году, спустя почти четверть века после первого открытия соотношений, Редфилд склоняется к последнему механизму в своей рукописи «Биологический контроль химических факторов в окружающей среде».[3] Редфилд предположил, что соотношение азота и фосфора в планктоне привело к тому, что глобальный океан имел удивительно похожее соотношение растворенных веществ. нитрат к фосфат (16: 1). Он подумал, как циклы не только N и P, но также C и O могут взаимодействовать, чтобы привести к этому совпадению.
Объяснение
Редфилд обнаружил замечательное соответствие между химией глубинного океана и химией живых существ, таких как фитопланктон на поверхности океана. Оба имеют отношение N: P примерно 16: 1 в пересчете на атомы. Когда питательных веществ нет ограничение, то коренной зуб соотношение элементов C: N: P в большинстве фитопланктона составляет 106: 16: 1. Редфилд считал, что химия бескрайних океанов, идеально отвечающая требованиям живых организмов, не является чистой случайностью.
Лабораторные эксперименты в контролируемых химических условиях показали, что биомасса фитопланктона будет соответствовать коэффициенту Редфилда, даже когда уровни питательных веществ в окружающей среде превышают их, что позволяет предположить, что экологическая адаптация к соотношению питательных веществ в океане не является единственным регулирующим механизмом (вопреки одному из механизмов, первоначально предложенных Редфилдом. ). [5] Однако последующее моделирование механизмов обратной связи, в частности потоков нитрат-фосфорного сцепления, действительно поддерживает предложенный им механизм биотического равновесия с обратной связью, хотя эти результаты противоречат ограничениям в нашем нынешнем понимании потоков питательных веществ. [6]
В океане большая часть биомассы состоит из богатого азотом планктона. Многие из этого планктона потребляются другой биомассой планктона, имеющей аналогичный химический состав. Это приводит к аналогичному соотношению азота и фосфора в среднем для всего планктона во всем Мировом океане, которое, как было установлено эмпирическим путем, в среднем составляет примерно 16: 1. Когда эти организмы опускаются в глубь океана, их биомасса потребляется бактериями, которые в аэробный условия, окислять органическое вещество для образования растворенных неорганических питательных веществ, в основном углекислый газ, нитраты и фосфаты.
То, что соотношение нитратов и фосфатов внутри всех основных океанических бассейнов очень похоже, возможно, из-за времени пребывания этих элементов в океане по сравнению со временем циркуляции океанов, примерно 100000 лет для фосфора и 2000 лет для азота. .[7] Тот факт, что времена пребывания этих элементов больше, чем времена перемешивания океанов (~ 1000 лет)[8] может привести к тому, что соотношение нитратов и фосфатов во внутренней части океана останется довольно однородным.
Хотя такие аргументы могут потенциально объяснить, почему отношения довольно постоянны, они не решают вопрос, почему отношение N: P составляет почти 16, а не какое-то другое число.
Использует
Исследования, которые привели к этому соотношению, стали фундаментальной особенностью в понимании биогеохимических циклов океанов и одним из ключевых принципов биогеохимии. Коэффициент Редфилда играет важную роль в оценке потоков углерода и биогенных веществ в модели глобальной циркуляции. Они также помогают определить, какие питательные вещества ограничивают локализованную систему, если есть ограничивающие питательные вещества. Это соотношение также можно использовать для понимания формирования цветения фитопланктона и, как следствие, гипоксии, путем сравнения соотношения между различными регионами, например, сравнения соотношения Редфилда реки Миссисипи с соотношением северной части Мексиканского залива.[9] Контроль N: P может быть средством устойчивого управления резервуаром.[10]
Отклонения от канонического соотношения Редфилда
Коэффициент Редфилда первоначально был получен эмпирическим путем из измерений элементного состава планктона в дополнение к содержанию нитратов и фосфатов в морской воде, собранной на нескольких станциях в Атлантический океан. Позже это было подтверждено сотнями независимых измерений. Однако, глядя на состав отдельных разновидность фитопланктона, выращенного в условиях ограничения азота или фосфора, показывает, что это отношение азота к фосфору может варьироваться от 6: 1 до 60: 1. Понимая эту проблему, Редфилд никогда не пытался ее объяснить, за исключением того, что отмечал, что отношение N: P в неорганических питательных веществах во внутренней части океана было средним с ожидаемой незначительной изменчивостью.
Хотя соотношение Редфилда удивительно стабильно в глубоководных районах океана, фитопланктон может иметь большие вариации в составе C: N: P, и их жизненная стратегия играет роль в соотношении C: N: P, что заставило некоторых исследователей предположить, что Коэффициент Редфилда, возможно, является скорее общим средним, чем конкретным требованием для роста фитопланктона.[11] Однако недавно было обнаружено, что соотношение Редфилда связано с гомеостатическим отношением белка крРНК ratio в основном присутствует как у прокариот, так и у эукариот.[12] Кроме того, было показано, что коэффициент Редфилда варьируется в разных пространственных масштабах, а также в среднем немного выше (166: 20: 1), чем исходная оценка Редфилда.[13] Было также показано, что в некоторых экосистемах коэффициент Редфилда значительно варьируется в зависимости от доминирующих таксонов фитопланктона, присутствующих в экосистеме, даже в системах с обильными питательными веществами. Следовательно, коэффициент Редфилда для конкретной системы может служить представителем структуры планктонного сообщества. [14]
Несмотря на сообщения о том, что элементный состав организмы например, морской фитопланктон в океаническом регионе не соответствует каноническому коэффициенту Редфилда, фундаментальная концепция этого отношения остается актуальной и полезной. В 2014 году в журнале Scientific Data была опубликована статья, в которой были агрегированы измерения коэффициентов Редфилда из наблюдательных круизов по всему миру с 1970 по 2010 год. Эта статья представляет собой большую базу данных, которую можно использовать для изучения эволюции определенных фосфора, углерода и азота. по морским станциям и времени.[2]
Расширенный коэффициент Редфилда
Некоторые считают, что есть и другие элементы, например калий, сера, цинк, медь, и утюг которые также важны в химия океана.[15]
В частности, железо (Fe) считалось очень важным еще в биологические океанографы предположил, что железо также может быть ограничивающий фактор за основное производство В океане.[16] В результате был разработан расширенный коэффициент Редфилда, чтобы включить его в этот баланс. Этот новый стехиометрическое соотношение заявляет, что соотношение должно быть 106 C: 16 N: 1 P: 0,1-0,001 Fe. Большой разброс по Fe является результатом значительных препятствий со стороны судов и научного оборудования, загрязняющих любые образцы, собранные в море, с избытком Fe.[17]. Это было загрязнение в результате были получены первые свидетельства того, что концентрация железа была высокой и не являлась ограничивающим фактором для первичной продукции в море.
Диатомеи потребность, среди других питательных веществ, кремниевая кислота создавать биогенный кремнезем для них панцири (клеточные стенки). В результате этого для диатомовых водорослей было предложено соотношение питательных веществ Редфилда-Бжезинского и составило C: Si: N: P = 106: 15: 16: 1.[18] Выходя за рамки самого первичного производства, кислород, потребляемый аэробным дыханием биомассы фитопланктона, также, как было показано, следует предсказуемой пропорции по отношению к другим элементам. О2Соотношение: C составляет 138: 106. [6]
Смотрите также
Рекомендации
- ^ а б c d Редфилд, AC (1934). «О соотношении органических производных в морской воде и их связи с составом планктона» (PDF). Том Мемориала Джеймса Джонстона: 176. Получено 1 марта, 2019.
- ^ а б Мартини, Адам С .; Vrugt, Jasper A .; Ломас, Майкл В. (9 декабря 2014 г.). «Концентрации и соотношения твердых частиц органического углерода, азота и фосфора в Мировом океане». Научные данные. 1 (1): 140048. Дои:10.1038 / sdata.2014.48. ЧВК 4421931. PMID 25977799.
- ^ а б РЕДФИЛД, АЛЬФРЕД К. (1958). «Биологический контроль химических факторов в окружающей среде». Американский ученый. 46 (3): 205–221, 230A. ISSN 0003-0996. JSTOR 27827150. PMID 24545739.
- ^ Грубер, Николас; Сармьенто, Хорхе Л. (1997). «Глобальные закономерности морской азотфиксации и денитрификации». Глобальные биогеохимические циклы. 11 (2): 235–266. Bibcode:1997GBioC..11..235G. Дои:10.1029 / 97GB00077. ISSN 1944-9224.
- ^ Гольдман, Джоэл С. (1986). «О темпах роста фитопланктона и соотношении C: N: P в твердых частицах при слабом освещении1». Лимнология и океанография. 31 (6): 1358–1363. Bibcode:1986LimOc..31.1358G. Дои:10.4319 / lo.1986.31.6.1358. ISSN 1939-5590.
- ^ а б Лентон, Тимоти М .; Уотсон, Эндрю Дж. (2000). «Еще раз о Редфилде: 1. Регулирование нитратов, фосфатов и кислорода в океане». Глобальные биогеохимические циклы. 14 (1): 225–248. Bibcode:2000GBioC..14..225L. Дои:10.1029 / 1999GB900065. ISSN 1944-9224.
- ^ http://lgmacweb.env.uea.ac.uk/green_ocean/positions/diazotroph.shtml
- ^ «Химическая сенсорная группа». 2015-08-20.
- ^ Доддс, Уолтер (2006). «Питательные вещества и« мертвая зона »: связь между соотношением питательных веществ и растворенным кислородом в Северном Мексиканском заливе». Границы экологии и окружающей среды. 4 (4): 211–217. Дои:10.1890 / 1540-9295 (2006) 004 [0211: NATDZT] 2.0.CO; 2.
- ^ Левич, А. П. (1 февраля 1996 г.). «Роль соотношения азота и фосфора в отборе цианобактерий или зеленых водорослей для доминирования фитопланктона и его применение в управлении водохранилищами». Журнал здоровья водных экосистем. 5 (1): 55–61. Дои:10.1007 / BF00691729. ISSN 1573-5141.
- ^ Арриго, КР (2005). «Морские микроорганизмы и глобальный круговорот питательных веществ». Природа. 437 (7057): 349–55. Bibcode:2005Натура.437..349А. Дои:10.1038 / природа04159. PMID 16163345.
- ^ Лоладзе Ираклий; Эльзер, Джеймс Дж. (2011). «Происхождение соотношения азота и фосфора Редфилда лежит в гомеостатическом соотношении белка и рРНК». Письма об экологии. 14 (3): 244–250. Дои:10.1111 / j.1461-0248.2010.01577.x. ISSN 1461-0248. PMID 21244593.
- ^ Стернер, Роберт В .; Андерсен, Том; Эльзер, Джеймс Дж .; Hessen, Dag O .; Худ, Джеймс М .; Макколи, Эдвард; Урабе, Джотаро (2008). «Масштабно-зависимая стехиометрия сестона углерода: азота: фосфора в морской и пресной воде». Лимнология и океанография. 53 (3): 1169–1180. Bibcode:2008LimOc..53.1169S. Дои:10.4319 / lo.2008.53.3.1169. ISSN 1939-5590.
- ^ Арриго, Кевин Р .; Данбар, Роберт Б .; Lizotte, Michael P .; Робинсон, Д. Х. (2002). «Специфические для таксонов различия в усадке C / P и N / P для фитопланктона в море Росса, Антарктида». Письма о геофизических исследованиях. 29 (19): 44–1–44-4. Bibcode:2002GeoRL..29.1938A. Дои:10.1029 / 2002GL015277. ISSN 1944-8007.
- ^ Беннер, Р. Пакульски, JD; Маккарти, М; Hedges, JI; Хэтчер, PG (1992). «Объемные химические характеристики растворенного органического вещества в океане». Наука. 255 (5051): 1561–4. Bibcode:1992Научный ... 255.1561B. Дои:10.1126 / science.255.5051.1561. PMID 17820170.
- ^ Tortell, PD; Мальдонадо, штат Монтана; Грейнджер, Дж; Прайс, Н. М. (1999). «Морские бактерии и биогеохимический круговорот железа в океанах». Микробиология Экология. 29 (1): 1. Дои:10.1111 / j.1574-6941.1999.tb00593.x.
- ^ Broecker, WS; Пэн, Т; Бенг, З. (1982). Трассеры в море. Геологическая обсерватория Ламонт-Доэрти.
- ^ Бжезинский, Марк А. (1985). «СООТНОШЕНИЕ Si: C: N МОРСКИХ ДИАТОМОВ: МЕЖСПЕЦИАЛЬНАЯ ИЗМЕНЧИВОСТЬ И ВЛИЯНИЕ НЕКОТОРЫХ ЭКОЛОГИЧЕСКИХ ПЕРЕМЕННЫХ1». Журнал психологии. 21 (3): 347–357. Дои:10.1111 / j.0022-3646.1985.00347.x. ISSN 1529-8817.
- Джонсон, Закари. «Биогеохимия IV». Школа наук и технологий об океане и Земле Гавайского университета. Интернет. <http://www.soest.hawaii.edu/oceanography/zij/ocn621/OCN621-20060215-biogeochemistry.pdf >.
- Ленц, Дженнифер. «Стехиометрия питательных веществ - соотношения Редфилда». Школа побережья и окружающей среды LSU, 2010. Web. <http://www.sce.lsu.edu/cego/Documents/Reviews/Oceanography/Nutrient_Stoichiometry.pdf >.
- П.Г. Фальковски и К.С. Дэвис. «МОРСКАЯ БИОГЕОХИМИЯ: ОТНОШЕНИЯ РЕДФИЛДА». ScienceWeek. Природа, 2004. Интернет. <http://scienceweek.com/2004/sa041119-5.htm >.