Неконкурентное торможение - Non-competitive inhibition - Wikipedia
Неконкурентное торможение это тип ингибирование ферментов где ингибитор снижает активность фермента и одинаково хорошо связывается с ферментом независимо от того, связал он уже субстрат или нет.[1]
Ингибитор может связываться с ферментом независимо от того, был ли субстрат уже связан, но если он имеет более высокое сродство к связыванию фермента в том или ином состоянии, это называется смешанный ингибитор.[1]
История
За годы работы врачом Михаэлис и его друг (Питер Рона) построили небольшую лабораторию в больнице, и в течение пяти лет Михаэлис успешно публиковался более 100 раз. Во время своего исследования в больнице он первым увидел различные типы торможения; специально используя фруктозу и глюкозу в качестве ингибиторов мальтаза Мероприятия. Мальтаза расщепляет мальтозу на две части: глюкоза или же фруктоза. Результаты этого эксперимента учитывают расхождение неконкурентных и конкурентное торможение. Неконкурентное торможение влияет на kКот значение (но не Kм) на любом заданном графе; этот ингибитор связывается с сайтом, специфичным для определенной молекулы. Михаэлис определил, что при связывании ингибитора фермент будет инактивирован.[2]
Как и многие другие ученые своего времени, Леонор Михаэлис и Мод Ментен работал над реакцией, которая использовалась для изменения конформации сахароза и сделай это лизировать на два продукта - фруктозу и глюкозу.[2] Фермент, участвующий в этой реакции, называется инвертаза, и именно этот фермент, кинетика которого, как утверждали Михаэлис и Ментен, стал революционным для кинетики других ферментов. Выражая скорость изучаемой реакции, они вывели уравнение, описывающее скорость таким образом, чтобы предположить, что она в основном зависит от концентрации фермента, а также от присутствия субстрата, но только в определенной степени.[2][3]
Адриан Браун и Виктор Анри заложили основу для открытий в кинетике ферментов, которыми известны Михаэлис и Ментен.[4] Браун теоретически представил механизм, принятый в настоящее время для кинетики ферментов, но не имел количественных данных, чтобы утверждать это.[4] Виктор Анри внес значительный вклад в кинетику ферментов во время своей докторской диссертации, однако он не заметил важности концентрации ионов водорода и мутаротации глюкозы. Целью диссертации Анри было сравнить его знания о реакциях, катализируемых ферментами, с признанными законами физической химии.[2] Генри считается первым, кто написал уравнение, которое теперь известно как уравнение Михаэлиса-Ментен. Используя глюкозу и фруктозу в каталитических реакциях, контролируемых мальтазой и инвертазой, Леонор Михаэлис был первым ученым, который различил различные типы ингибирования с помощью шкалы pH, которой не было во времена Генри.[2]
В частности, во время своей работы по описанию скорости этой реакции они также протестировали и экстраполировали идею другого ученого: Виктор Анри, тот фермент, который они использовали, имел некоторое сродство к обоим продуктам этой реакции - фруктозе и глюкозе.[2][3] Используя методы Анри, Михаэлис и Ментен почти усовершенствовали эту концепцию метода начальной скорости для стационарных экспериментов. Они изучали торможение, когда обнаружили, что неконкурентное (смешанное) торможение характеризуется своим влиянием на kКот (скорость катализатора), в то время как конкурентоспособная, характеризуется своим влиянием на скорость (V).[2] В экспериментах Михаэлиса и Ментен они в значительной степени сосредоточились на влиянии инвертазы на pH с использованием ионов водорода.[2] Инвертаза - это фермент, обнаруженный во внеклеточных дрожжах и катализируемый реакциями путем гидролиза или превращения сахарозы (смеси сахарозы и фруктозы) в «инвертный сахар». Основная причина использования инвертазы заключалась в том, что ее можно было легко проанализировать, а эксперименты можно было проводить быстрее. Сахароза вращается в поляриметр в качестве правовращающие-D тогда как инвертный сахар левовращающие-Л. Это сделало отслеживание инверсии сахара относительно простым. Они также обнаружили, что α-D-глюкоза высвобождается в реакциях, катализируемых инвертазой, которая очень нестабильна и спонтанно превращается в β-D-глюкоза.[4] Хотя они оба находятся в правовращающей форме, именно здесь они отметили, что глюкоза может изменяться спонтанно, также известное как мутаротация. Неспособность принять это во внимание была одной из главных причин, по которой эксперименты Генри не оправдались. Используя инвертазу для катализа инверсии сахарозы, они смогли увидеть, насколько быстро реагирует фермент, с помощью поляриметрии; поэтому было обнаружено, что неконкурентное ингибирование происходит в реакции, в которой сахароза инвертировалась инвертазой.[2]
Терминология
Важно отметить, что, хотя все неконкурентные ингибиторы связывают фермент в аллостерических сайтах (т.е. в других местах, кроме его активный сайт ) - не все ингибиторы, которые связываются в аллостерических сайтах, являются неконкурентными ингибиторами.[1] Фактически, аллостерические ингибиторы могут действовать как конкурентный, неконкурентоспособный, или неконкурентоспособный ингибиторы.[1]
Многие источники продолжают объединять эти два термина,[5] или сформулируйте определение аллостерического ингибирования как определение неконкурентного ингибирования.
Механизм
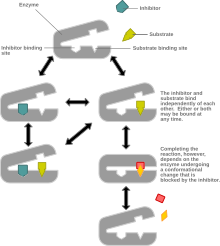
Неконкурентное ингибирование моделирует систему, в которой как ингибитор, так и субстрат могут быть связаны с ферментом в любой момент времени. Когда и субстрат, и ингибитор связаны, комплекс фермент-субстрат-ингибитор не может образовывать продукт и может быть преобразован только обратно в комплекс фермент-субстрат или комплекс фермент-ингибитор. Неконкурентное ингибирование отличается от общего смешанного ингибирования тем, что ингибитор имеет одинаковое сродство к ферменту и комплексу фермент-субстрат.
Например, в ферментативно-катализируемых реакциях гликолиз, накопление фосфоенола катализируется пируваткиназа в пируват. Аланин представляет собой аминокислоту, которая синтезируется из пирувата, также ингибирует фермент пируваткиназу во время гликолиза. Аланин - неконкурентный ингибитор, поэтому он связывается от активного центра с субстратом, чтобы он оставался конечным продуктом.[6]
Другой пример неконкурентного торможения дает глюкозо-6-фосфат ингибирование гексокиназы в головном мозге. Углероды 2 и 4 на глюкозо-6-фосфате содержат гидроксильные группы, которые присоединяются вместе с фосфатом на углероде 6 к комплексу фермент-ингибитор. Субстрат и фермент различаются по групповым комбинациям, к которым присоединяется ингибитор. Способность глюкозо-6-фосфата связываться в разных местах одновременно делает его неконкурентным ингибитором.[7]
Наиболее распространенный механизм неконкурентного ингибирования включает обратимое связывание ингибитора с аллостерический сайт, но ингибитор может действовать другими способами, включая прямое связывание с активным сайтом. Оно отличается от конкурентного ингибирования тем, что связывание ингибитора не предотвращает связывание субстрата и наоборот, а просто предотвращает образование продукта в течение ограниченного времени.

Этот тип торможения снижает максимальная ставка из химическая реакция без изменения видимой привязки близость из катализатор для субстрат (Kмприложение - видеть Кинетика Михаэлиса-Ментен ). Когда добавляется неконкурентный ингибитор, Vmax изменяется, а Km остается неизменным. Согласно Заговор Лайнуивера-Берка Vmax уменьшается во время добавления неконкурентного ингибитора, что показано на графике изменением наклона кривой и пересечения по оси y при добавлении неконкурентного ингибитора.[8]
Основное различие между конкурентным и неконкурентным является то, что конкурентное ингибирование влияет на способность субстрата связываться путем связывания ингибитора вместо субстрата, что снижает сродство фермента к субстрату. При неконкурентном ингибировании ингибитор связывается с аллостерическим сайтом и предотвращает выполнение химической реакции комплексом фермент-субстрат. Это не влияет на Km (сродство) фермента (к субстрату). Неконкурентное ингибирование отличается от неконкурентного ингибирования тем, что оно все еще позволяет субстрату связываться с комплексом фермент-ингибитор и образовывать комплекс фермент-субстрат-ингибитор, это неверно в случае неконкурентного ингибирования, оно предотвращает связывание субстрата с ферментом. ингибитор через конформационное изменение при аллостерическом связывании.
Уравнение
В присутствии неконкурентного ингибитора кажущееся сродство к ферменту эквивалентно фактическому сродству. С точки зрения Кинетика Михаэлиса-Ментен, Kмприложение = Kм. Это можно рассматривать как следствие Принцип Ле Шателье потому что ингибитор одинаково связывается как с ферментом, так и с комплексом фермент-субстрат, так что равновесие сохраняется. Однако, поскольку некоторые ферменты всегда препятствуют превращению субстрата в продукт, эффективная концентрация фермента снижается.
Математически,
![V _ {{max}} ^ {{app}} = { frac {V _ {{max}}} {1 + { frac {[I]} {K_ {I}}}}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/1b945d1d3deff510520bdb20f9865068d4d12c4d)
![{кажущийся [E] _ {0}} = { frac {[E] _ {0}} {1 + { frac {[I]} {K_ {I}}}}}](https://wikimedia.org/api/rest_v1/media/math/render/svg/b5dc3a4908e8409124b361d5e495f6d2f54f636e)
Пример: неконкурентные ингибиторы фермента CYP2C9.
Неконкурентные ингибиторы CYP2C9 фермент включают нифедипин, транилципромин, фенэтилизотиоцианат и 6-гидроксифлавон. Компьютерное моделирование стыковки и замененные сконструированные мутанты показывают, что неконкурентный сайт связывания 6-гидроксифлавона является известным сайтом аллостерического связывания CYP2C9 фермент.[9]
Рекомендации
- ^ а б c d Джон Стрелоу, Уолтер Дью, Филип Иверсен, Гарольд Брукс, Джеффри Рэддинг, Джеймс МакГи и Джеффри Вайднер, «Анализ механизма действия ферментов», в Дж. С. Ситтампаламе, Н. П. Кусенс, Х. Нельсон и другие. (редакторы), Руководство по проведению анализа, Eli Lilly & Company и Национальный центр развития переводческих наук, 2004 г.
- ^ а б c d е ж грамм час я Корниш-Боуден, Атель (2015-03-01). «Сто лет кинетики Михаэлиса – Ментен». Перспективы в науке. Материалы симпозиума Beilstein ESCEC - Празднование 100-летия Михаэлиса Ментен-Кинетикс. 4 (Дополнение C): 3–9. Дои:10.1016 / j.pisc.2014.12.002.
- ^ а б Michaelis, L .; Ментен, мисс Мод Л. (2 сентября 2013 г.). «Кинетика действия инвертина». Письма FEBS. 587 (17): 2712–2720. Дои:10.1016 / j.febslet.2013.07.015. ISSN 1873-3468. PMID 23867202. S2CID 43226286.
- ^ а б c Корниш-Боуден, Атель (2 сентября 2013 г.). «Истоки ферментативной кинетики». Письма FEBS. Кинетика века Михаэлиса - Ментен. 587 (17): 2725–2730. Дои:10.1016 / j.febslet.2013.06.009. PMID 23791665. S2CID 12573784.
- ^ «Неконкурентное торможение и аллостерическое торможение». Биология онлайн (форум). Архивировано из оригинал 25 апреля 2015 г.. Получено 2 апреля 2012.
- ^ Берг, Джереми М .; Тимочко, Джон Л .; Страйер, Люберт (2002). «Гликолитический путь жестко контролируется». Биохимия. 5-е издание.
- ^ Крейн, Роберт; Сольс, Альберто (8 февраля 1954 г.). «Неконкурентное ингибирование гексокиназы головного мозга глюкозой-6-фосфатом и родственными соединениями *» (PDF). Департамент биологической химии Медицинской школы Вашингтонского университета. Получено 3 ноября, 2017.
- ^ Уолдроп, Г. Л. (22 января 2009 г.). Качественный подход к ингибированию ферментов. Получено 31 октября 2017 г. из http://onlinelibrary.wiley.com/doi/10.1002/bmb.20243/pdf
- ^ Си, Дайонг; Ван, Инь; Чжоу И-Хан; Го, Инцзе; Ван, Хуан; Чжоу, Хуэй; Ли, Цзэ-Шэн; Фосетт, Дж. Пол (12 декабря 2008 г.). «Механизм ингибирования CYP2C9 флавонами и флавонолами». Метаболизм и утилизация лекарств. Американское общество фармакологии и экспериментальной терапии (ASPET). 37 (3): 629–634. Дои:10.1124 / dmd.108.023416. ISSN 0090-9556. [1]