Круговая РНК - Circular RNA
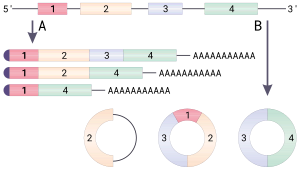
Круговая РНК (или же circRNA) представляет собой одноцепочечный РНК который, в отличие от линейной РНК, образует ковалентно замкнутый непрерывный цикл. В кольцевой РНК 3 'и 5' концы обычно присутствующие в молекуле РНК, были соединены вместе. Эта особенность наделяет круговую РНК многочисленными свойствами, многие из которых были идентифицированы только недавно.
Многие типы кольцевой РНК возникают из генов, кодирующих иначе белки. Было показано, что некоторые кольцевые РНК кодируют белки.[1][2] Некоторые типы кольцевой РНК недавно показали потенциал в качестве регуляторов генов. Биологическая функция большинства кольцевых РНК неясна.
Поскольку кольцевая РНК не имеет 5 'или 3' концов, они устойчивы к экзонуклеаза опосредованной деградацией и предположительно более стабильны, чем большинство линейных РНК в клетках.[3] Циркулярная РНК связана с некоторыми заболеваниями, такими как рак.[4]
Сплайсинг РНК
В отличие от генов в бактерии, эукариотический гены разделены некодирующими последовательностями, известными как интроны. У эукариот, когда ген транскрибируется из ДНК в информационная РНК (мРНК), промежуточные интроны удаляются, остается только экзоны в зрелой мРНК, которую впоследствии можно транслировать с образованием белкового продукта.[5] В сплайсосома,[5] Комплекс белок-РНК, расположенный в ядре, катализирует сплайсинг следующим образом:
- В сплайсосома признает интрон, который фланкирован специфическими последовательностями на его 5 'и 3' концах, известных как донорный сайт сплайсинга (или 5 'сайт сплайсинга) и акцепторный сайт сплайсинга (или 3' сайт сплайсинга), соответственно.
- Затем 5'-последовательность сайта сплайсинга подвергается нуклеофильный атака нисходящей последовательностью, называемой точкой ветвления, в результате образуется круговая структура, называемая лариатом.
- Затем свободный 5 'экзон атакует сайт сплайсинга 3', соединяя два экзона и высвобождая структуру, известную как интронный лариат. В дальнейшем интронный лариат разветвляется и быстро разрушается.[5]

Альтернативная сварка
Альтернативная сварка представляет собой феномен, посредством которого один транскрипт РНК может давать различные белковые продукты в зависимости от того, какие сегменты считаются «интронами», а какие - «экзонами» во время каждого события сплайсинга.[5] Хотя это не относится к людям, это частичное объяснение того факта, что человек и другие, гораздо более простые виды (например, нематоды) имеют одинаковое количество генов (в диапазоне от 20 до 25 тысяч).[6] Один из самых ярких примеров альтернативного сращивания - это Дрозофила DSCAM ген. Этот единственный ген может дать начало примерно 30 тысячам различных изоформ с альтернативным сплайсингом.[7]
Неканонический сплайсинг
Скремблирование экзонов
Скремблирование экзонов, также называемое перетасовкой экзонов, описывает событие, при котором экзоны сплайсируются в «неканоническом» (атипичном) порядке. Скремблирование экзонов может происходить тремя способами:
- Дупликация тандемного экзона в геноме, что часто встречается при раке
- Трансплайсинг (в котором сливаются два транскрипта РНК), что приводит к линейному транскрипту, который содержит экзоны, которые, например, могут происходить из генов, кодируемых на двух разных хромосомах. Транс-сплайсинг очень распространен в C. elegans
- Донорный сайт сплайсинга присоединяется к акцепторному сайту сплайсинга дальше по ходу первичного транскрипта, что дает кольцевой транскрипт.[8]
Представление о том, что циркуляризованные транскрипты являются побочными продуктами несовершенного сплайсинга, подтверждается низкой численностью и отсутствием консервативности последовательности большинства циркулярных РНК,[9] но недавно был брошен вызов.[8][10][11]
Характеристики кольцевой РНК
Полногеномная идентификация circRNA
Несколько исследовательских групп охарактеризовали кольцевые РНК, отсортировав обширные коллекции данных секвенирования РНК.[8][9][11][12][13] Поскольку circRNA не являются полиаденилированными, данные poly (A)-selected RNA-seq не могут быть использованы для обнаружения circRNA. Чтобы идентифицировать потенциальные изоформы кольцевой РНК, обычно ищут последовательность чтения показывает соединение между двумя "зашифрованными" экзонами.
Ниже приводится краткое изложение результатов каждой группы:
Salzman et al. 2012 г.[8]
Исследовательская группа Salzman et al. изначально хотел идентифицировать специфические для рака события скремблирования экзонов. В итоге они обнаружили зашифрованные экзоны в большом количестве нормальных и раковых клеток. Они обнаружили, что изоформы скремблированных экзонов составляют около 10% от общего количества изоформ транскриптов в лейкоциты. Они также идентифицировали 2748 скремблированных изоформ в HeLa и H9 эмбриональные стволовые клетки. Они обнаружили, что примерно 1 из 50 экспрессируемых генов продуцирует изоформы скремблированных транскриптов по крайней мере в 10% случаев. Некоторые из тестов на округлость включали: (1) Образцы, обработанные РНКаза R, фермент, который разрушает линейные РНК, но не кольцевые РНК, и (2) Проверено на наличие поли-А хвосты (не должно присутствовать в круговой молекуле). Их вывод заключался в том, что 98% скремблированных изоформ представляли circRNA.
Jeck et al. 2013[11]
- Лечил человека фибробласт РНК с РНКазой R для обогащения кольцевыми РНК
- Использованы три категории «строгости» (низкая, средняя, высокая) для классификации циклических транскриптов на основе их уровней распространенности.
- Включая "низкую" категорию, ~ 1 из 8 экспрессируемых генов продуцировал определяемые уровни circRNA.
- Значительно выше, чем число Зальцмана (вверху)
- Может быть из-за большего глубина секвенирования
Memczak et al. 2013[12]
- Разработал вычислительный метод для обнаружения циркуРНК.
- de novo обнаружены циркуРНК у людей, мышей и C. elegans и всесторонне их проверили
- Обнаружено, что circRNA часто экспрессируются тканью / стадией развития.
- Описано, что circRNA могут действовать как антагонисты miRNA, как показано на примере circRNA CDR1as (см. Ниже).
Guo et al. 2014 г.[9]
- Идентифицированные и количественно оцененные человеческие circRNA из данных ENCODE Ribozero RNA-seq
- Большинство circRNA являются минорными изоформами сплайсинга и экспрессируются только в нескольких типах клеток.
- 7 112 человеческих circRNA имеют кольцевые фракции не менее 10%
- circRNA не более консервативны, чем их линейные контроли
- Профилирование рибосом указывает на то, что circRNA не транслируются
- За исключением CDR1as, очень немногие circRNA могут действовать как губки микроРНК.
- Сделали вывод, что большинство circRNA являются несущественными побочными продуктами несовершенного сплайсинга.
Zhang et al. 2014 г.[14]
- Развитый CIRCexplorer идентифицировать тысячи circRNA у человека с помощью p (A) - w / o RNase R RNA-seq data
- Подавляющее большинство идентифицированных высокоэкспрессируемых кольцевых РНК были процессированы из экзонов, расположенных в середине генов RefSeq, что позволяет предположить, что образование кольцевой РНК обычно связано со сплайсингом РНК.
- Большинство кольцевых РНК содержат несколько экзонов, чаще всего от двух до трех экзонов.
- Экзоны кольцевых РНК только с одним кольцевым экзоном были намного длиннее, чем экзоны кольцевых РНК с несколькими кольцевыми экзонами, что указывает на то, что процессинг может предпочесть определенную длину, чтобы максимизировать циркуляризацию экзона (ов).
- Элементы Alu, которые могут образовывать IRAlus, либо конвергентные, либо расходящиеся, накладываются друг на друга через фланкирующие интроны circRNAs параллельным образом с одинаковыми расстояниями до соседних экзонов.
- Формированию кольцевой РНК способствует IRAlus или другие неповторяющиеся, но комплементарные последовательности.
- На эффективность циркуляризации экзонов влияет конкуренция спаривания РНК.
- Альтернативные пары РНК и их конкуренция приводят к альтернативной циркуляризации.
- Как циркуляризация экзона, так и ее регуляция эволюционно динамичны.
Dube et al. 2019 г.[15]
Dube et al., Из лаборатории Cruchaga (https://neurogenomics.wustl.edu/ ) впервые выполнили общегеномный вызов circRNA в нескольких областях мозга в случаях болезни Альцгеймера и в контрольных группах, демонстрируя роль этого вида РНК в здоровье и болезни.
- Оптимизирован и валидирован конвейер для вызова circRNA из рибо-истощенной РНК-seq. Трубопровод доступен здесь[15]
- 3547 circRNA прошли строгий контроль качества в когорте Knight ADRC, которая включает РНК-seq от 13 контрольных и 83 случаев болезни Альцгеймера
- 3924 circRNA прошли строгий контроль качества в наборе данных MSBB.
- Мета-анализ результатов открытия и репликации выявил в общей сложности 148 circRNA, которые были значительно коррелированы с CDR после коррекции FDR.
- 33 прохождение строгой генной коррекции множественного теста Бонферрони 5 × 10−6, включая circHOMER1 (п = 2.21 × 10−18) и circCDR1-AS (п = 2.83 × 10−8), среди прочего
- Экспрессия circRNA не зависела от линейной формы
- Экспрессия circRNA также корректировалась пропорцией клеток.
- Дополнительные исследования демонстрируют, что circRNA коэкспрессируют с известными причинными генами болезни Альцгеймера, такими как ПРИЛОЖЕНИЕ и PSEN1, что указывает на то, что некоторые circRNA также являются частью причинного пути.
- Экспрессия CirRNA в мозге больше объясняла клинические проявления болезни Альцгеймера, чем количество аллелей APOε4, что позволяет предположить, что это может быть использовано в качестве потенциального биомаркера болезни Альцгеймера.
Длина circRNA
Недавнее исследование circRNA человека показало, что эти молекулы обычно состоят из 1–5 экзонов.[12] Каждый из этих экзонов может быть до 3 раз длиннее, чем средний экспрессируемый экзон,[11] предполагая, что длина экзона может играть роль в принятии решения о том, какие экзоны циркуляризировать. 85% кольцевых экзонов перекрываются с экзонами, которые кодируют белок,[12] хотя сами кольцевые РНК, по-видимому, не транслируются. Во время образования circRNA экзон 2 часто является «акцепторным» экзоном выше по течению.[8]
Интроны, окружающие экзоны, выбранные для циркуляризации, в среднем в 3 раза длиннее, чем те, которые не фланкируют экзоны до окружности,[8][11] хотя пока не ясно, почему это так. По сравнению с областями, не образующими кружков, эти интроны с гораздо большей вероятностью содержат дополнительные инвертированные Алу повторяется; Alu - самый распространенный транспозон в геноме.[11] Было высказано предположение, что за счет того, что Alu повторяет спаривание оснований друг с другом, это может позволить сайтам сплайсинга находить друг друга, тем самым облегчая циркуляризацию.[10][11]
Интроны в circRNA удерживаются с относительно высокой частотой (~ 25%),[9] таким образом добавляя дополнительную последовательность к зрелым circRNA.
Расположение circRNA в клетке
В клетке circRNA преимущественно находятся в цитоплазма, где количество кольцевых РНК-транскриптов, полученных из гена, может быть до десяти раз больше, чем количество связанных линейных РНК, полученных из этого гена. локус. Неясно, как кольцевые РНК выходят из ядро через относительно небольшой ядерная пора. Поскольку ядерная оболочка ломается во время митоз, одна из гипотез состоит в том, что молекулы покидают ядро во время этой фазы клеточный цикл.[11] Однако определенные circRNA, такие как CiRS-7 / CDR1as, экспрессируются в нейрональных тканях,[12][16] где митотическое деление не преобладает.

CircRNA стабильны по сравнению с линейными РНК
У CircRNA отсутствует полиаденилированный хвост, и поэтому предполагается, что они менее подвержены деградации экзонуклеазами. В 2015 году Энука и другие. измерили период полужизни 60 circRNA и их линейных аналогов, экспрессируемых из того же гена-хозяина, и выявили, что средний период полужизни circRNA из клеток молочной железы (18,8-23,7 часа) как минимум в 2,5 раза больше, чем средний период полужизни их линейные аналоги (4,0–7,4 ч).[17] Как правило, время жизни молекул РНК определяет время их отклика.[18] Соответственно, сообщалось, что циркулярные РНК молочных желез медленно реагируют на стимуляцию факторами роста.[17]
Возможные функции кольцевой РНК
Эволюционное сохранение механизмов и сигналов циркуляризации
CircRNA были идентифицированы у разных видов в разных области жизни. В 2011 году Данан и другие. секвенированная РНК из Археи. После переваривания общей РНК с помощью РНКазы R они смогли идентифицировать кольцевые виды, что указывает на то, что circRNA не специфичны для эукариот.[19] Однако эти кольцевые виды архей, вероятно, не образуются посредством сплайсинга, что позволяет предположить, что существуют другие механизмы генерации кольцевой РНК.
Что касается более тесной эволюционной связи, то сравнение РНК из семенников мыши и РНК из клетки человека обнаружило 69 ортологичный circRNA. Например, и люди, и мыши кодируют HIPK2 и HIPK3 гены, два паралогичный киназы, которые продуцируют большое количество circRNA из одного конкретного экзона у обоих видов.[11] Эволюционная консервация усиливает вероятность важной и важной роли циркуляризации РНК.
CDR1as / CiRS-7 как губка miR-7
микроРНК (miRNA) представляют собой небольшие (~ 21nt) некодирующие РНК, которые подавляют трансляцию информационных РНК, участвующих в большом, разнообразном наборе биологических процессов.[20] Они напрямую устанавливают пары оснований для целевой информационные РНК (мРНК) и может запускать расщепление мРНК в зависимости от степени комплементарности.
МикроРНК сгруппированы в «семенные семейства». Члены семьи имеют общие нуклеотиды № 2–7, называемые семенной областью.[21] Аргонавт белки являются «эффекторными белками», которые помогают миРНК выполнять свою работу, в то время как губки микроРНК представляют собой РНК, которые «губят» миРНК определенного семейства, тем самым служа в качестве конкурентные ингибиторы которые подавляют способность miRNA связывать свои мРНК-мишени, благодаря наличию множества сайтов связывания, которые распознают конкретную затравочную область.[21] Определенные кольцевые РНК имеют много сайтов связывания miRNA, что дает ключ к разгадке того, что они могут функционировать в губчатом состоянии. Две недавние статьи подтвердили эту гипотезу, подробно изучив кольцевую губку под названием CDR1as / CiRS-7, в то время как другие группы не нашли прямых доказательств того, что кольцевые РНК действуют как губки miRNA, анализируя потенциальное взаимодействие кольцевых РНК с белком аргонавта (AGO) с использованием данные высокопроизводительного секвенирования РНК, выделенной перекрестным связыванием и иммунопреципитацией (HITS-CLIP).[13]
CDR1as / CiRS-7 кодируется в геноме антисмысловой к человеку CDR1 (ген) локус (отсюда и название CDR1as),[12] и нацелен на miR-7 (отсюда и название CiRS-7 - Губка с круговой РНК для miR-7).[16] Он имеет более 60 сайтов связывания miR-7, что намного больше, чем у любой известной линейной губки miRNA.[12][16]
AGO2 - это белок Argonaute, связанный с miR-7 (см. Выше). Хотя CDR1as / CiRS-7 может расщепляться miR-671 и связанным с ним белком Argonaute,[16] он не может расщепляться miR-7 и AGO2. Активность расщепления микроРНК зависит от комплементарности за пределами 12-го нуклеотидного положения; ни один из сайтов связывания CiRS-7 не отвечает этому требованию.
Эксперимент с данио, которые не имеют локуса CDR1 в своем геноме, свидетельствует об активности губки CiRS-7. Во время развития miR-7 сильно экспрессируется в головном мозге рыбок данио. Чтобы заставить замолчать экспрессию miR-7 у рыбок данио, Мемчак и его коллеги воспользовались инструментом под названием морфолино, которые могут образовывать пары и связывать целевые молекулы.[22] Лечение морфолино оказывало такое же сильное влияние на развитие среднего мозга, как эктопическая экспрессия CiRS-7 в мозге рыбок данио с использованием инъекционных плазмиды. Это указывает на значительное взаимодействие между CiRS-7 и miR-7 in vivo.[12]
Еще одна примечательная круглая губка miRNA - это SRY. SRY, который высоко экспрессируется в семенниках мышей, функционирует как miR-138 губка.[16][23] В геноме SRY окружен длинным перевернутые повторы (IR) длиной более 15,5 килобаз (кб). Когда один или оба IR удаляются, циркуляризации не происходит. Именно это открытие привело к идее инвертированных повторов, обеспечивающих циркуляризацию.[24]
Поскольку губки с кольцевой РНК характеризуются высокими уровнями экспрессии, стабильностью и большим количеством сайтов связывания miRNA, они, вероятно, будут более эффективными губками, чем те, которые являются линейными.[10]
Другие возможные функции circRNA
Хотя в последнее время внимание было сосредоточено на функциях «губки» circRNA, ученые рассматривают также несколько других функциональных возможностей. Например, некоторые участки взрослой мыши гиппокамп демонстрируют экспрессию CiRS-7, но не miR-7, предполагая, что CiRS-7 может выполнять роли, которые не зависят от взаимодействия с miRNA.[12]
Возможные роли включают следующее:
- Привязка к РНК-связывающие белки (RBP) и РНК, помимо миРНК, с образованием комплексов РНК-белок.[10] Эти комплексы могут регулировать взаимодействия RBP и РНК, например, с каноническим линейным транскриптом гена.[8]
- Производство белка
- Чен и Сарнов 1995 показали, что синтетическая циркулярная РНК, содержащая IRES (внутренний сайт входа в рибосомы), продуцирует белковый продукт. in vitro, тогда как без IRES этого не произошло. Хотя протестированная circRNA была чисто искусственной конструкцией, Чен и Сарнов заявили в своей статье, что им было бы интересно узнать, содержат ли круги элементы IRES.[25]
- Jeck и другие. 2013: Испытаны природные circRNA, которые содержат трансляцию «стартовый кодон». Однако ни одна из этих молекул не связывалась с рибосомами, что позволяет предположить, что многие circRNA не могут транслироваться. in vivo.[11]
- Транспортировка miRNA внутри клетки. Тот факт, что CiRS-7 может быть разрезан с помощью miR-671, может указывать на существование системы для высвобождения "нагрузки" miRNAs в подходящее время.[26]
- Регулирование мРНК в клетке посредством ограниченного спаривания оснований. Формально возможно, что miR-7 сдерживает регуляторную активность CiRS-7, а не наоборот![12][26]
Циркулярные интронные длинные некодирующие РНК (циРНК)
Обычно интронные лариаты (см. Выше) разветвляются и быстро деградируют. Однако отказ от разветвления может привести к образованию кольцевых интронных длинных некодирующих РНК, также известных как ciRNAs.[27] Образование CiRNA, скорее, чем случайный процесс, по-видимому, зависит от присутствия специфических элементов рядом с сайтом 5 'сплайсинга и сайтом точки ветвления (см. Выше).
CiRNA отличаются от circRNA тем, что они заметно присутствуют в ядро а не цитоплазма. Кроме того, эти молекулы содержат мало (если вообще есть) сайтов связывания miRNA. Вместо того, чтобы действовать как губки, ciRNAs, по-видимому, регулируют экспрессию своих родительских генов. Например, относительно обильная циРНК, называемая ci-ankrd52, положительно регулирует Pol II транскрипция. Многие циРНК остаются на своих «сайтах синтеза» в ядре. Однако циРНК может выполнять другие функции, помимо простой регуляции своих родительских генов, поскольку циРНК действительно локализуются в дополнительных сайтах ядра, отличных от их «сайтов синтеза».[27]
Циркулярная РНК и болезнь
Как и в большинстве тем в молекулярная биология, важно рассмотреть, как круговую РНК можно использовать в качестве инструмента для помощи человечеству. Учитывая ее (1) изобилие, (2) эволюционную сохранность и (3) потенциальную регулирующую роль, стоит изучить, как кольцевую РНК можно использовать для изучения патогенез и разработать терапевтические вмешательства. Например:
- Круговой ANRIL (cANRIL) - это кольцевая форма ANRIL, длинной некодирующей РНК (нкРНК). Экспрессия канрила коррелирует с риском атеросклероз, заболевание, при котором артерии становятся твердыми. Было высказано предположение, что CANRIL может изменять INK4 / Экспрессия ARF, что, в свою очередь, увеличивает риск атеросклероза.[28] Дальнейшее изучение экспрессии cANRIL потенциально может быть использовано для предотвращения или лечения атеросклероза.
- miR-7 играет важную регуляторную роль в нескольких раки И в болезнь Паркинсона, что является дегенеративным заболеванием нейронов.[16] Возможно, активность губки CiRS-7 может помочь в противодействии активности miR-7. Если круговая активность губки действительно может помочь в противодействии вредной активности miRNA, ученым нужно будет найти лучший способ ввести экспрессию губки, возможно, через трансген, который является синтетическим геном, который передается между организмами. Также важно учитывать, как трансгены могут экспрессироваться только в определенных тканях или экспрессироваться только при индукции.[21]
- Было обнаружено, что циркулярные РНК регулируются гипоксией, особенно было обнаружено, что циркулярная РНК cZNF292 обладает проангиогенной активностью в эндотелиальных клетках.[13]
Циркулярные РНК играют роль в патогенезе болезни Альцгеймера
Dube et al.,[29] впервые продемонстрировали, что кольцевые РНК мозга (circRNA) являются частью патогенных событий, которые приводят к Болезнь Альцгеймера (Дубе и др., Nature Neuroscience 2019). Dube et al. Предположили, что специфическая circRNA будет по-разному экспрессироваться в случаях AD по сравнению с контролем, и что эти эффекты могут быть обнаружены на ранней стадии заболевания. Они оптимизировали и утвердили новый конвейер анализа кольцевых РНК (circRNA). Они выполнили трехэтапный дизайн исследования, используя данные последовательности РНК мозга Knight ADRC в качестве открытия (этап 1), используя данные с горы Синай в качестве репликации (этап 2) и метаанализ (этап 3), чтобы определить наиболее значительная circRNA, дифференциально экспрессируемая при болезни Альцгеймера. Используя его конвейер, они обнаружили 3547 circRNA, которые прошли строгий контроль качества в когорте Knight ADRC, включая RNA-seq от 13 контрольных и 83 случаев болезни Альцгеймера, и 3924 circRNA прошли строгий контроль в наборе данных MSBB. Мета-анализ результатов открытия и репликации выявил в общей сложности 148 circRNA, которые значительно коррелировали с CDR после коррекции FDR. Кроме того, 33 circRNA прошли строгую генную коррекцию множественного теста Бонферрони 5 × 10-6, включая circHOMER1 (P = 2,21 × 10−18) и circCDR1-AS (P = 2,83 × 10−8) и другие. Они также выполнили дополнительный анализ, чтобы продемонстрировать, что экспрессия circRNA не зависела от линейной формы, а также от доли клеток, которая может затруднять анализ последовательности РНК мозга в исследованиях болезни Альцгеймера. Они выполнили анализ коэкспрессии всех circRNA вместе с линейными формами и обнаружили, что circRNA, включая те, которые дифференциально экспрессировались при болезни Альцгеймера по сравнению с контролем, коэкспрессировались с известными причинными генами болезни Альцгеймера, такими как APP и PSEN1, что указывает на то, что некоторые circRNA также являются частью причинного пути. Они также продемонстрировали, что экспрессия цирРНК в мозге больше объясняет клинические проявления болезни Альцгеймера, чем количество аллелей APOε4, что позволяет предположить, что это может быть использовано в качестве потенциального биомаркера болезни Альцгеймера. Это важное исследование для данной области, поскольку это первый раз, когда circRNA количественно оцениваются и проверяются (с помощью ПЦР в реальном времени) в образцах человеческого мозга в масштабе всего генома и в больших и хорошо изученных когортах. Это также демонстрирует, что эти формы РНК могут быть связаны со сложными признаками, включая болезнь Альцгеймера, поможет понять биологические события, которые приводят к заболеванию.
Вироиды как кольцевые РНК
Вироиды - это в основном патогены растений, которые состоят из коротких участков (несколько сотен азотистых оснований) высоко комплементарных, кольцевых, одноцепочечных и некодирующих РНК без белковой оболочки. По сравнению с другими инфекционными патогенами растений, вироиды чрезвычайно малы по размеру - от 246 до 467 азотистых оснований; таким образом, они состоят из менее чем 10 000 атомов. Для сравнения, длина генома самых маленьких известных вирусов, способных вызвать инфекцию, составляет около 2000 нуклеотидных оснований.[30]
Базы данных
Были созданы различные базы данных для идентификации circRNA, корреляции с типами рака и проверки. [1] [2] [3] [4] [5].
Смотрите также
Рекомендации
- ^ «Новое исследование показывает, что кольцевая РНК может кодировать белки». Science Daily. 23 марта 2017 г.. Получено 3 мая 2018.
- ^ Памудурти, Нагарджуна Редди; Барток, Оснат; Йенс, Марвин; и другие. (Апрель 2017 г.). «Трансляция CircRNA». Молекулярная клетка. 66 (1): 9–21.e7. Дои:10.1016 / j.molcel.2017.02.021. ЧВК 5387669. PMID 28344080.

- ^ Джек, WR; Соррентино, штат Джерси; Ван, К; и другие. (Февраль 2013). «Круглые РНК многочисленны, консервативны и связаны с повторами ALU». РНК. 19 (2): 141–57. Дои:10.1261 / rna.035667.112. ЧВК 3543092. PMID 23249747.
- ^ Вромман, Мариеке; Вандесомпеле, Джо; Волдерс, Питер-Ян. «Замыкая круг: современное состояние и перспективы создания кольцевых баз данных РНК». Брифинги по биоинформатике. Дои:10.1093 / bib / bbz175.
- ^ а б c d Рис, JB (2010). Кэмпбелл Биология (9-е изд.). Сан-Франциско: Бенджамин Каммингс.
- ^ Ю, Дж; Hu, S; Ван, Дж; Вонг, Г.К .; Ли, S; Лю, Б; Дэн, Y; Дай, L; Чжоу, Y; Чжан, X; Цао, М; Лю, Дж; Вс, Дж; Тан, Дж; Чен, Y; Хуанг, X; Линь, Вт; Ye, C; Тонг, Вт; Cong, L; Гэн, Дж; Хан, Y; Ли, Л; Ли, Вт; Обнимать; Хуанг, X; Ли, Вт; Ли, Дж; Лю, Дж; и другие. (2002). «Проект последовательности генома риса (Orya sativa L. ssp. Indica)». Наука. 296 (5565): 79–92. Bibcode:2002Наука ... 296 ... 79Y. Дои:10.1126 / science.1068037. PMID 11935017.
- ^ Celotto, A.M .; Грейвли, Б. (2001). «Альтернативный сплайсинг пре-мРНК Drosophila Dscam регулируется как во времени, так и в пространстве». Генетика. 159 (2): 599–608. ЧВК 1461822. PMID 11606537.
- ^ а б c d е ж грамм Зальцман, Дж; Gawad, C .; Wang, P.L .; Lacayo, N; Браун, ПО (2012). «Циркулярные РНК - преобладающая изоформа транскриптов сотен генов человека в различных типах клеток». PLOS ONE. 7 (2): e30733. Bibcode:2012PLoSO ... 730733S. Дои:10.1371 / journal.pone.0030733. ЧВК 3270023. PMID 22319583.
- ^ а б c d Guo, J.U .; Agarwal, V; Guo, H; Бартель, Д.П. (2014). «Расширенная идентификация и характеристика кольцевых РНК млекопитающих». Геномная биология. 15 (7): 409. Дои:10.1186 / s13059-014-0409-z. ЧВК 4165365. PMID 25070500.
- ^ а б c d Wilusz, J.E .; Шарп, Пенсильвания (2013). «Замкнутый путь к некодирующей РНК» (PDF). Наука. 340 (6131): 440–41. Bibcode:2013Наука ... 340..440Вт. Дои:10.1126 / science.1238522. ЧВК 4063205. PMID 23620042.
- ^ а б c d е ж грамм час я j Джек, WR; Соррентино, штат Джерси; Ван, К; Слевин, М.К .; Бурд, CE; Лю, Дж; Марзлуфф, ВФ; Шарплесс, NE (2013). «Круглые РНК многочисленны, консервативны и связаны с повторами ALU». РНК. 19 (2): 141–57. Дои:10.1261 / rna.035667.112. ЧВК 3543092. PMID 23249747.
- ^ а б c d е ж грамм час я j Мемчак, S; Йенс, М; Элефсиниоти, А; Torti, F; Крюгер, Дж; Рыбак, А; Maier, L; Mackowiak, SD; Gregersen, LH; Munschauer, M; Loewer, A; Зибольд, У; Ландталер, М; Кокс, С; le Noble, F; Раевский, N (2013). «Циркулярные РНК - это большой класс животных РНК с регулирующей способностью». Природа. 495 (7441): 333–8. Bibcode:2013Натура.495..333М. Дои:10.1038 / природа11928. PMID 23446348.
- ^ а б c Бекель, Джес-Нильс; Хаэ, Николас; Heumüller, Andreas W .; Чен, Вэй; Boon, Reinier A .; Стеллос, Константинос; Zeiher, Andreas M .; Джон, Дэвид; Учида, Сидзука (2015-10-23). «Идентификация и характеристика регулируемой гипоксией эндотелиальной циркулярной РНК». Циркуляционные исследования. 117 (10): 884–890. Дои:10.1161 / CIRCRESAHA.115.306319. ISSN 1524-4571. PMID 26377962.
- ^ Чжан, Сяо-Оу; Ван, Хай-Бин; Чжан, Ян; Лу, Сюйхуа; Чен, Линь-Лин; Ян, Ли (2014-09-25). «Опосредованная комплементарной последовательностью циркуляризация экзона». Клетка. 159 (1): 134–147. Дои:10.1016 / j.cell.2014.09.001. ISSN 1097-4172. PMID 25242744.
- ^ а б Дубе, Умбра; Del-Aguila, Хорхе Л .; Ли, Зеран; Бадд, Джон П .; Цзян, Шань; Сюй, Саймон; Ибанез, Лаура; Фернандес, Мария Виктория; Фариас, Фабиана; Нортон, Джоанна; Генч, Джен (07.10.2019). «Атлас экспрессии корковой кольцевой РНК в мозге при болезни Альцгеймера демонстрирует клинические и патологические ассоциации». Природа Неврология. 22 (11): 1903–1912. Дои:10.1038 / s41593-019-0501-5. ISSN 1546-1726. ЧВК 6858549. PMID 31591557.
- ^ а б c d е ж Hansen, T.B .; Дженсен, Т.И.; Clausen, BH; Bramsen, JB; Finsen, B; Дамгаард, СК; Kjems, J (2013). «Природные круги РНК функционируют как эффективные губки для микроРНК». Природа. 495 (7441): 384–88. Bibcode:2013Натура.495..384H. Дои:10.1038 / природа11993. PMID 23446346.
- ^ а б Enuka, Y .; Лауриола, М; Feldman, M.E .; Sas-Chen, A .; Улицкий, И .; Ярден, Ю. (2015). «Циркулярные РНК являются долгоживущими и обнаруживают лишь минимальные ранние изменения в ответ на фактор роста». Исследования нуклеиновых кислот. 44 (3): 1370–83. Дои:10.1093 / нар / gkv1367. ЧВК 4756822. PMID 26657629.
- ^ Снеппен, К. (2014). Модели жизни - динамика и регуляция в биологических системах. Сан-Франциско: Издательство Кембриджского университета. ISBN 978-1-107-06190-3.
- ^ Данан, М; Шварц, S; Edelheit, S; Сорек, Р. (2012). «Обширное транскриптомное открытие кольцевых РНК в архее». Исследования нуклеиновых кислот. 40 (7): 3131–42. Дои:10.1093 / nar / gkr1009. ЧВК 3326292. PMID 22140119.
- ^ Дин, XC; Вейлер, Дж; Гроссханс, Х (2009). «Регулирующие регуляторы: механизмы, контролирующие созревание микроРНК». Тенденции в биотехнологии. 27 (1): 27–36. Дои:10.1016 / j.tibtech.2008.09.006. PMID 19012978.
- ^ а б c Эберт, MS; Шарп, Пенсильвания (2010). «Губки MicroRNA: успехи и возможности». РНК. 16 (11): 2043–50. Дои:10.1261 / rna.2414110. ЧВК 2957044. PMID 20855538.
- ^ Саммертон, Дж (1999). «Морфолино-антисмысловые олигомеры: случай независимого от РНКазы Н структурного типа». Biochimica et Biophysica Acta (BBA) - Структура и экспрессия гена. 1489 (1): 141–58. Дои:10.1016 / S0167-4781 (99) 00150-5. PMID 10807004.
- ^ Capel, B; Свейн, А; Николис, S; Хакер, А; Уолтер, М; Купман, П; Гудфеллоу, П; Ловелл-Бэдж, Р. (1993). «Циркулярные транскрипты гена Sry, определяющего семенники, в семенниках взрослых мышей». Клетка. 73 (5): 1019–30. Дои:10.1016 / 0092-8674 (93) 90279-у. PMID 7684656.
- ^ Дубин Р.А.; Казми, Массачусетс; Острер, H (1995). «Инвертированные повторы необходимы для циркуляризации транскрипта Sry семенников мыши». Ген. 167 (1–2): 245–48. Дои:10.1016/0378-1119(95)00639-7. PMID 8566785.
- ^ Chen, CY; Сарнов, П. (1995). «Инициирование синтеза белка эукариотическим трансляционным аппаратом на кольцевых РНК». Наука. 268 (5209): 415–17. Bibcode:1995 Наука ... 268..415C. Дои:10.1126 / science.7536344. PMID 7536344.
- ^ а б Hentze, MW; Прейсс, Т. (2013). «Циркулярные РНК: вариации загадки сплайсинга». Журнал EMBO. 32 (7): 923–25. Дои:10.1038 / emboj.2013.53. ЧВК 3616293. PMID 23463100.
- ^ а б Zhang, Y; Чжан, старший офицер; Чен, Т; Xiang, JF; Инь, QF; Xing, YH; Чжу, S; Ян, Л; Чен, LL (2013). «Циркулярные интронные длинные некодирующие РНК». Молекулярная клетка. 51 (6): 1–15. Дои:10.1016 / j.molcel.2013.08.017. PMID 24035497.
- ^ Бурд, CE; Джек, WR; Лю, Y; Sanoff, HK; Ван, З; Шарплесс, NE (2010). "Экспрессия линейных и новых круговых форм INK4 / ARF-ассоциированной некодирующей РНК коррелирует с риском атеросклероза". PLOS Genetics. 6 (12): e1001223. Дои:10.1371 / journal.pgen.1001233. ЧВК 2996334. PMID 21151960.
- ^ Дубэ, У; Del-Aguila, JL; Ли, Z; Budde, JP; Цзян, S; Hsu, S; Ibanez, L; Фернандес, М.В.; Farias, F; Нортон, Дж; Gentsch, J; Ванга, Ф; Сеть доминантно наследуемой болезни Альцгеймера, (ДИАН) .; Саллоуэй, S; Мастера, CL; Ли, JH; Graff-Radford, NR; Chhatwal, JP; Bateman, RJ; Моррис, JC; Карч, СМ; Харари, О; Кручага, К. (ноябрь 2019 г.). «Атлас экспрессии корковой кольцевой РНК в мозге при болезни Альцгеймера демонстрирует клинические и патологические ассоциации». Природа Неврология. 22 (11): 1903–1912. Дои:10.1038 / s41593-019-0501-5. ЧВК 6858549. PMID 31591557.
- ^ Sanger, H.L .; Klotz, G .; Riesner, D .; Gross, H.J .; Кляйншмидт, А. К. (1 ноября 1976 г.). «Вироиды представляют собой одноцепочечные ковалентно замкнутые кольцевые молекулы РНК, существующие в виде стержневидных структур с большим количеством пар оснований». Труды Национальной академии наук. 73 (11): 3852–3856. Bibcode:1976PNAS ... 73.3852S. Дои:10.1073 / pnas.73.11.3852. ЧВК 431239. PMID 1069269.
внешняя ссылка
- База данных CIRCpedia[1]
- база данных circRNABase [2]
- база данных circBase.org - недавно опубликованная обширная база данных кольцевых РНК[3]
- База данных Circ2Traits
- ^ "ЦИРЦпедия". www.picb.ac.cn. Получено 2016-08-09.
- ^ Ли, JH; Лю, S; Чжоу, H; Qu, LH; Ян, Дж. Х. (январь 2014 г.). «starBase v2.0: расшифровка сетей взаимодействия miRNA-ceRNA, miRNA-ncRNA и белок-РНК из крупномасштабных данных CLIP-Seq». Исследования нуклеиновых кислот. 42 (Выпуск базы данных): D92–7. Дои:10.1093 / nar / gkt1248. ЧВК 3964941. PMID 24297251.
- ^ Глазар, Петар (2014). "circBase: база данных кольцевых РНК". РНК. 20 (11): 1666–70. Дои:10.1261 / rna.043687.113. ЧВК 4201819. PMID 25234927.