Обратный повтор - Inverted repeat
Было высказано предположение, что палиндромная последовательность быть слился в эту статью. (Обсудить) Предлагается с октября 2020 года. |
An перевернутый повтор (или ИК) представляет собой одноцепочечную последовательность нуклеотиды далее по течению обратное дополнение.[1] Промежуточная последовательность нуклеотидов между начальной последовательностью и обратным комплементом может иметь любую длину, включая ноль. Например, 5 '--- TTACGnnnnnnCGTAA --- 3 ' представляет собой инвертированную повторяющуюся последовательность. Когда промежуточная длина равна нулю, составная последовательность представляет собой палиндромная последовательность.
Оба перевернутых повтора и прямые повторы составляют типы нуклеотидные последовательности которые происходят повторно. Эти повторяющиеся последовательности ДНК часто варьируются от пары нуклеотидов до целого ген, в то время как близость повторяющихся последовательностей варьирует от широко рассредоточенных до простых тандемные массивы.[2] Короткая тандемный повтор последовательности могут существовать от нескольких копий в небольшой области до тысяч копий, разбросанных по геному большинства эукариоты.[3] Повторите последовательности примерно с 10–100 пар оснований известны как мини-спутники, в то время как более короткие повторяющиеся последовательности, содержащие в основном 2–4 пары оснований, известны как микроспутники.[4] Наиболее распространенные повторы включают динуклеотидные повторы, которые имеют базы AC на одной цепи ДНК и GT на дополнительной цепи.[2] Некоторые элементы геном с уникальными последовательностями функционируют как экзоны, интроны и регуляторная ДНК.[5] Хотя наиболее знакомыми локусами повторяющихся последовательностей являются центромера, а теломер,[5] большая часть повторяющихся последовательностей в геноме находится среди некодирующая ДНК.[4]
Инвертированные повторы выполняют ряд важных биологических функций. Они определяют границы в транспозоны и указать регионы, способные к самодополнению. базовая пара (области в одной последовательности, которые могут образовывать пары оснований друг с другом). Эти свойства играют важную роль в нестабильности генома.[6] и способствовать не только клеточная эволюция и генетическое разнообразие[7] но также мутация и болезнь.[8] Для детального изучения этих эффектов был разработан ряд программ и баз данных, помогающих в обнаружении и аннотации инвертированных повторов в различных геномах.
Что такое перевернутые повторы
Пример перевернутого повтора
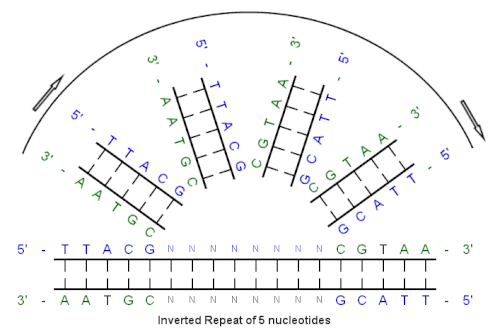
Начиная с этой начальной последовательности:
5'-TTACG-3 '
Дополнение, созданное спариванием оснований:
3'-AATGC-5 '
Обратное дополнение:
5'-CGTAA-3 '
И инвертированная повторяющаяся последовательность:
5 '--- TTACGnnnnnnCGTAA --- 3 '
«nnnnnn» представляет собой любое количество промежуточных нуклеотидов.
Vs. прямой повтор
Прямое повторение происходит, когда последовательность повторяется с тем же шаблоном ниже по течению.[1] Нет ни инверсии, ни обратного дополнения, связанных с прямым повторением. Нуклеотидная последовательность, выделенная жирным шрифтом, означает повторяющуюся последовательность. Он может иметь или не иметь промежуточных нуклеотидов.
- 5´ TTACGnnnnnnTTACG 3´
- 3´ AATGCnnnnnnAATGC 5´
С лингвистической точки зрения типичное прямое повторение сравнимо с рифмой, например "твремя на двремя".
Vs. тандемный повтор
Прямое повторение с нет промежуточные нуклеотиды между исходной последовательностью и ее нисходящей копией представляют собой Тандемный повтор. Нуклеотидная последовательность, выделенная жирным шрифтом, означает повторяющуюся последовательность.
- 5´ TTACGTTACG 3´
- 3´ AATGCAATGC 5´
С лингвистической точки зрения типичный тандемный повтор можно сравнить с заиканием или намеренно повторяемыми словами, например, «пока».
Vs. палиндром
Инвертированная повторяющаяся последовательность с нет промежуточные нуклеотиды между исходной последовательностью и ее нижележащей последовательностью обратное дополнение это палиндром.[1]
ПРИМЕР:
Шаг 1: начните с перевернутого повтора: 5 'TTACGnnnnnnCGTAA 3'
Шаг 2: удалите промежуточные нуклеотиды: 5 'TTACGCGTAA 3'
Эта результирующая последовательность является палиндромной, потому что она является обратным дополнением самой себя.[1]
- 5 'TTACGCGTAA 3' тестовая последовательность (из шага 2 с удаленными промежуточными нуклеотидами)
- 3 'AATGCGCATT 5' дополнение тестовой последовательности
- 5 'TTACGCGTAA 3' обратное дополнение Это то же самое, что и тестовая последовательность выше, и, следовательно, это палиндром.
Биологические особенности и функциональность
Условия, благоприятствующие синтезу
Разнообразные повторы всего генома происходят от сменные элементы, которые теперь понимают, что они «прыгают» по разным участкам генома, не передавая свои оригинальные копии.[9] Последующее перемещение одних и тех же последовательностей в течение многих поколений обеспечивает их множественность по всему геному.[9] Ограниченный рекомбинация последовательностей между двумя отдельными элементами последовательности, известными как консервативная сайт-специфическая рекомбинация (CSSR) приводит к инверсиям сегмента ДНК на основе расположения последовательностей распознавания рекомбинации на донорской ДНК и реципиентной ДНК.[9] Опять же, ориентация двух сайтов рекомбинации в молекуле донорной ДНК относительно асимметрии промежуточных последовательностей расщепления ДНК, известной как область кроссовера, имеет решающее значение для образования либо инвертированных повторов, либо прямых повторов.[9] Таким образом, рекомбинация, происходящая в паре перевернутых сайтов, инвертирует последовательность ДНК между двумя сайтами.[9] Наблюдались очень стабильные хромосомы со сравнительно меньшим количеством инвертированных повторов, чем прямых повторов, что предполагает связь между стабильностью хромосом и количеством повторов.[10]
Регионы, где присутствие обязательно
Конечные инвертированные повторы наблюдались в ДНК различных эукариотических транспозонов, хотя их источник остается неизвестным.[11] Инвертированные повторы в основном обнаруживаются в истоках репликации клеточного организма и органелл, которые варьируются от фаговых плазмид, митохондрий и эукариотических вирусов до клеток млекопитающих.[12] Источники репликации фага G4 и других родственных фагов включают сегмент из почти 139 нуклеотидных оснований, который включает три инвертированных повтора, которые необходимы для примирования репликации.[12]
В геноме
В значительной степени участки нуклеотидных повторов довольно часто встречаются в составе редких комбинаций ДНК.[13] Три основных повтора, которые в основном встречаются в конкретных конструкциях ДНК, включают очень точные инвертированные повторы гомопурин-гомопиримидин, которые иначе называются H-палиндромами, обычное явление в тройных спиральных H-конформациях, которые могут включать нуклеотидные триады TAT или CGC. Другие могут быть описаны как длинные инвертированные повторы, имеющие тенденцию к образованию шпилек и крестообразных, и, наконец, прямые тандемные повторы, которые обычно существуют в структурах, описываемых как скользящая петля, крестообразная и левосторонняя Z-ДНК.[13]
Распространен у разных организмов
Предыдущие исследования показывают, что повторы являются общей чертой эукариоты в отличие от прокариоты и археи.[13] Другие сообщения предполагают, что, несмотря на сравнительную нехватку повторяющихся элементов в геномах прокариот, они, тем не менее, содержат сотни или даже тысячи больших повторов.[14] Текущий геномный анализ, кажется, предполагает существование большого избытка совершенных инвертированных повторов во многих прокариотических геномах по сравнению с эукариотическими геномами.[15]

Для количественной оценки и сравнения инвертированных повторов у нескольких видов, а именно у архей, см. [16]
Инвертированные повторы в псевдоузлах
Псевдоузлы являются общими структурными мотивами, обнаруженными в РНК. Они образованы двумя вложенными стебель-петли так, чтобы стержень одной конструкции образовывался из петли другой. Есть несколько складных топологии среди псевдоузлов и большой разброс по длине петель, что делает их структурно разнообразной группой.[17]
Инвертированные повторы являются ключевым компонентом псевдоузлов, что можно увидеть на иллюстрации естественного псевдоузла, обнаруженного у человека. компонент теломеразной РНК.[18] В этой структуре задействованы четыре различных набора инвертированных повторов. Наборы 1 и 2 являются стержнем для стержня-петли A и являются частью петли для стержня-петли B. Точно так же наборы 3 и 4 являются стержнем для стержня-петли B и являются частью петли для стержня-петли A.
Псевдоузлы играют в биологии несколько различных ролей. Псевдоузел теломеразы на иллюстрации имеет решающее значение для активности этого фермента.[18] В рибозим для вирус гепатита дельта (HDV) сворачивается в структуру с двойным псевдоузлом и саморасщепляется в своем кольцевом геноме с образованием РНК с одним геномом. Псевдоузлы также играют роль в запрограммированных рибосомальный сдвиг рамки обнаружен в некоторых вирусах и требуется при репликации ретровирусы.[17]
В рибопереключателях
Инвертированные повторы играют важную роль в рибопереключатели, которые представляют собой регуляторные элементы РНК, контролирующие экспрессию генов, продуцирующих мРНК, частью которой они являются.[9] Упрощенный пример флавинмононуклеотид Рибопереключатель (FMN) показан на рисунке. Этот рибопереключатель существует в мРНК стенограмма и имеет несколько стебель-петля структур выше по течению от кодирующая область. Однако на иллюстрации показаны только ключевые стержни-петли, которые были значительно упрощены, чтобы показать роль перевернутых повторов. В этом рибопереключателе есть несколько перевернутых повторов, обозначенных зеленым (желтый фон) и синим (оранжевый фон).
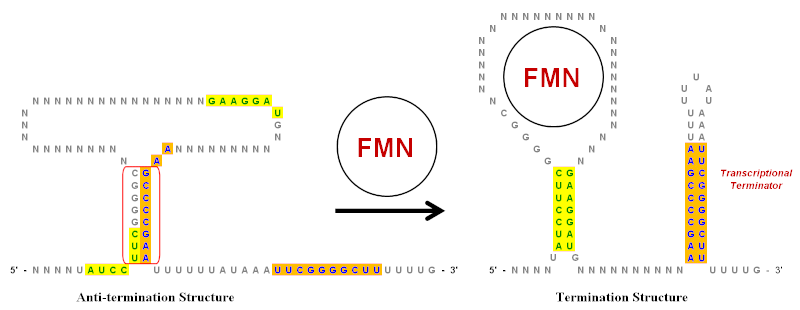
В отсутствие FMN предпочтительной является структура, препятствующая обрыву. конформация для транскрипта мРНК. Он создается спариванием оснований области инвертированного повтора, обведенной красным. Когда FMN присутствует, он может связываться с петлей и предотвращать образование структуры Anti-termination. Это позволяет использовать два разных набора инвертированных повторов для пары оснований и формирования структуры завершения.[19] Стебель-петля на конце 3 'представляет собой терминатор транскрипции потому что последовательность, следующая сразу за ней, представляет собой строку урацилов (U). Если эта стебель-петля образуется (из-за присутствия FMN), когда растущая цепь РНК выходит из РНК-полимераза комплекса, он создаст достаточное структурное напряжение, чтобы вызвать диссоциацию цепи РНК и, таким образом, остановить транскрипцию. Диссоциация происходит легко, потому что пары оснований между U в РНК и A в матричной цепи являются самыми слабыми из всех пар оснований.[9] Таким образом, при более высоких уровнях концентрации FMN подавляет собственную транскрипцию, увеличивая образование терминирующей структуры.
Мутации и болезни
Инвертированные повторы часто описываются как «горячие точки» эукариотической и прокариотической геномной нестабильности.[6] Считается, что длинные инвертированные повторы сильно влияют на стабильность генома различных организмов.[20] Это проиллюстрировано в Кишечная палочка, где геномные последовательности с длинными инвертированными повторами редко реплицируются, а скорее удаляются с большой скоростью.[20] Опять же, длинные инвертированные повторы, наблюдаемые у дрожжей, в значительной степени способствуют рекомбинации в пределах одной и соседних хромосом, что приводит к столь же очень высокой скорости делеции.[20] Наконец, очень высокая скорость делеции и рекомбинации также наблюдалась в областях хромосом млекопитающих с инвертированными повторами.[20] Сообщенные различия в стабильности геномов взаимосвязанных организмов всегда указывают на несоответствие инвертированных повторов.[10] Нестабильность возникает из-за тенденции инвертированных повторов складываться в шпильки или крестообразные структуры ДНК. Эти особые структуры могут препятствовать или мешать репликации ДНК и другим геномным действиям.[6] Таким образом, инвертированные повторы приводят к особым конфигурациям в обоих РНК и ДНК что в конечном итоге может вызвать мутации и болезнь.[8]

На рисунке показан перевернутый повтор, подвергающийся крестообразной экструзии. ДНК в области инвертированного повтора раскручивается, а затем рекомбинирует, образуя четырехстороннее соединение с двумя стебель-петля конструкции. Крестообразная структура возникает из-за того, что инвертированные повторяющиеся последовательности самостоятельно спариваются друг с другом на своей собственной цепи.[21]
Экструдированные крестовины могут привести к мутации сдвига рамки считывания когда последовательность ДНК имеет инвертированные повторы в виде палиндром в сочетании с регионами прямые повторы с обеих сторон. В течение транскрипция, проскальзывание и частичная диссоциация полимеразы из цепи-шаблона может привести как к удаление и вставка мутации.[8] Удаление происходит при раскручивании части шаблон прядь образует стебель-петлю, которую «пропускает» аппарат транскрипции. Вставка происходит, когда стебель-петля формируется в диссоциированной части возникающей (вновь синтезированной) цепи, вызывая двойную транскрипцию части матричной цепи.[8]
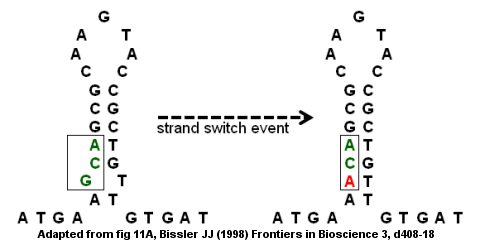
Дефицит антитромбина из-за точечной мутации
Несовершенные перевернутые повторы могут привести к мутации посредством внутрипрядной и межнитевой коммутации.[8] В антитромбин Кодирующая область гена III является примером несовершенного инвертированного повтора, как показано на рисунке справа. стебель-петля структура формируется с выпуклостью внизу, потому что G и T не образуют пары. Событие переключения нити может привести к замене G (в выступе) буквой A, которая устраняет «несовершенство» в перевернутом повторе и обеспечивает более прочную структуру стержень-петля. Однако замена также создает точечная мутация преобразование кодона GCA в ACA. Если за событием переключения нити следует второй раунд Репликация ДНК, мутация может закрепиться в геном и привести к болезни. В частности, миссенс-мутация приведет к дефектному гену и дефициту антитромбина, что может привести к развитию Венозная тромбоэмболия (сгустки крови в вене).[8]

Несовершенный остеогенез в результате мутации сдвига рамки считывания
Мутации в коллаген ген может привести к болезни Несовершенный остеогенез, который характеризуется ломкостью костей.[8] На иллюстрации стержневая петля, образованная из несовершенного инвертированного повтора, мутирована вставкой нуклеотида тимина (Т) в результате переключения между нитями или между нитями. Добавление буквы T создает спаривание оснований «сопоставьте» с аденином (A), который ранее был «бугорком» на левой стороне стержня. Хотя это добавление делает стебель более сильным и улучшает перевернутый повтор, оно также создает мутация сдвига рамки в нуклеотидной последовательности, которая изменяет рамка чтения и приведет к неправильной экспрессии гена.[8]
Программы и базы данных
В следующем списке представлена информация и внешние ссылки на различные программы и базы данных для инвертированных повторов:
- не БД База данных для интегрированных аннотаций и анализа не-B-образующих ДНК мотивов.[22] Эта база данных предоставлена Центром передовых биомедицинских вычислений (ABCC) в тогдашней Национальной лаборатории исследования рака имени Фредерика (FNLCR). Он охватывает А-ДНК и Z-ДНК конформации, иначе известные как "ДНК не-B", потому что они не более обычная B-ДНК форма правши Watson-Crick двойная спираль. Эти «не-B ДНК» включают левостороннюю Z-ДНК, крестообразный, триплекс, тетраплекс и шпилька конструкции.[22] Поиск может выполняться по множеству «повторяющихся типов» (включая инвертированные повторы) и по нескольким видам.
- База данных инвертированных повторов Бостонский университет. Эта база данных представляет собой веб-приложение, которое позволяет запрашивать и анализировать повторы, хранящиеся в проекте PUBLIC DATABASE. Ученые также могут анализировать свои собственные последовательности с помощью алгоритма поиска инвертированных повторов.[23]
- P-MITE: база данных растений MITE - эта база данных для Миниатюрные транспонированные элементы с перевернутым повторением (MITEs) содержит последовательности из геномы растений. Последовательности можно искать или загружать из базы данных.[24]
- EMBOSS это «Открытый программный пакет европейской молекулярной биологии», который работает на UNIX и UNIX-подобный операционные системы.[25] Документация и исходные файлы программы доступны на Сайт EMBOSS. Приложения, специально относящиеся к инвертированным повторам, перечислены ниже:
- EMBOSS einverted: Находит перевернутые повторы в нуклеотидные последовательности. Можно установить пороговые значения, чтобы ограничить область поиска.[25]
- EMBOSS палиндром: Находит палиндромы такие как стержень петля области в нуклеотидных последовательностях. Программа найдет последовательности, которые включают участки несоответствий и промежутков, которые могут соответствовать выпуклости в петле стержня.[25]
использованная литература
- ^ а б c d Ussery, Дэвид В .; Вассенаар, Труди; Борини, Стефано (22 декабря 2008 г.). «Частоты слов, повторы и структуры, связанные с повторами в бактериальных геномах». Вычисления для сравнительной микробной геномики: биоинформатика для микробиологов. Вычислительная биология. 8 (1-е изд.). Springer. С. 133–144. ISBN 978-1-84800-254-8.
- ^ а б Ричардс, Г. Р.; Ричардс, Род-Айленд (25 апреля 1995 г.). «Простые тандемные повторы ДНК и генетическое заболевание человека». Труды Национальной академии наук Соединенных Штатов Америки. 92 (9): 3636–41. Bibcode:1995PNAS ... 92.3636S. Дои:10.1073 / пнас.92.9.3636. ЧВК 42017. PMID 7731957.
- ^ ван Белкум, А; Шерер, S; ван Альфен, L; Verbrugh, H (июнь 1998 г.). «Короткие последовательности ДНК-повторов в прокариотических геномах». Обзоры микробиологии и молекулярной биологии. 62 (2): 275–93. Дои:10.1128 / MMBR.62.2.275-293.1998. ЧВК 98915. PMID 9618442.
- ^ а б Рамель, К. (июнь 1997 г.). «Мини- и микроспутники». Перспективы гигиены окружающей среды. 105 Дополнение 4 (Дополнение 4): 781–9. Дои:10.2307/3433284. JSTOR 3433284. ЧВК 1470042. PMID 9255562.
- ^ а б Эйхлер, EE (август 1998 г.). «Маскарад повторяется: паралогичные ловушки человеческого генома». Геномные исследования. 8 (8): 758–62. Дои:10.1101 / гр. 8.8.758. PMID 9724321.
- ^ а б c Миркин, я; Нараянан, V; Лобачев К.С.; Миркин С.М. (22 июля 2008 г.). «Замедление репликации при нестабильных инвертированных повторах: взаимодействие между шпильками ДНК и белками, стабилизирующими вилку». Труды Национальной академии наук Соединенных Штатов Америки. 105 (29): 9936–41. Bibcode:2008PNAS..105.9936V. Дои:10.1073 / pnas.0804510105. ЧВК 2481305. PMID 18632578.
- ^ Lin, CT; Линь, WH; Лю, Ю.Л .; Whang-Peng, J (1 сентября 2001 г.). «Инвертированные повторы как генетические элементы, способствующие инвертированной дупликации ДНК: последствия для амплификации генов». Исследования нуклеиновых кислот. 29 (17): 3529–38. Дои:10.1093 / nar / 29.17.3529. ЧВК 55881. PMID 11522822.
- ^ а б c d е ж г час Бисслер, Дж. Дж. (27 марта 1998 г.). «Инвертированные повторы ДНК и болезнь человека» (PDF). Границы биологических наук. 3 (4): d408–18. Дои:10.2741 / a284. PMID 9516381. S2CID 12982.
- ^ а б c d е ж г Школа, Джеймс Д. Уотсон, Лаборатория Колд-Спринг-Харбор, Таня А. Бейкер, Массачусетский технологический институт, Стивен П. Белл, Массачусетский технологический институт, Александр Ганн, Лаборатория Колд-Спринг-Харбор, Майкл Левин, Калифорнийский университет, Беркли, Ричард Лосик, Гарвардский университет; со Стивеном К. Харрисоном, Harvard Medical (2014). Молекулярная биология гена (Седьмое изд.). Бостон: Издательство Бенджамин-Каммингс. ISBN 9780321762436.
- ^ а б Ахаз, G; Coissac, E; Неттер, П; Роча, EP (август 2003 г.). «Связь между инвертированными повторами и структурной эволюцией бактериальных геномов». Генетика. 164 (4): 1279–89. ЧВК 1462642. PMID 12930739.
- ^ Zhang, HH; Сюй, ОН; Шен, YH; Хан, MJ; Чжан, З. (январь 2013 г.). «Происхождение и эволюция шести миниатюрных перевернутых транспонированных элементов в Bombyx mori и Rhodnius prolixus». Геномная биология и эволюция. 5 (11): 2020–31. Дои:10.1093 / gbe / evt153. ЧВК 3845634. PMID 24115603.
- ^ а б Пирсон, CE; Зорбас, Н; Цена, ГБ; Заннис-Хаджопулос, М. (октябрь 1996 г.). «Инвертированные повторы, стволовые петли и крестообразные формы: значение для инициации репликации ДНК». Журнал клеточной биохимии. 63 (1): 1–22. Дои:10.1002 / (SICI) 1097-4644 (199610) 63: 1 <1 :: AID-JCB1> 3.0.CO; 2-3. PMID 8891900. S2CID 22204780.
- ^ а б c Херинга, Дж (июнь 1998 г.). «Обнаружение внутренних повторов: насколько они распространены?». Текущее мнение в структурной биологии. 8 (3): 338–45. Дои:10.1016 / S0959-440X (98) 80068-7. PMID 9666330.
- ^ Treangen, TJ; Abraham, AL; Touchon, M; Роча, EP (май 2009 г.). «Генезис, эффекты и судьба повторов в геномах прокариот» (PDF). Обзор микробиологии FEMS. 33 (3): 539–71. Дои:10.1111 / j.1574-6976.2009.00169.x. PMID 19396957.
- ^ Ladoukakis, ED; Эйр-Уокер, А (сентябрь 2008 г.). «Избыток мелких инвертированных повторов у прокариот» (PDF). Журнал молекулярной эволюции. 67 (3): 291–300. Bibcode:2008JMolE..67..291L. CiteSeerX 10.1.1.578.7466. Дои:10.1007 / s00239-008-9151-z. PMID 18696026.
- ^ Хоссейни, М; Пратас, Д; Пинхо, AJ (2017). О роли инвертированных повторов в сходстве последовательностей ДНК. 11-я Международная конференция по практическому применению вычислительной биологии и биоинформатики. PACBB 2017. Достижения в области интеллектуальных систем и вычислений, Том 616. Спрингер, Чам. Достижения в интеллектуальных системах и вычислениях. 616. С. 228–236. Дои:10.1007/978-3-319-60816-7_28. ISBN 978-3-319-60815-0.
- ^ а б Скоба, DW; Мясник, ЮВ (июнь 2005 г.). «Псевдоузлы: структуры РНК с разнообразными функциями». PLOS Биология. 3 (6): e213. Дои:10.1371 / journal.pbio.0030213. ЧВК 1149493. PMID 15941360.

- ^ а б Chen, JL; Грейдер, CW (7 июня 2005 г.). «Функциональный анализ структуры псевдоузла теломеразной РНК человека». Труды Национальной академии наук Соединенных Штатов Америки. 102 (23): 8080–5, обсуждение 8077–9. Bibcode:2005PNAS..102.8080C. Дои:10.1073 / pnas.0502259102. ЧВК 1149427. PMID 15849264.
- ^ Винклер, WC; Коэн-Чаламиш, S; Breaker, RR (10 декабря 2002 г.). «Структура мРНК, которая контролирует экспрессию гена путем связывания FMN». Труды Национальной академии наук Соединенных Штатов Америки. 99 (25): 15908–13. Bibcode:2002PNAS ... 9915908W. Дои:10.1073 / pnas.212628899. ЧВК 138538. PMID 12456892.
- ^ а б c d Стормо, G; Чанг, Кентукки; Варлей, К; Стормо, GD (28 февраля 2007 г.). Холл, Нил (ред.). «Доказательства активного поддержания структур с перевернутыми повторами, идентифицированных с помощью сравнительного геномного подхода». PLOS ONE. 2 (2): e262. Bibcode:2007PLoSO ... 2..262Z. Дои:10.1371 / journal.pone.0000262. ЧВК 1803023. PMID 17327921.
- ^ Рамредди, Т; Сачиданандам, Р. Стрик, TR (май 2011 г.). «Обнаружение крестообразной экструзии в реальном времени с помощью наноманипуляций одномолекулярной ДНК». Исследования нуклеиновых кислот. 39 (10): 4275–83. Дои:10.1093 / nar / gkr008. ЧВК 3105387. PMID 21266478.
- ^ а б Cer, RZ; Донохью, Делавэр; Мудунури, США; Темиз, Н.А. Убыток, MA; Старнер, штат Нью-Джерси; Halusa, GN; Вольфовский, Н; Yi, M; Люк, БТ; Bacolla, A; Коллинз, младший; Стивенс, РМ (январь 2013 г.). "Non-B DB v2.0: база данных предсказанных не-B ДНК-образующих мотивов и связанных с ней инструментов". Исследования нуклеиновых кислот. 41 (Проблема с базой данных): D94 – D100. Дои:10.1093 / нар / gks955. ЧВК 3531222. PMID 23125372.
- ^ Гельфанд, Y; Родригес, А; Бенсон, Г. (январь 2007 г.). «TRDB - База данных тандемных повторов». Исследования нуклеиновых кислот. 35 (Выпуск базы данных): D80–7. Дои:10.1093 / нар / gkl1013. ЧВК 1781109. PMID 17175540.
- ^ Чен, Дж; Ху, Q; Zhang, Y; Лу, С; Куанг, Х (29 октября 2013 г.). «P-MITE: база данных миниатюрных транспонированных элементов с перевернутыми повторами растений». Исследования нуклеиновых кислот. 42 (1): D1176–81. Дои:10.1093 / nar / gkt1000. ЧВК 3964958. PMID 24174541.
- ^ а б c Рис, P; Лонгден, я; Близби, А. (июнь 2000 г.). "EMBOSS: Открытый программный пакет европейской молекулярной биологии". Тенденции в генетике. 16 (6): 276–7. Дои:10.1016 / S0168-9525 (00) 02024-2. PMID 10827456.
внешние ссылки
- Инвертированный + повтор + последовательность в Национальной медицинской библиотеке США Рубрики медицинской тематики (MeSH)