Фотосинтетический реакционный центр - Photosynthetic reaction centre - Wikipedia

А фотосинтетический реакционный центр представляет собой комплекс нескольких белков, пигментов и других кофакторов, которые вместе выполняют первичные реакции преобразования энергии фотосинтез. Молекулярные возбуждения, возникающие либо непосредственно от солнечного света, либо передающиеся в виде энергия возбуждения через светособирающие антенные системы, породить перенос электронов реакции на пути ряда связанных с белками кофакторов. Эти кофакторы представляют собой светопоглощающие молекулы (также называемые хромофоры или же пигменты ) Такие как хлорофилл и феофитин, а также хиноны. Энергия фотон используется для возбуждения электрон пигмента. В свободная энергия created затем используется для сокращения цепочки ближайших акцепторы электронов, у которых все больше редокс-потенциалы. Эти перенос электронов Шаги - это начальная фаза ряда реакций преобразования энергии, в конечном итоге приводящих к преобразованию энергии фотонов в накопление этой энергии путем образования химических связей.
Преобразование световой энергии в разделение зарядов
Центры реакции присутствуют во всем зеленом растения, водоросли, и много бактерии. Разнообразие в светоуборочные комплексы существуют между фотосинтезирующими видами. Зеленые растения и водоросли имеют два разных типа реакционных центров, которые являются частью более крупных суперкомплексов, известных как P700 в фотосистема I и P680 в фотосистема II. Структуры этих суперкомплексов имеют большие размеры и включают в себя несколько светоуборочные комплексы. Центр реакции найден в Rhodopseudomonas бактерии в настоящее время изучены лучше всего, так как это был первый реакционный центр известной структуры и полипептидные цепи чем примеры с зелеными растениями.[1]
Реакционный центр устроен таким образом, что он захватывает энергию фотона с помощью молекул пигмента и превращает ее в пригодную для использования форму. Как только световая энергия была поглощена непосредственно молекулами пигмента или передана им через резонансный перенос из окружения светоуборочный комплекс, они выпускают два электроны в электронная транспортная цепь. У зеленых растений в цепи переноса электронов есть много акцепторов электронов, включая феофитин, хинон, пластохинон, цитохром bf, и ферредоксин, что в итоге приводит к восстановленной молекуле НАДФН и хранение энергии. Прохождение электрона через цепь переноса электронов также приводит к накачке протоны (ионы водорода) из хлоропласт с строма и в просвет, что приводит к протонному градиенту по тилакоидная мембрана которые можно использовать для синтеза АТФ с использованием АТФ-синтаза молекула. И АТФ, и НАДФН используются в Цикл Кальвина фиксировать углекислый газ в триозные сахара.
В бактериях
Классификация
Различают два класса реакционных центров. Тип I, найденный в зелено-серные бактерии, Гелиобактерии и PS-I растений / цианобактерий используют кластеры серы железа в качестве акцепторов электронов. Тип II, обнаруженный в хлорофлексус, пурпурные бактерии и PS-II растений / цианобактерий используют хиноны. Мало того, что все члены внутри каждого класса имеют общее происхождение, эти два класса также кажутся связанными посредством общей структуры.[2][3] В этом разделе рассматривается система типа II, обнаруженная у пурпурных бактерий.
Структура

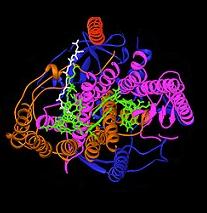
Центр реакции бактериального фотосинтеза был важной моделью для понимания структуры и химии биологического процесса улавливания световой энергии. В 1960-е гг. Родерик Клейтон был первым, кто очистил комплекс реакционного центра от пурпурных бактерий. Однако первая кристаллическая структура (верхнее изображение справа) была определена в 1984 г. Хартмут Мишель, Иоганн Дайзенхофер и Роберт Хубер[4] для чего они разделили Нобелевская премия в 1988 г.[5] Это было также важно для того, чтобы быть первой трехмерной кристаллической структурой любого мембранного белкового комплекса.
Было обнаружено, что четыре различные субъединицы важны для функции фотосинтетического реакционного центра. Субъединицы L и M, показанные синим и фиолетовым на изображении конструкции, оба охватывают липидный бислой плазматической мембраны. Они структурно похожи друг на друга, оба имеют 5 трансмембранных альфа спирали.[6] Четыре бактериохлорофилл b (БХл-b) молекулы, две бактериофеофитин b молекулы (BPh) молекулы, две хиноны (QА и QB), а ион двухвалентного железа связан с L- и M-субъединицами. Субъединица H, показанная золотом, находится на цитоплазматической стороне плазматической мембраны. Субъединица цитохрома, здесь не показана, содержит четыре гема c-типа и расположена на периплазматической поверхности (внешней) мембраны. Последняя субъединица не является общим структурным мотивом фотосинтезирующих бактерий. Субъединицы L и M связывают функциональные и взаимодействующие со светом кофакторы, показанные здесь зеленым.
Центры реакции из разных видов бактерий могут содержать слегка измененные хромофоры бактериохлорофилла и бактериофеофитина в качестве функциональных кофакторов. Эти изменения вызывают изменение цвета света, который может быть поглощен. В реакционном центре находятся два пигмента, которые служат для сбора и передачи энергии от поглощения фотонов: BChl и Bph. БХл примерно напоминает молекулу хлорофилла, обнаруженную в зеленых растениях, но из-за незначительных структурных отличий длина волны его пика поглощения смещена в инфракрасный, с длинами волн до 1000 нм. Bph имеет ту же структуру, что и BChl, но центральный ион магния заменен двумя протонами. Это изменение вызывает как сдвиг максимума поглощения, так и пониженный окислительно-восстановительный потенциал.
Механизм

Процесс начинается, когда свет поглощается двумя молекулами БХл, лежащими вблизи периплазматический сторона мембраны. Эта пара молекул хлорофилла, часто называемая «особой парой», поглощает фотоны с длиной волны 870 или 960 нм, в зависимости от вида, и поэтому называется P870 (для Rhodobacter sphaeroides ) или P960 (для Blastochloris viridis ), с п означает «пигмент»). Как только P поглощает фотон, он выбрасывает электрон, который передается через другую молекулу Bchl к BPh в субъединице L. Это начальное разделение зарядов дает положительный заряд на P и отрицательный заряд на BPh. Этот процесс занимает 10 пикосекунд (10−11 секунд).[1]
Обвинения по П+ и BPh− в этом состоянии может происходить рекомбинация заряда. Это приведет к потере электронов высокой энергии и преобразованию поглощенной световой энергии в высокая температура. Этому способствует несколько факторов структуры реакционного центра. Во-первых, перенос электрона с БП− к P960+ относительно медленный по сравнению с двумя другими окислительно-восстановительные реакции в реакционном центре. Более быстрые реакции включают перенос электрона от BPh− (BPh− окисляется до BPh) до хинона-акцептора электронов (QА), а перенос электрона на P960+ (P960+ восстанавливается до P960) из гема в субъединице цитохрома над реакционным центром.
Электрон высокой энергии, находящийся на прочно связанной молекуле хинона QА передается на заменяемую молекулу хинона QB. Эта молекула слабо связана с белком и довольно легко отделяется. Два высокоэнергетических электрона необходимы для полного уменьшения QB в QH2, забирая при этом два протона из цитоплазмы. Восстановленный хинон QH2 диффундирует через мембрану к другому белковому комплексу (цитохром bc1-сложный ), где он окисляется. В процессе снижения мощности QH2 используется для перекачки протонов через мембрану в периплазматическое пространство. Электроны цитохрома bc1-комплекс затем переносится через растворимый промежуточный цитохром с, называемый цитохромом с2в периплазме к субъединице цитохрома.
У цианобактерий и растений
Цианобактерии, предшественники хлоропласты найденные в зеленых растениях, имеют обе фотосистемы с обоими типами реакционных центров. Объединение двух систем позволяет производить кислород.
Кислородный фотосинтез
В 1772 г. химик Джозеф Пристли провел серию экспериментов, касающихся газов, участвующих в дыхании и горении. В своем первом эксперименте он зажег свечу и поместил ее под перевернутый сосуд. Спустя небольшой промежуток времени свеча перегорела. Он провел аналогичный эксперимент с мышь в замкнутом пространстве горящей свечи. Он обнаружил, что мышь умерла вскоре после того, как погасла свеча. Однако он мог оживить зловонный воздух, поместив зеленые растения в этом районе и выставив их на свет. Наблюдения Пристли были одними из первых экспериментов, которые продемонстрировали активность фотосинтетического реакционного центра.
В 1779 г. Ян Ингенхауз провел более 500 экспериментов в течение 4 месяцев, пытаясь понять, что происходит на самом деле. Он описал свои открытия в книге под названием Эксперименты с овощами. Ингенхауз взял зеленые растения и погрузил их в воду в прозрачном резервуаре. Он наблюдал множество пузырей, поднимающихся с поверхности листьев, когда растения подвергались воздействию света. Ингенхауз собрал газ, выделяемый заводами, и провел несколько различных тестов, пытаясь определить, что это за газ. Испытание, которое, наконец, выявило идентичность газа, заключалось в помещении тлеющего конуса в образец газа и его повторном зажигании. Этот тест доказал, что это кислород или, как назвал его Джозеф Пристли, «де-флогистированный воздуха'.
В 1932 г. Роберт Эмерсон и его ученик Уильям Арнольд использовали метод повторяющихся вспышек для точного измерения небольших количеств кислорода, выделяемого хлорофиллом в водорослях. Хлорелла. Их эксперимент подтвердил существование фотосинтетической единицы. Позже Гаффрон и Воль интерпретировали эксперимент и поняли, что свет, поглощаемый фотосинтетическим устройством, передается.[7] Эта реакция происходит в реакционном центре фотосистемы II и имеет место у цианобактерий, водорослей и зеленых растений.[8]
Фотосистема II

Фотосистема II фотосистема, которая генерирует два электрона, которые в конечном итоге уменьшают НАДФ+ в ферредоксин-НАДФ-редуктазе. Фотосистема II присутствует на мембранах тилакоидов внутри хлоропластов, месте фотосинтеза у зеленых растений.[9] Структура фотосистемы II удивительно похожа на бактериальный реакционный центр, и предполагается, что у них есть общий предок.
Ядро фотосистемы II состоит из две субъединицы, обозначаемые как D1 и D2. Эти две субъединицы подобны субъединицам L и M, присутствующим в бактериальном реакционном центре. Фотосистема II отличается от бактериального реакционного центра тем, что в ней есть много дополнительных субъединиц, которые связывают дополнительные хлорофиллы для повышения эффективности. Общая реакция катализированный по фотосистеме II:
- 2Q + 2H2O + hν → O2 + 2QH2
Q представляет собой пластохинон, окисленную форму Q. QH2 представляет собой пластохинол, восстановленную форму Q. Этот процесс восстановления хинона сравним с тем, который имеет место в бактериальном реакционном центре. Фотосистема II получает электроны путем окисления воды в процессе, называемом фотолиз. Молекулярный кислород является побочным продуктом этого процесса, и именно эта реакция обеспечивает атмосфера с кислородом. Тот факт, что кислород зеленых растений поступает из воды, был впервые сделан американским биохимиком канадского происхождения. Мартин Дэвид Камен. Он использовал стабильный изотоп кислорода, 18O, чтобы проследить путь кислорода от воды до газообразного молекулярного кислорода. Эта реакция катализируется реакционным центром в фотосистеме II, содержащей четыре марганец ионы.

Реакция начинается с возбуждения пары молекул хлорофилла, аналогичных молекулам бактериального реакционного центра. Из-за наличия хлорофилла а, в отличие от бактериохлорофилл, фотосистема II поглощает свет с более короткой длиной волны. Пару молекул хлорофилла в реакционном центре часто называют P680.[1] Когда фотон поглощается, полученный электрон высокой энергии передается соседней молекуле феофитина. Он находится вверху и справа от пары на диаграмме и окрашен в серый цвет. Электрон проходит от молекулы феофитина через две молекулы пластохинона, первая из которых тесно связана, а вторая - слабо. Плотно связанная молекула показана над молекулой феофитина и окрашена в красный цвет. Слабосвязанная молекула находится слева от нее и тоже окрашена в красный цвет. Этот поток электронов подобен потоку бактериального реакционного центра. Два электрона необходимы для полного восстановления слабосвязанной молекулы пластохинона до QH.2 а также поглощение двух протонов.
Разница между фотосистемой II и бактериальным реакционным центром заключается в источнике электрона, нейтрализующего пару хлорофилла. а молекулы. В бактериальном реакционном центре электрон получается из группы восстановленного соединения гема в субъединице цитохрома или из водорастворимого белка цитохрома-c.
Один раз фотоиндуцированное разделение зарядов молекула P680 несет положительный обвинять. P680 очень сильный окислитель и извлекает электроны из двух воды молекулы, которые связаны в центре марганца непосредственно под парой. Этот центр ниже и слева от пары на диаграмме содержит четыре иона марганца: кальций ион, а хлористый ион, а тирозин остаток. Марганец хорошо подходит для этих реакций, потому что он способен существовать в четырех степенях окисления: Mn2+, Mn3+, Mn4+ и Mn5+. Марганец также образует прочные связи с кислородсодержащими молекулами, такими как вода.
Каждый раз, когда P680 поглощает фотон, он излучает электрон, приобретая положительный заряд. Этот заряд нейтрализуется извлечением электрона из марганцевого центра, который находится прямо под ним. Процесс окисления двух молекул воды требует четырех электронов. Молекулы воды, которые окисляются в центре марганца, являются источником электронов, которые восстанавливают две молекулы Q до QH.2. На сегодняшний день этот каталитический центр расщепления воды не воспроизводится ни одним искусственным катализатором.
Фотосистема I
После того, как электрон покинул фотосистему II, он переходит в комплекс цитохрома b6f а затем в пластоцианин, синий медь белок и переносчик электронов. Комплекс пластоцианина несет электрон, который нейтрализует пару в следующем реакционном центре, фотосистема I.
Как и в случае фотосистемы II и бактериального реакционного центра, пара хлорофилла а молекулы инициируют фотоиндуцированное разделение зарядов. Эта пара называется P700. 700 - это ссылка на длина волны при котором молекулы хлорофилла максимально поглощают свет. P700 находится в центре белка. Как только фотоиндуцированное разделение зарядов инициировано, электрон перемещается по пути через молекулу хлорофилла α, расположенную непосредственно над P700, через молекулу хинона, расположенную непосредственно над ней, через три кластера 4Fe-4S и, наконец, к взаимозаменяемому комплексу ферредоксина.[10] Ферредоксин представляет собой растворимый белок, содержащий кластер 2Fe-2S, координированный четырьмя остатками цистеина. Положительный заряд, оставшийся на P700, нейтрализуется переносом электрона из пластоцианин. Таким образом, общая реакция, катализируемая фотосистемой I, такова:
- ПК (Cu+) + Fd[бык] + hν → Pc (Cu2+) + Fd[красный]
Сотрудничество между фотосистемами I и II создает поток электронов из H2O в NADP+. Этот путь называется 'Z-схема ' поскольку редокс диаграмма от P680 до P700 напоминает букву z.[11]
Смотрите также
- Светоуборочный комплекс
- Фотосинтез
- Фотографиисистема
- Фикобилисома
- Семейство белков фотосинтетического реакционного центра
Рекомендации
- ^ а б c Берг Дж. М., Тимочко Дж. Л., Страйер Л. (2002). «Глава 19: Световые реакции фотосинтеза». Биохимия (5-е изд.). Нью-Йорк: WH Freeman.
- ^ Садекар, S; Раймонд, Дж; Бланкеншип, RE (ноябрь 2006 г.). «Сохранение отдаленно родственных мембранных белков: центры фотосинтетических реакций имеют общее структурное ядро». Молекулярная биология и эволюция. 23 (11): 2001–7. Дои:10.1093 / molbev / msl079. PMID 16887904.
- ^ Орф Г.С., Гизриэль С., Реддинг К.Э. (октябрь 2018 г.). «Эволюция фотосинтетических реакционных центров: понимание структуры реакционного центра гелиобактерий». Фотосинтез Исследования. 138 (1): 11–37. Дои:10.1007 / s11120-018-0503-2. OSTI 1494566. PMID 29603081. S2CID 4473759.
- ^ Дайзенхофер Дж., Эпп О, Мики К., Хубер Р., Мишель Х (декабрь 1984 г.). «Рентгеноструктурный анализ мембранного белкового комплекса. Карта электронной плотности при разрешении 3 A и модель хромофоров фотосинтетического реакционного центра из Rhodopseudomonas viridis». Журнал молекулярной биологии. 180 (2): 385–98. Дои:10.1016 / S0022-2836 (84) 80011-X. PMID 6392571.
- ^ "Нобелевская премия по химии 1988 г.". NobelPrize.org.
- ^ Фотосинтетические реакционные центры пурпурных бактерий В архиве 2006-05-14 на Wayback Machine (2 февраля 1999 г.). Проверено 28 февраля 2010 года.
- ^ Говинджи Р. (2000). "Глава 1: Вехи фотосинтеза". В Юнус М., Патре У, Моханти П. (ред.). Зондирование фотосинтеза: механизмы, регуляция и адаптация. Лондон: Тейлор и Фрэнсис. ISBN 978-0-7484-0821-4.
- ^ Kaiser GE (24 февраля 2003 г.). «Кислородный фотосинтез: рост бактерий и микробный метаболизм». Архивировано из оригинал 4 мая 2006 г.
- ^ «Хлоропласт». Ультранет Биология. Архивировано из оригинал 3 августа 2003 г.
- ^ Джаганнатан Б., Голбек Дж. (2009). «Фотосинтез: микробный». В Schaechter M (ред.). Энциклопедия микробиологии (3-е изд.). С. 325–341. Дои:10.1016 / B978-012373944-5.00352-7. ISBN 978-0-12-373944-5.
- ^ Говинджи Р. «Z-схема фотосинтеза». Университет Иллинойса в Урбане-Шампейн.
внешняя ссылка
- Орр Л., Говинджи Р. "Веб-ресурсы фотосинтеза". Иллинойсский университет в Урбане-Шампейн.