Клеточная механика - Cell mechanics
Клеточная механика является подполе биофизика который фокусируется на механических свойствах и поведении живых клеток, а также на том, как это связано с функцией клеток.[1] Он охватывает аспекты биофизика клетки, биомеханика, физика мягкой материи и реология, механобиология и клеточная биология.
Механика эукариотических клеток
Эукариотические клетки [2] клетки, которые состоят из мембраносвязанных органеллы мембраносвязанный ядро, и более одного линейного хромосома. Быть намного сложнее, чем прокариотический клетки, клетки без настоящего ядра, эукариоты должны защищать свои органеллы от внешних сил.
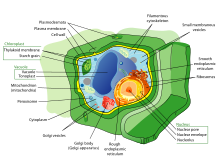
Механика растительных клеток
Механика растительной клетки объединяет принципы биомеханика и механобиология исследовать рост и формирование растительных клеток. Клетки растений, как и клетки животных, реагируют на внешние силы, например, реорганизацией своей цитоскелетной сети. Наличие значительно жесткого внеклеточный матрикс, то клеточная стенка однако наделяет клетки растений рядом определенных свойств. В основном рост растительных клеток контролируется механикой и химическим составом клеточной стенки.[3] Основная часть исследований в области механики растительных клеток направлена на измерение и моделирование механики клеточной стенки, чтобы понять, как изменение ее состава и механических свойств влияет на функцию, рост и рост клеток. морфогенез.[4][5]
Механика клеток животных

Потому что клетки животных[6] не имеют клеточных стенок, чтобы защитить их, как клетки растений, им требуются другие специализированные структуры, чтобы выдерживать внешние механические силы. Все клетки животных заключены в клеточную мембрану, состоящую из тонкого липидного бислоя, который защищает клетку от воздействия внешней среды. Используя рецепторы, состоящие из белковых структур, клеточная мембрана может впускать выбранные молекулы внутрь клетки. Внутри клеточной мембраны находится цитоплазма, в котором находится цитоскелет.[7] Сеть нитчатых белков, включая микротрубочки, промежуточные нити, и актиновые нити составляет цитоскелет и помогает поддерживать форму клетки. Работая вместе, три типа полимеров могут организовать себя, чтобы противостоять приложенным внешним силам и сопротивляться деформации. Однако между тремя полимерами есть различия.
Основной структурный компонент цитоскелет актиновые филаменты. Самый узкий (диаметр 7 нм) и самый гибкий из трех типов полимеры актиновые филаменты обычно находятся на самом краю цитоплазмы в клетках животных.[8] Образуется путем связывания полимеров белка, называемого актин, они помогают придавать клеткам форму и структуру, а также способны транспортировать белковые пакеты и органеллы. Более того, актиновые филаменты обладают способностью быстро собираться и разбираться, что позволяет им участвовать в подвижности клеток.[9]
С другой стороны, промежуточные волокна представляют собой более постоянные структуры диаметром от 8 до 10 нм.[10] Состоящие из многочисленных волокнистых белковых нитей, намотанных вместе, основная роль промежуточных белков заключается в том, чтобы нести напряжение и сохранять форму и структуру клетки, закрепляя ядро и другие органеллы в предназначенных для них областях.
Самая крупная цитоскелетная структура из трех типов полимеров - это микротрубочки диаметром 25 нм.[9] В отличие от актиновых филаментов, микротрубочки представляют собой жесткие полые структуры, исходящие наружу от центр организации микротрубочек (MTOC). Состоящие из белков тубулина микротрубочки представляют собой динамические структуры, которые позволяют им сокращаться или расти при добавлении или удалении белков тубулина. С точки зрения клеточной механики, основная цель микротрубочек - противостоять сжимающим клеточным силам и действовать как транспортная система для моторных белков.[9]
Механика измерительной ячейки
Поскольку клетки представляют собой крошечные мягкие объекты, которые необходимо измерять иначе, чем такие материалы, как металл, пластик и стекло, были разработаны новые методы точного измерения механики клеток. Разнообразие техник можно разделить на две категории: методы приложения силы и методы измерения силы.[8]В случае клеток с стенками, таких как клетки растений или грибов, из-за наличия жесткой, анизотропной и изогнутой клеточной стенки, инкапсулирующей клетки, могут потребоваться особые соображения и индивидуальные подходы по сравнению с методами, используемыми для измерения механики клеток животных. [11]
Применение силы
Методы приложения силы используют реакцию ячейки деформации на силу, приложенную к ячейке, как способ измерения механических свойств ячейки.[12] Существует несколько различных методов приложения силы, в том числе:
- Для аспирации микропипеткой используется прикладываемое давление всасывания с помощью стеклянной пипетки малого диаметра. Измерение длины аспирации, вызванной давлением всасывания, может выявить несколько механических свойств ячейки.[13]
- Управление кантилевером осуществляется посредством магнитного, электрического или механического взаимодействия между зондом и поверхностью ячейки, которое дает сигнал, который можно использовать для измерения механических свойств.[14]
- Оптические методы включают использование захваченных фотонов для манипулирования клетками. В фотоны будет меняться в направлении в зависимости от показателя преломления ячейки, что вызовет изменение количества движения, что приведет к силе, приложенной к ячейке.[12]
- Механические методы используют введение ферромагнитных шариков в клетку или прикрепление к определенным рецепторам на клетке. При приложении магнитной силы растяжение мембраны можно измерить для расчета механических свойств.[12]
- Деформация субстрата измеряет эластичность посредством растяжения клетки. В эластичность клетки предоставляет информацию, которая может определять подвижность и адгезию.[12][15]
- Сжатие требует приложения давления на всю ячейку. Вычисляя изменения формы клетки, сжатие позволяет измерить механическую реакцию на силу.[12]
- Техника потока использует Число Рейнольдса, безразмерное число в механике жидкости, чтобы различить, подвержена ли ячейка ламинарному, переходному или турбулентному потоку.[12]
Определение силы
- Сморщивание мембран требует помещения клетки в гибкую силиконовую оболочку. По мере того как клетка сокращается, величину сил можно оценить, используя длину и количество морщин.[12]
- Тяговая силовая микроскопия обнаруживает деформации путем сравнения изображений движения флуоресцентных шариков, которые были прикреплены к клетке.[16]
- Зондирование кантилевера может обнаруживать поверхностные напряжения с прикреплением микромеханических лучей на одном конце ячейки.[17]
- Биореакторы позволяют измерять многоклеточные силы в трехмерной системе при одновременном приложении внешних сил. Это позволяет получать лучшие результаты и более точные данные сложных экспериментов.[12]
- Когда прилипшие клетки возбуждаются поверхностными акустическими волнами, они начинают генерировать акустический микропоток. Величина скорости этого потока около клеточной мембраны прямо пропорциональна жесткости (то есть модулю упругости) клетки.[18]
Исследование
Исследователи, изучающие клеточную механику, интересуются механикой и динамикой сборок и структур, из которых состоит клетка, включая мембраны, цитоскелет, органеллы, и цитоплазма, и как они взаимодействуют, чтобы вызвать эмерджентные свойства клетки в целом.[19]
Особое внимание во многих исследованиях клеточной механики уделялось цитоскелет, который в клетки животных ) можно подумать, что он состоит из:
- актомиозиновые сборки (F-актин, миозин моторы и связанные с ними связывающие, зародышевые, кэпирующие, стабилизирующие и сшивающие белки),
- микротрубочки и связанные с ними моторные белки (кинезины и динеины ),
- промежуточные нити,
- другие сборки, такие как спектрины и септины.
Активные неравновесные и нелинейные реологические свойства клеточных ансамблей стали предметом пристального внимания исследований в последнее время.[20][21] Еще один интересный момент: как клеточный цикл -связанные изменения в цитоскелет активность влияет на глобальные свойства клеток, такие как повышение внутриклеточного давления во время округление митотических клеток.[22]
Рекомендации
- ^ Моендарбари, Эмад; Харрис, Эндрю (2014). «Клеточная механика: принципы, практика, перспективы». Междисциплинарные обзоры Wiley: системная биология и медицина. 6: 371–388. Дои:10.1002 / wsbm.1275. ЧВК 4309479. PMID 25269160.
- ^ «Введение в эукариотические клетки». Ханская академия.
- ^ Бидхенди, Амир Дж; Альтартури, Бара; Gosselin, Frédérick P .; Гайтманн, Аня (июль 2019 г.). «Механический стресс инициирует и поддерживает морфогенез эпидермальных клеток волнистых листьев». Отчеты по ячейкам. 28 (5): 1237–1250. Дои:10.1016 / j.celrep.2019.07.006. PMID 31365867.
- ^ Бидхенди, Амир Дж; Гайтманн, Аня (январь 2016 г.). «Связь механики первичной клеточной стенки растений с морфогенезом» (PDF). Журнал экспериментальной ботаники. 67 (2): 449–461. Дои:10.1093 / jxb / erv535. PMID 26689854.
- ^ Бидхенди, Амир Дж; Гайтманн, Аня (январь 2018 г.). «Конечно-элементное моделирование изменения формы растительных клеток» (PDF). Физиология растений. 176 (1): 41–56. Дои:10.1104 / стр.17.01684. ЧВК 5761827. PMID 29229695.
- ^ МакГрегор, Джессика (6 августа 2018 г.). "Части клетки животного". Научные тенденции. Дои:10.31988 / SciTrends.24128.
- ^ Кларк, Эндрю Дж .; Wartlick, Ortrud; Сальбре, Гийом; Палуч, Ева К. (май 2014 г.). «Стрессы на поверхности клетки во время морфогенеза клеток животных». Текущая биология. 24 (10): R484 – R494. Дои:10.1016 / j.cub.2014.03.059. PMID 24845681.
- ^ а б Моендарбари, Эмад; Харрис, Эндрю Р. (NaN). «Клеточная механика: принципы, практика, перспективы». Междисциплинарные обзоры Wiley: системная биология и медицина. 6 (5): 371–388. Дои:10.1002 / wsbm.1275. ЧВК 4309479. PMID 25269160. Проверить значения даты в:
| дата =(помощь) - ^ а б c «Микротрубочки и нити». Scitable by Nature Education.
- ^ «Что такое промежуточные нити? | MBInfo». www.mechanobio.info.
- ^ Бидхенди, Амир Дж; Гайтманн, Аня (июль 2019 г.). «Методы количественной оценки механики первичной клеточной стенки растений» (PDF). Журнал экспериментальной ботаники. 70 (14): 3615–3648. Дои:10.1093 / jxb / erz281. PMID 31301141.
- ^ а б c d е ж грамм час Родригес, Марита Л .; МакГарри, Патрик Дж .; Сниадецки, Натан Дж. (15 октября 2013 г.). «Обзор клеточной механики: экспериментальный и модельный подходы». Обзоры прикладной механики. 65 (6): 060801–060801–41. Дои:10.1115/1.4025355.
- ^ «Лекция 17: Клеточная механика» (PDF).
- ^ Джалили, Надер (10 ноября 2012 г.). «Наномеханическая манипуляция на основе кантилевера для зондирования и визуализации». Наноробототехника. Springer Нью-Йорк. С. 29–40. Дои:10.1007/978-1-4614-2119-1_2. ISBN 978-1-4614-2118-4.
- ^ Гассеми, Саба; Меаччи, Джованни; Лю, Шуайминь; Гондаренко, Александр А .; Матур, Анураг; Рока-Кусакс, Пере; Sheetz, Майкл П .; Хоун, Джеймс (2012-04-03). «Клетки проверяют жесткость субстрата путем локального сжатия на субмикрометровых столбах». Труды Национальной академии наук. 109 (14): 5328–5333. Дои:10.1073 / pnas.1119886109. ISSN 0027-8424. ЧВК 3325713. PMID 22431603.
- ^ Плотников, Сергей В .; Сабасс, Бенедикт; Schwarz, Ulrich S .; Уотерман, Клэр М. (2014). Тяговая силовая микроскопия высокого разрешения. Методы клеточной биологии. 123. С. 367–394. Дои:10.1016 / B978-0-12-420138-5.00020-3. ISBN 9780124201385. ЧВК 4699589. PMID 24974038.
- ^ Датар, Рам; Ким, Сонхван; Чон, Санмин; Хескет, Питер; Маналис, Скотт; Бойзен, Аня; Thundat, Томас. «Консольные датчики: наномеханические инструменты диагностики» (PDF).
- ^ Salari, A .; Аппак-Баской, С .; Ezzo, M .; Hinz, B .; Колиос, M.C .; Цай, С.С.Х. (2019) Танцы с клетками: акустические микропотоки, создаваемые колеблющимися клетками. https://doi.org/10.1002/smll.201903788
- ^ Флетчер, Дэниел А; Маллинз, Дайч (28 января 2010 г.). «Клеточная механика и цитоскелет». Природа. 463 (7280): 485–492. Дои:10.1038 / природа08908. ЧВК 2851742. PMID 20110992.
- ^ Мизуно, Дайсуке; Тардин, Екатерина; Шмидт, Кристоф Ф.; МакКинтош, Фред С. (19 января 2007 г.). «Неравновесная механика активных цитоскелетных сетей». Наука. 315 (5810): 370–373. Дои:10.1126 / science.1134404. PMID 17234946.
- ^ Го, Мин; Эрлихер, Аллен Дж; Jensen, Mikkel H; Ренц, Мальте; Мур, Джеффри Р.; Гольдман, Роберт Д; Липпинкотт-Шварц, Дженнифер; Макинтош, Фред Си; Вайц, Дэвид А. (14 августа 2014 г.). «Исследование стохастических, двигательных свойств цитоплазмы с помощью микроскопии силового спектра». Клетка. 158 (4): 822–832. Дои:10.1016 / j.cell.2014.06.051. ЧВК 4183065. PMID 25126787.
- ^ Стюарт, Мартин П.; Хелениус, Йонне; Тойода, Юске; Раманатан, Субраманианский P; Мюллер, Дэниел Дж; Хайман, Энтони А. (2 января 2011 г.). «Гидростатическое давление и актомиозиновая кора приводят к округлению митотических клеток». Природа. 469 (7329): 226–230. Дои:10.1038 / природа09642. PMID 21196934.