Cyathus - Cyathus
| Cyathus | |
|---|---|
 | |
| Cyathus striatus | |
| Научная классификация | |
| Королевство: | |
| Разделение: | |
| Учебный класс: | |
| Заказ: | |
| Семья: | |
| Род: | Cyathus Халлер (1768) |
| Типовой вид | |
| Cyathus striatus | |
| Разновидность | |
| Cyathus | |
|---|---|
| glebal гимений | |
| колпачок является воронкообразный | |
| гимений вложение не применимо | |
| не хватает ножка | |
| экология сапротрофный | |
| съедобность: несъедобный | |
Cyathus это род из грибы в Nidulariaceae, а семья все вместе известные как грибы птичьего гнезда. Им дано это название, потому что они напоминают крошечные птичьи гнезда, заполненные «яйцами», - структуры, достаточно большие, чтобы их принимали в прошлом за семена. Однако теперь известно, что это репродуктивные структуры, содержащие споры. «Яйца», или перидиолы, прочно прикреплены к внутренней поверхности этого плодовое тело эластичным шнуром из мицелия известный как жаберник. В 45 видов широко распространены по всему миру, некоторые из них встречаются в большинстве стран, хотя некоторые существуют только в одном или двух регионах. Cyathus stercoreus Считается находящихся под угрозой исчезновения в ряде европейских стран. Виды Cyathus также известны как брызги, который относится к тому факту, что падающие капли дождя могут выбить перидиолы из плодового тела открытой чаши. Внутренняя и внешняя поверхности этой чашки могут иметь продольные гребни (обозначаемые как складчатый или же полосатый ); это один из примеров таксономический характеристика, которая традиционно служила для различения видов.
Обычно считается несъедобным, Cyathus виды сапробный, так как они получают питательные вещества от разложения органическая материя. Обычно они растут на гниющей древесине или древесных остатках, на коровьем и конском навозе или непосредственно на перегной -плодородная земля. В жизненный цикл этого рода позволяет ему воспроизводиться как половым путем, с мейоз, и бесполым путем через споры. Несколько Cyathus виды производят биоактивный соединения, некоторые с лечебными свойствами, а некоторые лигнин -унижение ферменты из рода могут быть полезны в биоремедиация и сельское хозяйство. Филогенетический анализ дает новое понимание эволюционный отношения между различными видами в Cyathus, и поставил под сомнение обоснованность более раннего классификация системы, основанные на традиционных таксономических характеристиках
История
Грибы птичьих гнезд были впервые упомянуты фламандским ботаником. Карол Клузиус в Rariorum plantarum история (1601). В течение следующих нескольких столетий эти грибы были предметом споров относительно того, были ли перидиолы семенами, и механизма, с помощью которого они были распространены в природе. Например, французский ботаник Жан-Жак Поле, в его работе Traité des шампиньоны (1790–173) предложили ошибочное представление о том, что перидиолы выбрасываются из плодовых тел каким-то пружинным механизмом.[2] Род был установлен в 1768 году швейцарским ученым. Альбрехт фон Галлер; в родовое имя Cyathus является латинский, но изначально произошел от Древнегреческий слово κύαθος, означающее «чаша».[3] Строение и биология рода Cyathus был более известен к середине 19 века, начиная с появления в 1842 году работы Карла Иоганна Фридриха Шмитца,[4] а два года спустя монография братьями Луи Рене и Чарльз Туласне.[5] Работа Туласнов была тщательной и точной и получила высокую оценку более поздних исследователей.[2][6][7] Впоследствии монографии были написаны в 1902 году Вайолет С. Уайт (об американских видах),[6] Кертис Гейтс Ллойд в 1906 г.,[7] Гордон Хэрриот Каннингем в 1924 г. (по новозеландским видам),[8] и Гарольд Дж. Броди в 1975 г.[9]
Описание
Виды в роду Cyathus есть плодовые тела (перидия ), которые имеют форму вазы, трубы или урны размером 4–8 мм в ширину и 7–18 мм в высоту.[10] Плодовые тела имеют цвет от коричневого до серо-коричневого, на внешней поверхности покрыты мелкими волосковидными структурами. Некоторые виды, например С. striatus и C. setosus, имеют на краю чашечки заметные щетинки, называемые щетинками. Плодовое тело часто расширяется у основания в твердую округлую массу из гифы так называемая установка, которая обычно запутывается и опутывается небольшими фрагментами подстилающей растущей поверхности, улучшая ее устойчивость и помогая не быть опрокинутой дождем.[11]

Незрелые плодовые тела имеют белесую оболочку, эпифрагму, покрывающую перидиум открытие в молодости, но со временем расшифровки, раскрывающиеся во время созревания. Под микроскопом перидий Cyathus Вид состоит из трех отдельных слоев - эндо-, мезо- и эктоперидиума, относящихся к внутреннему, среднему и внешнему слоям соответственно. В то время как поверхность эктоперидиума в Cyathus обычно волосистая, эндоперидиальная поверхность гладкая, в зависимости от вида может иметь продольные бороздки (бороздки).[3]
Поскольку основная структура плодового тела во всех роды семейства Nidulariaceae по существу похожи, Cyathus можно легко спутать с видами Нидула или же Crucibulum, особенно старые, выветрившиеся экземпляры Cyathus это может привести к стиранию волосатого эктоперидиума.[12] Он отличался от Нидула наличием семенного канатика, связки гиф, прикрепляющей перидиолу к эндоперидиуму. Cyathus отличается от рода Crucibulum за счет отчетливой трехслойной стенки и более сложного семенного канатика.[3]
Структура перидиола

Произведено от греческого слова перидион, что означает "маленький кожаный мешочек",[13] перидиол - «яйцо» птичьего гнезда. Это масса базидиоспоры и glebal ткань окружена твердой восковой оболочкой. Форма может быть описана как линзовидная - как двояковыпуклая линза - и в зависимости от вида может иметь цвет от беловатого до сероватого или черного. Внутренняя камера перидиолы содержит гимений это сделано из базидия, стерильные (не репродуктивные) структуры и споры. В молодых, только что открывшихся плодовых телах перидиолы находятся в прозрачном студенистом веществе, которое вскоре высыхает.[14]
Перидиолы прикреплены к плодовому телу семенным канатиком, сложной структурой из гифы его можно разделить на три области: базальная часть, которая прикрепляет ее к внутренней стенке перидиума, средняя часть и верхняя оболочка, называемая кошельком, соединенная с нижней поверхностью перидиола. В сумочке и средней части находится спиральная нить переплетенных гиф, называемая фуникулярным шнуром, прикрепленная одним концом к перидиоле, а другим концом к запутанной массе гиф, называемой хаптероном. У некоторых видов перидиолы могут быть покрыты оболочкой - тонкой белой мембраной (особенно заметно у С. striatus и C. crassimurus).[15]Споры обычно имеют эллиптическую или примерно сферическую форму и толстостенные, гиалиновый или светло-желто-коричневого цвета, размерами 5–15 на 5–8мкм.[10]
Среда обитания и распространение
Плодовые тела обычно растут гроздьями и обнаруживаются на мертвой или гниющей древесине или на древесных фрагментах коровьего или конского навоза.[10] К навозолюбивым (копрофильным) видам относятся: С. stercoreus, C. costatus, C. fimicola, и C. pygmaeus.[16] Некоторые виды были собраны на древесном материале, как мертвые травянистый стебли, пустые оболочки или шелуха орехов или волокнистых материалов, например кокос, джут, или же конопля волокна, вплетенные в циновки, мешки или ткань.[17] В природе плодовые тела обычно встречаются во влажных, частично затененных местах, например, на опушках лесов на тропах или вокруг освещенных проемов в лесах. Они реже встречаются среди густой растительности и глубоких мхов, поскольку в этих средах они могут препятствовать распространению перидиолей падающими каплями воды.[18] Внешний вид плодовых тел во многом зависит от особенностей окружающей среды, в которой они произрастают непосредственно; в частности, оптимальные условия температуры, влажности и наличия питательных веществ являются более важными факторами для фруктов, а не географическая территория, в которой расположены грибы, или время года.[18] Примеры умения Cyathus для процветания в несколько негостеприимной среде обеспечиваются С. striatus и С. stercoreus, которые могут пережить засуху и зимние холода в умеренном климате Северной Америки,[19] и виды C. helenae, который был найден растущим на мертвых альпийские растения на высоте 7000 футов (2100 м).[20]

В целом виды Cyathus иметь всемирный распределение, но редко встречаются в Арктический и субарктический.[3] Один из самых известных видов, С. striatus имеет циркумполярное распространение и обычно встречается повсюду умеренный локации, в то время как морфологически похожие C. poeppigii широко распространен в тропический районы, редко в субтропики и никогда в регионах с умеренным климатом.[21] Большинство видов обитают в теплом климате. Например, хотя в США и Канаде зарегистрировано 20 различных видов, обычно встречаются только 8; с другой стороны, 25 видов могут регулярно встречаться в Вест-Индии, а Гавайские острова только есть 11 видов.[22] Некоторые виды кажутся эндемичный в определенные регионы, такие как С. novae-zeelandiae нашел в Новая Зеландия, или же C. crassimurus, встречается только на Гавайях; однако этот очевидный эндемизм может быть просто результатом недостатка коллекций, а не различием в среде обитания, которое представляет собой препятствие для распространения.[22] Хотя широко распространен в тропиках и большей части умеренного мира, С. stercoreus редко встречается в Европе; это привело к его появлению на ряде Красные списки. Например, считается находящихся под угрозой исчезновения в Болгария,[23] Дания,[24] и Черногория,[25] и "почти под угрозой" в Великобритания.[26] Открытие Cyathus виды в Доминиканский янтарь (C. dominicanus ) предполагает, что основная форма грибов птичьего гнезда уже сформировалась Меловой эры и что группа диверсифицировалась к серединеКайнозойский.[27]
Жизненный цикл
Жизненный цикл рода Cyathus, который содержит как гаплоидный и диплоид этапов, типично для таксоны в базидиомицеты которые могут воспроизводить как бесполым путем (через вегетативный споры) или половым путем (с мейоз ). Как и у других древесных грибов, этот жизненный цикл можно рассматривать как две функционально разные фазы: вегетативную стадию распространения мицелия и репродуктивная стадия для создания спорообразующих структур - плодовых тел.[28]
Вегетативная стадия включает в себя те фазы жизненного цикла, которые связаны с прорастанием, распространением и выживанием мицелия. Споры прорастают при подходящих условиях влажности и температуры и превращаются в ветвящиеся нити, называемые гифы, уходя корнями в гниющее дерево. Эти гифы гомокариотический, содержащий один ядро в каждом отсеке; они увеличиваются в длине за счет добавления материала клеточной стенки к растущему кончику. По мере того, как эти советы расширяются и распространяются, создавая новые точки роста, развивается сеть, называемая мицелием. Рост мицелия происходит митоз и синтез биомассы гиф. Когда две гомокариотические гифы разных группы совместимости спаривания сливаются друг с другом, они образуют дикариотический мицелий в процессе, называемом плазмогамия. Предпосылки для выживания мицелия и заселения субстрата (например, гниющей древесины) включают подходящую влажность и доступность питательных веществ. Большая часть чего-либо Cyathus виды сапробный, поэтому рост мицелия в гниющей древесине становится возможным благодаря секреции ферменты которые ломают комплекс полисахариды (Такие как целлюлоза и лигнин ) в простые сахара, которые можно использовать в качестве питательных веществ.[29]
По прошествии определенного периода времени и в соответствующих условиях окружающей среды дикариотический мицелий может перейти в репродуктивную стадию жизненного цикла. На формирование тела плода влияют такие внешние факторы, как время года (которое влияет на температуру и влажность воздуха), питательные вещества и свет. По мере развития плодовых тел они производят перидиолы, содержащие базидия на котором образуются новые базидиоспоры. Молодые базидии содержат пару гаплоидных сексуально совместимых ядер, которые сливаются, и образующееся диплоидное слияние ядер подвергается мейозу с образованием базидиоспоры, каждый из которых содержит одно гаплоидное ядро.[30] Дикариотический мицелий, из которого производятся плодовые тела, является долговечным и будет продолжать производить последующие поколения плодовых тел, пока благоприятные условия окружающей среды.

Развитие Cyathus плодовые тела изучены в лабораторных условиях; Cyathus stercoreus наиболее часто использовался для этих исследований из-за легкости, с которой его можно выращивать экспериментально.[31] В 1958 году Э. Гарнетт впервые продемонстрировал, что развитие и форма плодовых тел хотя бы частично зависят от интенсивности света, получаемого во время развития. Например, экспонирование гетерокариотический мицелий чтобы зажечь плоды, и, кроме того, этот свет должен быть длина волны менее 530 нм. Постоянный свет не требуется для развития плодового тела; после того, как мицелий достигнет определенной стадии зрелости, необходимо лишь кратковременное воздействие света, и плодовые тела образуются, даже если впоследствии их держать в темноте.[32] Лу предположил в 1965 году, что определенные условия выращивания - такие как нехватка доступных питательных веществ - сдвигают грибок. метаболизм создать гипотетический «фоторецептивный предшественник», который позволяет стимулировать рост плодовых тел и воздействовать на них светом.[33] Грибки тоже положительно фототрофный, то есть он будет ориентировать свои плодовые тела в направлении источника света.[34] Время, необходимое для развития плодовых тел, зависит от ряда факторов, таких как температура или доступность и тип питательных веществ, но в целом «большинство видов, которые действительно плодоносят в лабораторных условиях, лучше всего себя чувствуют при температуре около 25 ° C. От 18 до 40 дней ".[35]
Распространение спор
Как и другие грибы птичьих гнезд в Nidulariaceae, виды Cyathus их споры рассеиваются, когда вода попадает в плодовое тело. Плодовое тело имеет такую форму, что кинетическая энергия упавшей капли перенаправляется вверх и немного наружу за счет угла стенки чашки, который составляет 70–75 ° по отношению к горизонтали.[36] Действие выталкивает перидиолы из так называемой «чашки для брызг», где они могут разбить и распространить споры внутри или быть съеденными и рассеяными животными после прохождения через пищеварительный тракт. Этот метод распространения спор в Nidulariaceae был экспериментально испытан Джорджем Уиллардом Мартином в 1924 г.[37] и позже разработал Артур Генри Реджинальд Буллер, который использовал Cyathus striatus в качестве модельного вида для экспериментального исследования явления.[38] Основные выводы Буллера относительно распространения спор были позже резюмированы его аспирантом Гарольдом Дж. Броди, с которым он провел несколько из этих экспериментов с брызговиками:
Капли дождя заставляют перидиолы Nidulariaceae разбрызгиваться примерно на четыре фута. В роду Cyathus, когда перидиол вырывается из чашки, семенной канатик разрывается, и это делает возможным расширение массы липких гиф (хаптерон), которые цепляются за любой объект на линии полета. Импульс перидиолы заставляет длинный шнур вытягиваться из оболочки, прикрепленной к перидиоле. Перидиол проверяется в полете, и толчок заставляет фуникулярный шнур наматываться на стебли или запутываться среди растительных волосков. Таким образом, перидиол прикрепляется к растительности и впоследствии может быть съеден травоядными животными.[39]
Хотя экспериментально не было показано, могут ли споры выжить при прохождении через пищеварительный тракт животного, регулярное присутствие Cyathus на коровий или конский навоз настоятельно предполагают, что это правда.[40] В качестве альтернативы, жесткая внешняя оболочка перидиол, выброшенная из брызговиков, может просто распадаться со временем, в конечном итоге высвобождая споры внутри.[41]
Биоактивные соединения

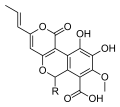
Ряд видов Cyathus производить метаболиты с биологической активностью и новыми химическими структурами, характерными для этого рода. Например, циатины находятся дитерпеноид соединения, производимые C. helenae,[44][45] C. africanus[46] и C. Earlei.[47] Некоторые циатины (особенно циатины B3 и C3), включая стриатин соединения из С. striatus,[48] показать сильный антибиотик Мероприятия.[44][49] Cyathane дитерпеноиды также стимулируют фактор роста нервов синтез и потенциально могут быть превращены в терапевтические агенты для нейродегенеративные расстройства Такие как Болезнь Альцгеймера.[50] Названные соединения циатускавины, выделенный из мицелия жидкая культура из С. stercoreus, имеют значительные антиоксидант Мероприятия,[43] как и соединения, известные как циатусалы, также из С. stercoreus.[51] Разные сесквитерпен соединения также были идентифицированы в C. Bulleri, включая кибродол (происходит от гумулен ),[52] нидулол, и буллерон.[53]
Разные Cyathus виды обладают противогрибковой активностью против патогенов человека, таких как Aspergillus fumigatus, грибковые микроорганизмы албиканс и Криптококк neoformans.[54] Экстракты С. striatus оказывают тормозящее действие на NF-κB, а фактор транскрипции ответственный за регулирование выражения нескольких генов, участвующих в иммунная система, воспаление, и смерть клетки.[55]
Человеческое использование
Съедобность
Виды семейства Nidulariaceae, включая Cyathus, считаются несъедобными, поскольку они «недостаточно крупные, мясистые или пахучие, чтобы представлять интерес для людей в качестве пищи».[56] Однако сообщений об отравлении не поступало. алкалоиды или другие вещества, которые считаются токсичными для человека. Броуди отмечает, что два Cyathus виды использовались коренными народами в качестве афродизиак, или стимулировать плодородие: C. limbatus в Колумбия, и C. microsporus в Гваделупа. Имеют ли эти виды какое-либо реальное влияние на физиология человека неизвестно.[57]
Биоразложение
Лигнин представляет собой сложное полимерное химическое соединение, входящее в состав древесины. Устойчивый к биологическому разложению, его присутствие в бумаге делает ее более слабой и склонной к обесцвечиванию под воздействием света. Виды C. Bulleri содержит три фермента, разрушающих лигнин: лигнин пероксидаза, пероксидаза марганца, и лакказа.[58] Эти ферменты имеют потенциальные приложения не только в целлюлозно-бумажная промышленность, но также для увеличения усвояемости и содержания белка корм для крупного рогатого скота. Потому что лаккасы могут сломаться фенольные соединения они могут использоваться для детоксикации некоторых загрязнителей окружающей среды, таких как красители используется в текстильная промышленность.[59][60][61] C. Bulleri лакказа также была генно-инженерный будет производиться кишечная палочка, что делает его первой грибковой лакказой, продуцируемой в бактериальном хозяине.[60] C. pallidus может биоразлагать взрывчатое вещество Гексоген (гексагидро-1,3,5-тринитро-1,3,5-триазин), предполагая, что он может быть использован для обеззараживания боеприпасы -загрязненные почвы.[62]
сельское хозяйство

Cyathus olla была исследована на предмет его способности ускорять разложение стерни, оставшейся на поле после уборки урожая, эффективно сокращая возбудитель населения и ускорение круговорот питательных веществ через минерализация основных питательных веществ для растений.[63][64]
Инфрагенерическая классификация
Род Cyathus был впервые разделен братьями Туласне на две внутриродовые группы (т. е. группирующие виды ниже ранга рода); группа "eucyathus" имела плодовые тела с внутренними поверхностями, сложенными в складки (пликации), в то время как группа «олла» отсутствовала.[5] Позже (1906), Ллойд опубликовал другую концепцию внутриродовой группировки в Cyathus, описывая пять групп, две в группе eucyathus и пять в группе olla.[7]
В 1970-х годах Броди в своей монографии о грибах птичьих гнезд выделил род Cyathus на семь связанных групп на основе ряда таксономический характеристики, включая наличие или отсутствие складок, структуру перидиол, цвет плодовых тел и характер волосков на внешнем перидии:[65]
Олла группа: Виды с tomentum наличие тонких приплюснутых волосков и отсутствие складок.
- C. olla, C. africanus, К. бадиус, C. canna, C. colensoi, C. confusus, C. Earlei, C. Hookeri, C. microsporus, C. minimus, C. pygmaeus
Паллидус группа: Виды с заметными длинными, направленными вниз волосками и гладким (не складчатым) внутренним перидием.
- C. pallidus, C. julietae
Триплекс группа: Виды с перидиями преимущественно темного цвета и серебристо-белой внутренней поверхностью.
- C. триплекс, C. setosus, C. sinensis
Грацилис группа: Виды с волосками войлока, собранными в пучки или холмики.
- C. gracilis, C. промежуточный, C. crassimurus, К. Эльмери
 Мохнатая (войлочная) внешняя перидиальная поверхность С. striatus
Мохнатая (войлочная) внешняя перидиальная поверхность С. striatus

Stercoreus группа: Виды с непластиковыми перидиями, мохнатыми или шерстистыми наружными перидиями и перидиолами от темных до черных.
- С. stercoreus, C. pictus, C. fimicola
Poeppigii группа: Виды с складчатыми внутренними перидиальными стенками, внешними стенками от волосистых до мохнатых, перидиолами от темных до черных и крупными, примерно сферическими или эллипсоидальными спорами.
- C. poeppigii, C. crispus, C. limbatus, C. gayanus, C. costatus, C. cheliensis, C. olivaceo-brunneus
Стриатус группа: Виды с складчатыми внутренними перидиями, от волосистых до мохнатых на внешних перидиях и в основном с эллиптическими спорами.
- С. striatus, C. annulatus, С. berkeleyanus, C. Bulleri, C. chevalieri, C. ellipsoideus, C. helenae, C. montagnei, C. nigro-albus, С. novae-zeelandiae, C. pullus, C. rudis
Филогенетический анализ
Публикация 2007 г. филогенетический анализ данных последовательностей ДНК многих Cyathus Виды поставили под сомнение обоснованность основанных на морфологии внутриродовых классификаций, описанных Броди. Это исследование предполагает, что Cyathus виды можно сгруппировать в три генетически связанных клады:[66]

ollum группа: C. africanus (тип ), C. africanus f. латиспор, C. conlensoi, С. griseocarpus, C. guandishanensis, C. Hookeri, С. jiayuguanensis, C. olla, C. olla f. anglicus, и C. olla f. Brodiensis.
полосатое тело группа: C. annulatus, C. crassimurus, C. helenae, C. poeppigii, C. renwei, C. setosus, С. stercoreus, и C. триплекс.
паллидум группа: С. berkeleyanus, C. olla f. ланатус, С. gansuensis, и C. pallidus.
Этот анализ показывает, что, а не структура плодового тела, спора размер, как правило, является более надежным признаком для разделения групп видов в Cyathus.[66] Например, виды в ollum все клады имеют длину спор менее 15 мкм, в то время как все представители паллидум группы имеют длину более 15 мкм; в полосатое тело группу, однако, нельзя отличить от паллидум группировать только по размеру спор. Две характеристики наиболее подходят для различения членов ollum группа из паллидум группа: толщина волосяного покрова на поверхности перидия и очертание плодовых тел. Ткань Паллидум Вид толстый, как войлок, и обычно собирается в пучки лохматых или пушистых волос. Их плодовые тела тигельной формы не имеют четко дифференцированной ножки. Экзоперидиум Ollum вид, для сравнения, имеет тонкий войлок из тонких волосков; плодовые тела имеют воронкообразную форму и имеют либо суженное основание, либо отчетливое ножка.[66]
Смотрите также
Рекомендации
- ^ Кирк MP, Cannon PF, Minter DW, Stalpers JA. (2008). Словарь грибов (10-е изд.). Уоллингфорд, Великобритания: CAB International. п. 185. ISBN 978-0-85199-826-8.CS1 maint: использует параметр авторов (связь)
- ^ а б Броди (1975), стр. 15.
- ^ а б c d Броди (1975), стр. 150.
- ^ Шмитц Дж. (1842). "Morphologische Beobachtungen als Beiträge zur Leben und Entwicklungsgeschichte einiger Schwämme aus der Klasse der Gastromyceten und Hymenomyceten". Linnea (на немецком). 16: 141–215.
- ^ а б Туласне Л. Р., Туласне К. (1844). "Исследования по организации и способу плодоношения шампиньонов из племени нидулевых, suivies d'un essai monographique". Анналы естественных наук, 3-я серия (На французском). 1: 41–107.CS1 maint: использует параметр авторов (связь)
- ^ а б Белый VS. (1902). «Nidulariaceae Северной Америки». Бюллетень Ботанического клуба Торри. 29 (5): 251–80. Дои:10.2307/2478721. JSTOR 2478721.
- ^ а б c Ллойд К.Г. (1906). «Нидулярные». Микологические сочинения. 2: 1–30.
- ^ Каннингем Г.Х. (1924). "Пересмотренный вариант новозеландских Nidulariales или грибов" птичьих гнезд ".'". Сделки Новозеландского института. 55: 59–66.
- ^ Броди, Грибы птичьего гнезда.
- ^ а б c Миллер HR, Миллер ОК. (1988). Гастеромицеты: морфологические особенности и особенности развития с ключами к порядкам, семействам и родам. Эврика, Калифорния: Mad River Press. п. 71. ISBN 978-0-916422-74-5.CS1 maint: использует параметр авторов (связь)
- ^ Броди (1975), стр. 5–7.
- ^ Броди (1975), стр. 147.
- ^ Алексополус и другие. (1996), стр. 545.
- ^ Броди (1975), стр. 6.
- ^ Броди (1975), стр. 129.
- ^ Brodie (1975), стр. 102–3.
- ^ Броди (1975), стр. 105.
- ^ а б Броди (1975), стр. 101.
- ^ Brodie HJ. (1958). «Возобновление роста и появление двойных плодовых тел у Nidulariaceae». Свенск Ботаниск Тидскрифт. 52: 373–8.
- ^ Brodie HJ. (1966). "Новый вид Cyathus из канадских Скалистых гор ». Канадский журнал ботаники. 44 (10): 1235–7. Дои:10.1139 / b66-138.
- ^ Броди (1975), стр. 116.
- ^ а б Броди (1975), стр. 117.
- ^ Гёшева ММ, Денчев СМ, Димитрова Э.Г., Ассев Б, Петрова РД, Стойчев ГТ. (2006). «Красный список грибов Болгарии» (PDF). Mycologia Balcanica. 3: 81–7.CS1 maint: использует параметр авторов (связь)
- ^ "DMU - B-FDC - Den danske Rødliste". Национальный институт экологических исследований. 1997. Архивировано с оригинал на 2011-07-18. Получено 2009-03-26.
- ^ Перич Б., Перич О. (2005). «Предварительный красный список исчезающих макромицетов Черногории» (PDF). Европейский совет по сохранению грибов. Получено 2009-03-26.CS1 maint: использует параметр авторов (связь)
- ^ Эванс С. (2007). «Красный список британских грибов, находящихся под угрозой исчезновения» (PDF). Британское микологическое общество. Получено 2009-03-26.
- ^ Пойнар Дж. Младший (2014). «Грибки птичьих гнезд (Nidulariales: Nidulariaceae) в балтийском и доминиканском янтаре». Грибковая биология. 118 (3): 325–29. Дои:10.1016 / j.funbio.2014.01.004. PMID 24607356.
- ^ Шмидт О. (2006). Древесина и древесные грибы: биология, повреждение, защита и использование. Берлин, Германия: Springer. С. 10–1. ISBN 978-3-540-32138-5.
- ^ Дьякон (2005), стр. 231–4.
- ^ Дьякон (2005) стр. 31–2.
- ^ Brodie HJ. (1948). "Изменения плодовых тел Cyathus stercoreus произведено в культуре ». Микология. 40 (5): 614–26. Дои:10.2307/3755260. JSTOR 3755260.
- ^ Гарнетт Э. (1958). Исследования факторов, влияющих на формирование плодовых тел у Cyathus stercoreus (Шв.) Де Тони (Кандидатская диссертация). Блумингтон, Индиана: Университет Индианы.
- ^ Лу Б. (1965). «Роль света в плодоношении базидиомицетов. Cyathus stercoreus". Американский журнал ботаники. 52 (5): 432–7. Дои:10.2307/2440258. JSTOR 2440258.
- ^ Броди (1975), стр. 57–8.
- ^ Броди (1975), стр. 48.
- ^ Броди (1975), стр. 89.
- ^ Мартин GW. (1927). «Базидии и споры Nidulariaceae». Микология. 19 (5): 239–47. Дои:10.2307/3753710. JSTOR 3753710.
- ^ Buller AHR. (1942). «Водяные брызги грибов птичьих гнезд, печеночников и мхов». Труды Королевского общества Канады. III. 36: 159.
- ^ Brodie HJ. (1951). «Механизм разбрызгивания в растениях». Канадский журнал ботаники. 29 (3): 224–34. Дои:10.1139 / b51-022.
- ^ Алексополус и другие. (1996), стр. 555.
- ^ Орр ДБ, Орр РТ. (1979). Грибы Западной Северной Америки. Беркли, Калифорния: Калифорнийский университет Press. п. 117. ISBN 978-0-520-03656-7.CS1 maint: использует параметр авторов (связь)
- ^ Сложный C09079 в КЕГГ База данных Pathway.
- ^ а б Кан Х.С., Ким К.Р., Джун Э.М., Пак Ш., Ли Т.С., Сух Дж. У., Ким Дж. П. (2008). "Циатускавины A, B и C, новые поглотители свободных радикалов с активностью защиты ДНК из базидиомицетов". Cyathus stercoreus". Письма по биоорганической и медицинской химии. 18 (14): 4047–50. Дои:10.1016 / j.bmcl.2008.05.110. PMID 18565749.CS1 maint: использует параметр авторов (связь)
- ^ а б Allbutt AD, Ayer WA, Brodie HJ, Johri BN, Taube H. (1971). "Циатин, новый комплекс антибиотиков, производимый Cyathus helenae". Канадский журнал микробиологии. 17 (11): 1401–7. Дои:10,1139 / м71-223. PMID 5156938.CS1 maint: использует параметр авторов (связь)
- ^ Айер В.А., Таубе Х. (1972). "Метаболиты Cyathus helenae". Буквы Тетраэдра. 13 (19): 1917–20. Дои:10.1016 / S0040-4039 (01) 84751-1.CS1 maint: использует параметр авторов (связь)
- ^ Айер В.А., Йошида Т., Ван Ши Д.М. (1978). «Дитерпеноидные метаболиты Cyathus africanus Броди ". Канадский химический журнал. 56 (16): 2197–9. Дои:10.1139 / v78-345.CS1 maint: использует параметр авторов (связь)
- ^ Айер WA, Ли SP. (1979). «Метаболиты грибов птичьих гнезд. Часть 11. Дитерпеноидные метаболиты Cyathus Earlei Ллойд ". Канадский химический журнал. 57 (24): 3332–7. Дои:10.1139 / v79-543.CS1 maint: использует параметр авторов (связь)
- ^ Анке Т., Обервинклер Ф. (1977). "Стриатины - новые антибиотики из базидиомицетов. Cyathus striatus (Huds. Ex Pers.) Willd ". Журнал антибиотиков. 30 (3): 221–5. Дои:10.7164 / антибиотики.30.221. PMID 863783.CS1 maint: использует параметр авторов (связь)
- ^ Johri BN, Brodie HJ, Allbutt AD, Ayer WA, Taube H. (1971). "Ранее неизвестный комплекс антибиотиков из грибка Cyathus helenae". Experientia. 27 (7): 853. Дои:10.1007 / BF02136907. PMID 5167804.CS1 maint: использует параметр авторов (связь)
- ^ Кшичковский В. (2008). «Строение, лечебные свойства и биосинтез циатановых дитерпеноидов». Биотехнология. 1 (80): 146–67.
- ^ Кан Х.С., Джун Э.М., Пак Ш., Хео СДЖ, Ли Т.С., Ю ID, Ким Дж.П. (2007). "Cyathusals A, B и C, антиоксиданты из ферментированных грибов. Cyathus stercoreus". Журнал натуральных продуктов. 70 (6): 1043–5. Дои:10.1021 / np060637h. PMID 17511503.CS1 maint: использует параметр авторов (связь)
- ^ Айер WA, Маккаскилл RH. (1981). «Кибродины, новый класс сесквитерпенов». Канадский химический журнал. 59 (14): 2150–8. Дои:10.1139 / v81-310.CS1 maint: использует параметр авторов (связь)
- ^ Айер WA, Маккаскилл RH. (1987). "Буллерон, новый сесквитерпеноид из Cyathus Bulleri Броди ". Канадский химический журнал. 65 (1): 15–7. Дои:10.1139 / v87-003.CS1 maint: использует параметр авторов (связь)
- ^ Лю Ю.Дж., Чжан К.К. (2004). «Антимикробная активность избранных Cyathus разновидность". Микопатология. 157 (2): 185–9. Дои:10.1023 / B: MYCO.0000020598.91469.d1. PMID 15119855. S2CID 26899906.CS1 maint: использует параметр авторов (связь)
- ^ Петрова Р.Д., Махаджна Дж., Резник А.З., Вассер С.П., Денчев С.М., Нево Э. (2007). «Грибковые вещества как модуляторы пути активации NF-κB». Отчеты по молекулярной биологии. 34 (3): 145–54. Дои:10.1007 / s11033-006-9027-5. PMID 17094008. S2CID 21776146.CS1 maint: использует параметр авторов (связь)
- ^ Броди (1975), стр. 119.
- ^ Броди (1975), стр. 120.
- ^ Васдев К. (1995). Ферменты, разрушающие лигнин, и лигнинолитическая система Cyathus зр (Кандидатская диссертация). Дели, Индия: Университет Дели.
- ^ Чабра М., Мишра С., Срикришнан Т.Р. (2008). «Обесцвечивание и детоксикация текстильных красок / смеси красителей с помощью медиатора путем Cyathus Bulleri лакказа ". Прикладная биохимия и биотехнология. 151 (2–3): 587–98. Дои:10.1007 / s12010-008-8234-z. PMID 18506632. S2CID 38874993.CS1 maint: использует параметр авторов (связь)
- ^ а б Салони, Гарг Н., Баранвал Р., Чхабра М., Мишра С., Чаудхури Т.К., Бисария В.С. (2008). "Лакказа Cyathus bulleri: структурная, каталитическая характеристика и выражение в кишечная палочка". Biochimica et Biophysica Acta (BBA) - Белки и протеомика. 1784 (2): 259–68. Дои:10.1016 / j.bbapap.2007.11.006. PMID 18083129.CS1 maint: использует параметр авторов (связь)
- ^ Salony, Mishra S, Bisaria VS. (2006). «Производство и характеристика лакказы из Cyathus Bulleri и его использование для обесцвечивания стойких красителей для текстиля ». Прикладная микробиология и биотехнология. 71 (5): 646–53. Дои:10.1007 / s00253-005-0206-4. PMID 16261367. S2CID 38760041.CS1 maint: использует параметр авторов (связь)
- ^ Байман П., Ричи С.Д., Беннетт Дж. В.. (1995). «Грибковые взаимодействия с взрывчатым гексогеном (гексагидро-1,3,5-тринитро-1,3,5-триазин)». Журнал промышленной микробиологии. 15 (5): 418–23. Дои:10.1007 / BF01569968. S2CID 20115808.CS1 maint: использует параметр авторов (связь)
- ^ Бленис П.В., Чоу П. (2005). «Оценка древесных грибов и канолы на их способность разлагать стерню канолы». Канадский журнал патологии растений. 27 (2): 259–67. Дои:10.1080/07060660509507223. S2CID 83908810.CS1 maint: использует параметр авторов (связь)
- ^ Shinners-Carnelley TC, Szpacenko A, Tewari JP, Palcic MM. (2002). «Ферментативная активность Cyathus olla во время твердофазной ферментации корней канолы » (PDF). Фитопротекция. 83 (1): 31–40. Дои:10.7202 / 706227ar. Архивировано из оригинал (PDF) на 2014-12-07.CS1 maint: использует параметр авторов (связь)
- ^ Броди (1975), стр. 150–80.
- ^ а б c Чжао Р.Л., Дживон Р., Дежардин Д.Е., Сойтонг К., Хайд К.Д. (2007). "Филогении рибосомной ДНК Cyathus: уместна ли нынешняя внутриродовая классификация? ». Микология. 99 (3): 385–95. Дои:10.3852 / mycologia.99.3.385. PMID 17883030.CS1 maint: использует параметр авторов (связь)
Цитированные тексты
- Алексопулос CJ, Мимс CW, Блэквелл М. (1996). Вводная микология (4-е изд.). Нью-Йорк, Нью-Йорк: Wiley. ISBN 0-471-52229-5.CS1 maint: использует параметр авторов (связь)
- Brodie HJ. (1975). Грибы птичьего гнезда. Торонто: Университет Торонто Press. ISBN 0-8020-5307-6.
- Дьякон Дж. (2005). Грибковая биология. Кембридж, Массачусетс: Blackwell Publishers. ISBN 1-4051-3066-0.