Гемозоин - Hemozoin

Гемозоин продукт утилизации, образовавшийся в результате переваривания кровь кровью паразиты. Эти гематофаг организмы, такие как малярия паразиты (Плазмодий виды), Родниус и Шистосома переваривать гемоглобин и выпустить большое количество бесплатных гем, который является небелковым компонентом гемоглобин. Гем - это протезная группа состоящий из утюг атом, содержащийся в центре гетероциклический порфирин звенеть. Свободный гем токсичен для клеток, поэтому паразиты превращают его в нерастворимую кристаллическую форму, называемую гемозоином. У малярийных паразитов гемозоин часто называют пигмент малярии.
Поскольку образование гемозоина необходимо для выживания этих паразитов, он является привлекательной мишенью для разработка лекарств и много изучается в Плазмодий как способ найти лекарства для лечения малярия (малярия Ахиллесова пята ). Несколько в настоящее время используются противомалярийные препараты, Такие как хлорохин и мефлохин, как полагают, убивают паразитов малярии, ингибируя гемозоин биокристаллизация.
Открытие
Черно-коричневый пигмент наблюдали Иоганн Генрих Меккель [1] в 1847 г. - в крови и селезенке человека, страдающего безумием.[2] Однако только в 1849 году присутствие этого пигмента было связано с заражением малярией.[3] Первоначально считалось, что этот пигмент вырабатывается организмом в ответ на инфекцию, но Шарль Луи Альфонс Лаверан в 1880 г. осознал, что «пигмент малярии» вместо этого вырабатывается паразитами, поскольку они размножаются в эритроцит.[4] Связь между пигментом и малярийными паразитами использовалась Рональд Росс определить этапы Плазмодий жизненный цикл, который происходит внутри комара, поскольку, хотя эти формы паразита по внешнему виду отличаются от стадий крови, они все же содержат следы пигмента.
Позже, в 1891 году Т. Карбоне и У. Браун (1911) опубликовал статьи, связывающие деградацию гемоглобина с производством пигмента, описывая пигмент малярии как форму гематина и опровергая широко распространенное представление о том, что он связан с меланин. Браун заметил, что все меланины быстро обесцвечиваются перманганатом калия, в то время как с этим реагентом малярийный пигмент не проявляет ни малейших признаков истинной реакции отбеливания.[5][6] Название «гемозоин» было предложено Луи Вестенра Самбон.[7] В 1930-х годах несколько авторов идентифицировали гемозоин как чистую кристаллическую форму α-гематина и показали, что это вещество не содержит белков внутри кристаллов,[4] но никакого объяснения различий в растворимости малярийного пигмента и кристаллов α-гематина дано не было.
Формирование
Во время цикла бесполого интраэритроцитарного воспроизводства Плазмодий falciparum потребляет до 80% хозяйской клетки гемоглобин.[8][9] При переваривании гемоглобина высвобождается мономерный α-гематин (феррипротопорфирин IX). Это соединение токсично, так как это прооксидант и катализирует производство активные формы кислорода. Окислительный стресс Полагают, что он образуется при превращении гема (ферропротопорфирин) в гематин (феррипротопорфирин). Свободный гематин также может связываться и нарушать клеточные мембраны, повреждая клеточные структуры и вызывая лизис эритроцитов хозяина.[10] Уникальная реакционная способность этой молекулы была продемонстрирована в нескольких in vitro и in vivo условия эксперимента.[11]

Таким образом, малярийный паразит выводит токсины из гематина за счет биокристаллизация - превращение его в нерастворимый и химически инертный β-гематин кристаллы (называется гемозоин).[13][14][15] В Плазмодий пищевая вакуоль заполняется кристаллами гемозоина, которых около 100-200 нанометры длинные и каждая содержит около 80 000 молекул гема.[4] Детоксикация посредством биокристаллизации отличается от процесса детоксикации у млекопитающих, когда фермент, называемый гемоксигеназа вместо этого разбивает лишний гем на биливердин, утюг, и монооксид углерода.[16]
Было предложено несколько механизмов производства гемозоина в Плазмодий, и область очень спорная, с мембраной липиды,[17][18] богатые гистидином белки,[19] или даже их комбинация,[20] предлагается катализировать образование гемозоина. Другие авторы описали белок детоксикации гема, который, как утверждается, более эффективен, чем липиды или богатые гистидином белки.[12] Возможно, что многие процессы способствуют образованию гемозоина.[21]Образование гемозоина у других организмов, питающихся кровью, изучено не так хорошо, как у Плазмодий.[22] Однако исследования по Schistosoma mansoni показали, что это паразитический червь производит большое количество гемозоина во время его роста в кровотоке человека. Хотя форма кристаллов отличается от кристаллов, производимых малярийными паразитами,[23] Химический анализ пигмента показал, что он состоит из гемозоина.[24][25] Подобным же образом кристаллы образовались в кишечнике целующего жука. Родниус проликсус При переваривании крови муки также имеют уникальную форму, но состоят из гемозоина.[26] Формирование Гц в Р. проликсус средняя кишка происходит в физиологически значимых физико-химических условиях, и липиды играют важную роль в биокристаллизации гема. Выявлено, что автокаталитическая кристаллизация гема до Hz является неэффективным процессом, и это преобразование дополнительно снижается по мере увеличения концентрации Hz.
[27] Несколько других механизмов были разработаны для защиты большого разнообразия гематофаг организмы против токсического действия свободного гема. Комаров переваривают пищу с кровью внеклеточно и не производят гемозоин. Гем сохраняется в перитрофическая матрица, слой белка и полисахаридов, который покрывает среднюю кишку и отделяет клетки кишечника от комка крови.[28]
Хотя β-гематин может производиться в анализы спонтанно на низком уровне pH, разработка простого и надежного метода измерения продукции гемозоина была трудной. Отчасти это связано с сохраняющейся неопределенностью в отношении того, какие молекулы участвуют в производстве гемозоина, а отчасти с трудностью измерения разницы между агрегированным или осажденным гемом и подлинным гемозоином.[29] Современные анализы чувствительны и точны, но требуют нескольких этапов промывки, поэтому они медленные и не идеальны для высокопроизводительный скрининг.[29] Однако с этими анализами были проведены некоторые скрининги.[30]
Структура


Кристаллы β-гематина состоят из димеры молекул гематина, которые, в свою очередь, соединяются водородными связями с образованием более крупных структур. В этих димерах утюг -кислород координационная связь связывает центральное железо одного гематина с кислородом карбоксилат боковая цепь соседнего гематина. Эти взаимные связи железо-кислород очень необычны и не наблюдались ни в одном другом димере порфирина. β-гематин может быть циклическим димер или линейный полимер,[31] полимерная форма никогда не была обнаружена в гемозоине, что опровергает широко распространенное представление о том, что гемозоин производится фермент гем-полимераза.[32]
Кристаллы гемозоина имеют ярко выраженный триклинический структура и слабо магнитный. Разница между диамагнитный низкоспиновый оксигемоглобин и парамагнитный гемозоин можно использовать для выделения.[33][34] Они также показывают оптические дихроизм, что означает, что они сильнее поглощают свет по длине, чем по ширине, что позволяет автоматизировать обнаружение малярии.[35] Гемозоин производится в форме, которая под действием применяемого магнитное поле, вызывает индуцированную оптическую дихроизм характеристика концентрации гемозоина; и точное измерение этого индуцированного дихроизм (Магнитный круговой дихроизм ) может использоваться для определения уровня малярийной инфекции.[36]
Ингибиторы
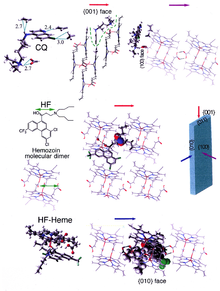
Образование гемозоина - отличная лекарственная мишень, поскольку он необходим для выживания малярийных паразитов и отсутствует у человека-хозяина. Гематин-мишень для лекарственного средства является производным от хозяина и в значительной степени находится вне генетического контроля паразита, что затрудняет развитие лекарственной устойчивости. Считается, что многие клинически применяемые препараты действуют путем ингибирования образования гемозоина в пищевой вакуоли.[37] Это предотвращает детоксикацию гема, высвобождаемого в этом отсеке, и убивает паразита.[38]
Наиболее понятные примеры такого гематина биокристаллизация ингибиторы хинолин наркотики, такие как хлорохин и мефлохин. Эти препараты связываются как со свободным гемом, так и с кристаллами гемозоина,[39] и, следовательно, блокировать добавление новых единиц гема в растущие кристаллы. Маленькое, наиболее быстро растущее лицо - это лицо, с которым, как полагают, связываются ингибиторы.[40][41]
Роль в патофизиологии
Гемозоин попадает в кровоток во время повторного заражения и фагоцитируется in vivo и in vitro фагоцитами хозяина и изменяет важные функции этих клеток. Большинство функциональных изменений были долгосрочными постфагоцитарными эффектами,[42][43] в том числе ингибирование эритропоэза показано in vitro.[44][45][46]Напротив, было показано, что мощная кратковременная стимуляция окислительного взрыва моноцитами человека также происходит во время фагоцитоза nHZ.[47]
Смотрите также
Рекомендации
- ^ Джанджуа Р.М., Шултка Р., Геббель Л., Пайт Т.Г., Шилдс CB (2010). «Наследие Иоганна Фридриха Меккеля Старшего (1724–1774): династия анатомов в 4 поколениях». Нейрохирургия. 66 (4): 758–770. Дои:10.1227 / 01.NEU.0000367997.45720.A6. PMID 20305497.
- ^ Меккель Х (1847). "Ueber schwarzes Pigment in der Milz und dem Blute einer Geisteskranken". Zeitschrift für Psychiatrie. IV: 198–226.
- ^ Вирхов Р. (1849). "Zur patologischen Physiologie des Bluts". Arch Pathol Anatomie Physiol Klin Med. 2 (3): 587–598. Дои:10.1007 / BF02114475.
- ^ а б c Салливан DJ (декабрь 2002 г.). «Теории образования малярийного пигмента и действия хинолина». Int J Parasitol. 32 (13): 1645–53. Дои:10.1016 / S0020-7519 (02) 00193-5. PMID 12435449.
- ^ Карбон Т (1891). "Sulla natura chimica del pigmento malarico". G R Accad Med Torino. 39: 901–906.
- ^ Браун WH (1911). «Малярийный пигмент (так называемый меланин): ЕГО природа и способ производства». J Exp Med. 13 (2): 290–299. Дои:10.1084 / jem.13.2.290. ЧВК 2124860. PMID 19867409.
- ^ Синтон Дж. А., Гош Б. Н. (1934). «Исследования малярийного пигмента (гемозоина). Часть I. Исследование действия растворителей на гемозоин и спектроскопические проявления, наблюдаемые в растворах». Записи обследования малярии в Индии. 4: 15–42.
- ^ Розенталь П.Дж., Мешник С.Р. (декабрь 1996 г.). «Катаболизм гемоглобина и утилизация железа малярийными паразитами». Мол Биохим Паразитол. 83 (2): 131–139. Дои:10.1016 / S0166-6851 (96) 02763-6. PMID 9027746.
- ^ Эспозито А., Тифферт Т., Мауриц Дж. М., Шлахтер С., Баннистер Л. Х., Камински К. Ф., Лью В. Л. (2008). Шнур JM (ред.). «Визуализация FRET концентрации гемоглобина в красных клетках, инфицированных Plasmodium falciparum». PLoS ONE. 3 (11): e3780. Дои:10.1371 / journal.pone.0003780. ЧВК 2582953. PMID 19023444.
- ^ Fitch CD, Чевли Р., Канджананггулпан П., Датта П., Чевли К., Чоу А.С. (1983). «Внутриклеточный феррипротопорфирин IX - литический агент». Кровь. 62 (6): 1165–1168. Дои:10.1182 / blood.V62.6.1165.1165. PMID 6640106.[постоянная мертвая ссылка ]
- ^ Hebbel RP, Eaton JW (апрель 1989 г.). «Патобиология взаимодействия гема с мембраной эритроцита». Семин Гематол. 26 (2): 136–149. PMID 2658089.
- ^ а б Яни Д., Нагаркатти Р., Битти В., Ангел Р., Слебодник С., Андерсен Дж., Кумар С., Ратор Д. (апрель 2008 г.). Ким К. (ред.). «HDP - новый белок гема для детоксикации от малярийных паразитов». PLoS Pathog. 4 (4): e1000053. Дои:10.1371 / journal.ppat.1000053. ЧВК 2291572. PMID 18437218.
- ^ Fitch CD, Kanjananggulpan P (ноябрь 1987 г.). «Состояние феррипротопорфирина IX в малярийном пигменте». J Biol Chem. 262 (32): 15552–1555. PMID 3119578.
- ^ Пагола С., Стивенс П. В., Боле Д. С., Косар А. Д., Мадсен С. К. (март 2000 г.). «Структура малярийного пигмента бета-гематина». Природа. 404 (6775): 307–310. Дои:10.1038/35005132. PMID 10749217.
- ^ Хемпельманн Э (2007). «Биокристаллизация гемозоина в Plasmodium falciparum и противомалярийная активность ингибиторов кристаллизации». Исследование паразитов. 100 (4): 671–676. Дои:10.1007 / s00436-006-0313-х. PMID 17111179. Архивировано из оригинал на 2011-06-10.
- ^ Кикучи Г., Ёсида Т., Ногучи М. (декабрь 2005 г.). «Гемоксигеназа и деградация гема». Biochem Biophys Res Commun. 338 (1): 558–567. Дои:10.1016 / j.bbrc.2005.08.020. PMID 16115609.
- ^ Пишотта Дж. М., Салливан Д. (июнь 2008 г.). «Гемозоин: масло против воды». Паразитол Инт. 57 (2): 89–96. Дои:10.1016 / j.parint.2007.09.009. ЧВК 2442017. PMID 18373972.
- ^ Хай НТ, Шима Й, Маэда А, Мэн ТТ, Хираяма К., Хирасе А, Миядзава А, Камей А (2013). «Фосфолипидное мембранно-опосредованное образование гемозоина: влияние физических свойств и доказательства гемозоина, окружающего мембрану». PLoS ONE. 8 (7): e70025. Дои:10.1371 / journal.pone.0070025. ЧВК 3720957. PMID 23894579.
- ^ Салливан Д.Д., Глузман И.Ю., Голдберг Д.Е. (январь 1996 г.). «Образование гемозоина плазмодия опосредовано белками, богатыми гистидином». Наука. 271 (5246): 219–222. Дои:10.1126 / science.271.5246.219. PMID 8539625.
- ^ Панди А.В., Баббарвал В.К., Окойе Д.Н., Джоши Р.М., Пури С.К., Сингх Р.Л., Чаухан В.С. (сентябрь 2003 г.). «Образование гемозоина при малярии: двухэтапный процесс с участием белков и липидов, богатых гистидином». Biochem Biophys Res Commun. 308 (4): 736–743. Дои:10.1016 / S0006-291X (03) 01465-7. PMID 12927780.
- ^ Чу М., Сундарараман В., Кумар С., Редди В. С., Сиддики В. А., Стюарт К. Д., Малхотра П. (апрель 2013 г.). «Белковый комплекс направляет образование гемоглобина в гемозоин у Plasmodium falciparum». Proc Natl Acad Sci U S A. 110 (14): 5392–7. Дои:10.1073 / pnas.1218412110. ЧВК 3619337. PMID 23471987.
- ^ Иган Т.Дж. (февраль 2008 г.). «Гемозоинформация». Мол Биохим Паразитол. 157 (2): 127–136. Дои:10.1016 / j.molbiopara.2007.11.005. PMID 18083247.
- ^ Мур GA, Homewood CA, Gilles HM (сентябрь 1975 г.). «Сравнение пигмента Schistosoma mansoni и Plasmodium berghei». Энн Троп Мед Паразитол. 69 (3): 373–374. Дои:10.1080/00034983.1975.11687021. PMID 1098591.
- ^ Оливейра М.Ф., д'Авила Дж.С., Торрес С.Р., Оливейра П.Л., Темпоне А.Дж., Румьянек Ф.Д., Брага С.М., Сильва-младший, Данса-Петрецки М., Оливейра М.А., де Соуза В., Феррейра СТ (ноябрь 2000 г.). «Гемозоин у Schistosoma mansoni». Мол Биохим Паразитол. 111 (1): 217–221. Дои:10.1016 / S0166-6851 (00) 00299-1. PMID 11087932.
- ^ Корреа Соарес Дж. Б., Менезеш Д., Ванье-Сантос М. А., Феррейра-Перейра А., Алмейда Г. Т., Венансио TM, Верджовски-Алмейда С., Зишири В. К., Кутер Д., Хантер Р., Иган Т. Дж., Оливейра М. Ф. (2009). Джонс МК (ред.). «Вмешательство в образование гемозоина представляет собой важный механизм шистосомицидного действия антималярийных хинолиновых метанолов». PLoS Negl Trop Dis. 3 (7): e477. Дои:10.1371 / journal.pntd.0000477. ЧВК 2703804. PMID 19597543.
- ^ Oliveira MF, Kycia SW, Gomez A, Kosar AJ, Bohle DS, Hempelmann E, Menezes D, Vannier-Santos MA, Oliveira PL, Ferreira ST (2005). «Структурная и морфологическая характеристика гемозоина, продуцируемого Schistosoma mansoni и Rhodnius prolixus». FEBS Lett. 579 (27): 6010–6016. Дои:10.1016 / j.febslet.2005.09.035. PMID 16229843.
- ^ Стиблер Р., Тимм Б.Л., Оливейра П.Л., Херн Г.Р., Иган Т.Дж., Оливейра М.Ф. (2010). «О физико-химических и физиологических требованиях образования гемозоина, которым способствуют перимикровиллярные мембраны в средней кишке Rhodnius prolixus». Насекомое Biochem Mol Biol. 40 (3): 284–292. Дои:10.1016 / j.ibmb.2009.12.013. PMID 20060043.
- ^ Паскоа В., Оливейра П.Л., Данса-Петрецки М., Сильва-младший, Альваренга П.Х., Якобс-Лорена М., Лемос Ф.Дж. (май 2002 г.). «Перитрофический матрикс Aedes aegypti и его взаимодействие с гемом при переваривании крови». Насекомое Biochem Mol Biol. 32 (5): 517–523. Дои:10.1016 / S0965-1748 (01) 00130-8. PMID 11891128.
- ^ а б Дорн А., Виппагунта С. Р., Матил Х, Бубендорф А., Веннерстрем Дж. Л., Ридли Р. Г. (март 1998 г.). «Сравнение и анализ нескольких способов стимулирования полимеризации гематина (гема) и оценка ее инициации in vitro». Биохим Фармакол. 55 (6): 737–747. Дои:10.1016 / S0006-2952 (97) 00509-1. PMID 9586945.
- ^ Теквани Б.Л., Уокер Л.А. (февраль 2005 г.). «Ориентация на путь синтеза гемозоина для открытия новых противомалярийных препаратов: технологии для анализа образования бета-гематина in vitro». Экран с высокой пропускной способностью Comb Chem. 8 (1): 63–79. Дои:10.2174/1386207053328101. PMID 15720198.
- ^ Лемберг Р., Легге Дж. В. (1949). «Гематиновые соединения и желчные пигменты». Interscience, Нью-Йорк.
- ^ Хемпельманн Э., Маркес Х.М. (сентябрь 1994 г.). «Анализ малярийного пигмента Plasmodium falciparum». J Pharmacol Toxicol Методы. 32 (1): 25–30. Дои:10.1016/1056-8719(94)90013-2. PMID 7833503.
- ^ Пол Ф., Роат С., Мелвилл Д., Уорхерст, округ Колумбия, Осисанья Дж. О. (1981). «Отделение инфицированных малярией эритроцитов из цельной крови: использование метода селективного высокоградиентного магнитного разделения». Ланцет. 2 (8237): 70–71. Дои:10.1016 / S0140-6736 (81) 90414-1. PMID 6113443.
- ^ Ким CC, Уилсон Е.Б., Деризи Дж.Л. (2010). «Усовершенствованные методы магнитной очистки от малярийных паразитов и гемозоина» (PDF). Малар Дж. 9 (1): 17. Дои:10.1186/1475-2875-9-17. ЧВК 2817699. PMID 20074366.
- ^ Mendelow BV, Lyons C, Nhlangothi P, Tana M, Munster M, Wypkema E, Liebowitz L, Marshall L, Scott S, Coetzer TL (1999). «Автоматизированное обнаружение малярии деполяризацией лазерного света». Br J Haematol. 104 (3): 499–503. Дои:10.1046 / j.1365-2141.1999.01199.x. PMID 10086786.[мертвая ссылка ]
- ^ Newman DM, Heptinstall J, Matelon RJ, Savage L, Wears ML, Beddow J, Cox M, Schallig HD, Mens P (2008). «Магнитооптический путь к диагностике малярии in vivo: предварительные результаты и данные доклинических испытаний» (PDF). Biophys J. 95 (2): 994–1000. Дои:10.1529 / biophysj.107.128140. ЧВК 2440472. PMID 18390603. Архивировано из оригинал (PDF) на 2011-07-27. Получено 2009-12-19.
- ^ Зиглер Дж, Линк Р., Райт Д.В. (февраль 2001 г.). «Ингибиторы агрегации гема: противомалярийные препараты, направленные на существенный процесс биоминерализации». Curr Med Chem. 8 (2): 171–89. Дои:10.2174/0929867013373840. PMID 11172673.
- ^ Коронадо Л.М., Надович СТ, Спадафора С. (2014). «Малярийный гемозоин: от цели к инструменту». Biochim Biophys Acta. 1840 (6): 2032–2041. Дои:10.1016 / j.bbagen.2014.02.009. ЧВК 4049529. PMID 24556123.
- ^ Салливан Д. Д., Глузман И. Ю., Рассел Д. Г., Голдберг Д. Е. (октябрь 1996 г.). «О молекулярном механизме противомалярийного действия хлорохина». Proc Natl Acad Sci USA. 93 (21): 11865–70. Дои:10.1073 / пнас.93.21.11865. ЧВК 38150. PMID 8876229.
- ^ де Вильерс К.А., Маркес Х.М., Эган Т.Дж. (август 2008 г.). «Кристаллическая структура галофантрин-феррипротопорфирина IX и механизм действия противомалярийных препаратов арилметанола». J Inorg Biochem. 102 (8): 1660–1667. Дои:10.1016 / j.jinorgbio.2008.04.001. PMID 18508124.
- ^ Weissbuch I, Leiserowitz L (2008). «Взаимодействие между малярией, образованием кристаллического гемозоина и действием и дизайном противомалярийных препаратов». Chem Rev. 108 (11): 4899–4914. Дои:10.1021 / cr078274t. PMID 19006402.
- ^ Арезе П., Шварцер Э. (1997). «Малярийный пигмент (гемозоин): очень активное« инертное »вещество». Энн Троп Мед Паразитол. 91 (5): 501–516. Дои:10.1080/00034989760879. PMID 9329987.
- ^ Скороход О.А., Алессио М., Мордмюллер Б., Арезе П., Шварцер Э. (2004). «Гемозоин (малярийный пигмент) ингибирует дифференцировку и созревание дендритных клеток человека, происходящих из моноцитов: гамма-опосредованный эффект рецептора, активируемого пролифератором пероксисом». J Immunol. 173 (6): 4066–74. Дои:10.4049 / jimmunol.173.6.4066. PMID 15356156.
- ^ Джирибальди Г., Ульерс Д., Шварцер Э., Робертс И., Пьяцибелло В., Арезе П. (2004). «Гемозоин- и 4-гидроксиноненал-опосредованное ингибирование эритропоэза. Возможная роль в малярийном дизеритропоэзе и анемии». Haematologica. 89 (4): 492–493. PMID 15075084.
- ^ Casals-Pascual C, Kai O, Cheung JO, Williams S, Lowe B, Nyanoti M, Williams TN, Maitland K, Molyneux M, Newton CR, Peshu N, Watt SM, Roberts DJ (2006). «Подавление эритропоэза при малярийной анемии связано с гемозоином in vitro и in vivo». Кровь. 108 (8): 2569–77. Дои:10.1182 / кровь-2006-05-018697. PMID 16804108.
- ^ Скороход О.А., Кайоне Л., Маррокко Т., Мильярди Г., Баррера В., Арезе П., Пьяцибелло В., Шварцер Е. (2010). «Ингибирование эритропоэза при малярийной анемии: роль гемозоина и 4-гидроксиноненала, вырабатываемого гемозоином». Кровь. 116 (20): 4328–37. Дои:10.1182 / blood-2010-03-272781. PMID 20686121. Архивировано из оригинал 19 июня 2013 г.
- ^ Баррера В., Скороход О.А., Бачи Д., Гремо Г., Арезе П., Шварцер Э. (2011). «Фибриноген хозяина, стабильно связанный с гемозоином, быстро активирует моноциты через TLR-4 и CD11b / CD18-интегрин: новая парадигма действия гемозоина». Кровь. 117 (21): 5674–82. Дои:10.1182 / blood-2010-10-312413. PMID 21460246. Архивировано из оригинал на 2013-07-09.