Зеркальный нейрон - Mirror neuron
Эта статья должна быть обновлено. (Апрель 2020) |
| Система зеркал | |
|---|---|
| Идентификаторы | |
| MeSH | D059167 |
| Анатомические термины нейроанатомии | |
А зеркальный нейрон это нейрон который пожары как когда животное действует, так и когда животное наблюдает за тем же действием, выполняемым другим.[1][2][3] Таким образом, нейрон «отражает» поведение другого, как если бы наблюдатель действовал сам. Такие нейроны непосредственно наблюдались у человека.[нужна цитата ] и примат разновидность,[4] и птицы.[5]
У людей активность мозга, соответствующая активности зеркальных нейронов, была обнаружена в премоторная кора, то дополнительная моторная зона, то первичная соматосенсорная кора, а нижняя теменная кора.[6] Функция зеркальной системы у людей является предметом множества предположений. Было показано, что птицы обладают имитационным резонансным поведением, а неврологические данные свидетельствуют о наличии некоторой формы зеркальной системы.[4][7]
На сегодняшний день не было предложено общепринятых нейронных или вычислительных моделей, описывающих, как активность зеркальных нейронов поддерживает когнитивные функции.[8][9][10] Тема зеркальных нейронов продолжает вызывать бурные споры. В 2014, Философские труды Королевского общества B опубликовал специальный выпуск, целиком посвященный исследованиям зеркальных нейронов.[11]
Некоторые исследователи когнитивной нейробиологии и когнитивная психология считают, что эта система обеспечивает физиологический механизм для связи восприятия / действия (см. общая теория кодирования ).[3] Они утверждают, что зеркальные нейроны могут быть важны для понимания действий других людей и для обучения новым навыкам путем имитации. Некоторые исследователи предполагают, что зеркальные системы могут имитировать наблюдаемые действия и тем самым способствовать теория разума навыки,[12][13] в то время как другие связывают зеркальные нейроны с язык способности.[14] Нейробиологи, такие как Марко Якобони (UCLA), утверждали, что зеркальные нейронные системы в человеческий мозг помочь нам понять действия и намерения других людей. В исследовании, опубликованном в марте 2005 г., Якобони и его коллеги сообщили, что зеркальные нейроны могут различать, планировал ли другой человек, взяв чашку чая, пить из нее или убрать ее со стола.[15] Кроме того, Якобони утверждал, что зеркальные нейроны являются нейронной основой человеческой способности к эмоциям, таким как сочувствие.[16]
Есть ученые, которые скептически относятся к теориям, которые выдвигаются для объяснения функции зеркальных нейронов. В статье 2013 г. Проводной Кристиан Джарретт предупредил, что:
... зеркальные нейроны - захватывающее, интригующее открытие, но когда вы видите их упоминание в СМИ, помните, что большая часть исследований этих клеток проводилась на обезьянах. Помните также, что существует много разных типов зеркальных нейронов. И что мы все еще пытаемся точно установить, существуют ли они у людей и как они сравниваются с версиями обезьян. Что касается понимания функционального значения этих клеток… не дайте себя обмануть: это путешествие только начинается.[17]
Открытие
В 1980-х и 1990-х годах нейрофизиологи Джакомо Риццолатти, Джузеппе Ди Пеллегрино, Лучано Фадига, Леонардо Фогасси и Витторио Галлезе на Университет Пармы размещен электроды в вентральная премоторная кора из макака обезьяна учиться нейроны специализируется на контроле движений рук и рта; например, захват объекта и манипулирование им. Во время каждого эксперимента исследователи позволяли обезьяне тянуться к кусочкам пищи и регистрировали отдельные нейроны в мозгу обезьяны, таким образом измеряя реакцию нейрона на определенные движения.[18][19] Они обнаружили, что некоторые нейроны реагировали, когда обезьяна наблюдала, как человек берет кусок пищи, а также когда обезьяна сама берет пищу. Природа, но был отклонен из-за «отсутствия всеобщего интереса» перед публикацией в менее конкурентоспособном журнале.[20]
Несколько лет спустя та же группа опубликовала еще одну эмпирическую работу, в которой обсуждалась роль системы зеркальных нейронов в распознавании действий и предполагалось, что человеческий Регион Брока был гомолог область вентральной премоторной коры обезьяны.[21]Хотя в этих статьях сообщалось о наличии зеркальных нейронов, реагирующих на движения рук, последующее исследование Пьера Франческо Феррари и его коллег[22] описал присутствие зеркальных нейронов, реагирующих на движения рта и мимические жесты.
Дальнейшие эксперименты подтвердили, что около 10% нейронов в нижней лобной и нижней теменной коре головного мозга обезьяны обладают «зеркальными» свойствами и дают аналогичные ответы на выполняемые действия руками и наблюдаемые действия. В 2002 Кристиан Кейзерс и его коллеги сообщили, что как у людей, так и у обезьян зеркальная система также реагирует на звук действий.[3][23][24]
Отчеты о зеркальных нейронах широко публиковались.[25] и подтвердил[26] с зеркальными нейронами, обнаруженными как в нижних лобных, так и в нижних теменных областях мозга. Недавно свидетельства из функциональная нейровизуализация настоятельно предполагает, что у людей есть похожие системы зеркальных нейронов: исследователи определили области мозга, которые реагируют как во время действия, так и во время наблюдения за действием. Неудивительно, что эти области мозга включают те, что обнаружены у макак.[1] Тем не мение, функциональная магнитно-резонансная томография (фМРТ) может исследовать весь мозг сразу и предполагает, что гораздо более широкая сеть областей мозга проявляет зеркальные свойства у людей, чем считалось ранее. Эти дополнительные области включают соматосенсорная кора и считается, что они заставляют наблюдателя почувствовать, что он чувствует, двигаясь наблюдаемым путем.[27][28]
Источник
Многие неявно предполагают, что зеркальность зеркальных нейронов обусловлена в первую очередь наследственными генетическими факторами и что генетическая предрасположенность к развитию зеркальных нейронов эволюционировала, потому что они облегчают понимание действий.[29] Напротив, ряд теоретических отчетов утверждает, что зеркальные нейроны могут просто возникнуть из-за усвоенных ассоциаций, включая Hebbian Theory,[30] в Ассоциативное обучение Теория,[29] Канализация [31] и Exaptation.[нужна цитата ]
У обезьян

Первое животное, у которого исследователи индивидуально изучили зеркальные нейроны, - это макака обезьяна. У этих обезьян зеркальные нейроны находятся в нижняя лобная извилина (область F5) и нижняя теменная долька.[1]
Считается, что зеркальные нейроны опосредуют понимание других животных. поведение. Например, зеркальный нейрон, который срабатывает, когда обезьяна разрывает лист бумаги, также срабатывает, когда обезьяна видит, как человек рвет бумагу, или слышит разрыв бумаги (без визуальных подсказок). Эти свойства заставили исследователей поверить в то, что зеркальные нейроны кодируют абстрактные концепции действий, таких как «рвать бумагу», независимо от того, выполняет это действие обезьяна или другое животное.[1]
Функция зеркальных нейронов у макак остается неизвестной. Взрослые макаки не учатся путем подражания. Недавние эксперименты Феррари и его коллег показывают, что детеныши макак могут имитировать движения лица человека, но только как новорожденные и в течение ограниченного временного окна.[32] Даже если это еще не было продемонстрировано эмпирически, было высказано предположение, что зеркальные нейроны вызывают такое поведение и другие имитационные явления.[33] В действительности, существует ограниченное понимание того, в какой степени обезьяны демонстрируют подражательное поведение.[8]
У взрослых обезьян зеркальные нейроны могут позволить обезьяне понять, что делает другая обезьяна, или распознать действия другой обезьяны.[34]
В людях
Обычно невозможно изучить отдельные нейроны в человеческом мозге, поэтому большинство доказательств наличия зеркальных нейронов у людей является косвенным. Эксперименты по визуализации мозга с использованием функциональная магнитно-резонансная томография (фМРТ) показали, что человек нижняя лобная кора и верхняя теменная доля активны, когда человек выполняет действие, а также когда человек видит, что другой человек выполняет действие. Было высказано предположение, что эти области мозга содержат зеркальные нейроны, и они были определены как система зеркальных нейронов человека.[35] Более поздние эксперименты показали, что даже на уровне отдельных участников, сканированных с помощью фМРТ, большие области, содержащие несколько вокселей фМРТ, увеличивают их активность как во время наблюдения, так и во время выполнения действий.[27]
Нейропсихологические исследования, изучающие области поражения, которые вызывают знание действия, интерпретацию пантомимы и биологические восприятие движения дефициты указали на причинную связь между целостностью нижней лобной извилины и этим поведением.[36][37][38] Транскраниальная магнитная стимуляция исследования также подтвердили это.[39][40] Эти результаты указывают на то, что активация в областях, связанных с зеркальными нейронами, вряд ли будет просто эпифеноменальной.
В исследовании, опубликованном в апреле 2010 года, сообщается о записях отдельных нейронов с зеркальными свойствами в человеческом мозге.[41] Мукамель и другие. (Current Biology, 2010), записанные в мозге 21 пациента, лечившегося в Медицинском центре Рональда Рейгана UCLA по поводу трудноизлечимой эпилепсии. Пациентам были имплантированы глубинные внутричерепные электроды для выявления очагов припадков для возможного хирургического лечения. Расположение электродов основывалось исключительно на клинических критериях; исследователи с согласия пациентов использовали те же электроды для «совмещения» своих исследований. Исследователи обнаружили небольшое количество нейронов, которые активизировались или проявляли наибольшую активность как при выполнении человеком задачи, так и при наблюдении за ней. Другие нейроны обладали антизеркальными свойствами, то есть они реагировали, когда участник выполнял действие, но подавлялись, когда участник видел это действие.
Обнаруженные зеркальные нейроны были расположены в дополнительной моторной области и медиальной височной коре (другие области мозга не были взяты). По чисто практическим причинам эти области не совпадают с теми, в которых были записаны зеркальные нейроны у обезьяны: исследователи в Парме изучали вентральную премоторную кору и связанную с ней нижнюю теменную долю, две области, в которых эпилепсия возникает редко, и следовательно, регистрация отдельных клеток в этих регионах обычно не выполняется у людей. С другой стороны, до сих пор никому не приходилось искать зеркальные нейроны в дополнительной моторной области или медиальной височной доле у обезьяны. Таким образом, в совокупности это не означает, что у людей и обезьян зеркальные нейроны находятся в разных местах, а скорее, что они могут иметь зеркальные нейроны как в вентральной премоторной коре, так и в нижней теменной доле, где они были зарегистрированы у обезьяны, и в дополнительных. двигательные области и медиальная височная доля, где они были зарегистрированы у человека - особенно потому, что подробный анализ человеческой фМРТ предполагает активность, совместимую с присутствием зеркальных нейронов во всех этих областях.[27]
Другое исследование показало, что у людей не обязательно должно быть больше зеркальных нейронов, чем у обезьян, но вместо этого существует базовый набор зеркальных нейронов, используемых для наблюдения и выполнения действий. Однако для других предполагаемых функций зеркальных нейронов зеркальная система может иметь возможность задействовать другие области мозга при выполнении своих слуховых, соматосенсорных и аффективных компонентов.[42]
У грызунов
Ряд исследований показал, что крысы и мыши проявляют признаки дистресса, когда становятся свидетелями ударов ногами другого грызуна.[43] Группа Кристиан Кейзерс записано с нейронов, в то время как крысы испытывали боль или были свидетелями боли других, и выявило присутствие зеркальных нейронов боли в передней поясной коре крысы, то есть нейронов, которые реагируют как на то, что животное испытывает боль, так и при наблюдении боли других.[44] Деактивация этой области поясной коры привела к уменьшению эмоционального заражения у крыс, так что крысы-наблюдатели демонстрировали уменьшение стресса, когда наблюдали, как другая крыса испытывает боль.[44] Гомологичная часть коры передней поясной извилины у людей ассоциируется с сочувствием к боли,[45] предполагая гомологию между системами, вовлеченными в эмоциональное заражение грызунов и сочувствие боли у людей.
Сомнения относительно зеркальных нейронов
Хотя многие в научном сообществе выразили волнение по поводу открытия зеркальных нейронов, есть ученые, которые выразили сомнения как в существовании, так и в роли зеркальных нейронов у людей. По мнению таких ученых, как Хикок, Пасколо и Динштейн, неясно, действительно ли зеркальные нейроны образуют отдельный класс клеток (в отличие от случайного явления, наблюдаемого в клетках, которые выполняют другие функции).[46] и является ли зеркальная активность отдельным типом реакции или просто артефактом общего облегчения двигательной системы.[9]
В 2008 году Илан Динштейн и др. утверждал, что первоначальный анализ был неубедительным, потому что он был основан на качественном описании свойств отдельных клеток и не принимал во внимание небольшое количество сильно зеркально-избирательных нейронов в двигательных областях.[8] Другие ученые утверждали, что измерения задержки возбуждения нейронов несовместимы со стандартным временем реакции.[46] и указал, что никто не сообщал, что прерывание двигательных областей в F5 приведет к снижению распознавания действий.[9] (Критики этого аргумента ответили, что эти авторы упустили из виду нейропсихологические и психологические исследования человека. ТМС исследования, сообщающие о нарушениях в этих областях, действительно вызывают дефицит действий[37][39] не влияя на другие виды восприятия.)[38]
В 2009 году Lingnau et al. провели эксперимент, в котором сравнивали двигательные акты, которые сначала наблюдались, а затем выполняли, с двигательными актами, которые сначала выполнялись, а затем наблюдались. Они пришли к выводу, что между двумя процессами существует значительная асимметрия, которая указывает на то, что зеркальные нейроны не существуют у людей. Они заявили: «Важно отметить, что мы не обнаружили никаких признаков адаптации для двигательных актов, которые сначала выполнялись, а затем наблюдались. Неспособность найти кросс-модальную адаптацию для выполняемых и наблюдаемых двигательных актов несовместима с основным предположением теории зеркальных нейронов, которое утверждает это действие. распознавание и понимание основаны на двигательной симуляции ".[47] Однако в том же году Kilner et al. показали, что если целенаправленные действия используются в качестве стимулов, как IPL, так и премоторные области демонстрируют подавление повторения между наблюдением и выполнением, которое предсказывается зеркальными нейронами.[48]
В 2009 году Грег Хикок опубликовал обширный аргумент против утверждения о том, что зеркальные нейроны участвуют в понимании действий: «Восемь проблем для понимания действия теории зеркальных нейронов у обезьян и людей». Он пришел к выводу, что «ранняя гипотеза о том, что эти клетки лежат в основе понимания действий, также является интересной и на первый взгляд разумной идеей. Однако, несмотря на ее широкое признание, это предложение никогда не было должным образом проверено на обезьянах, а на людях имеются убедительные эмпирические доказательства. форма физиологической и нейропсихологической (двойной) диссоциации против иска ".[9]
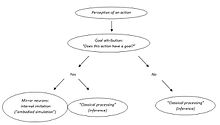
Владимир Косоногов видит еще одно противоречие. Сторонники теории действия зеркальных нейронов с пониманием постулируют, что зеркальные нейроны кодируют цели действий других, потому что они активируются, если наблюдаемое действие является целенаправленным. Однако зеркальные нейроны активируются только тогда, когда наблюдаемое действие является целенаправленным (объектно-направленное действие или коммуникативный жест, который, безусловно, тоже имеет цель). Как они «узнают», что конкретное действие целенаправленно? На каком этапе своей активации они обнаруживают цель движения или ее отсутствие? По его мнению, система зеркальных нейронов может быть активирована только после того, как цель наблюдаемого действия будет приписана какими-то другими структурами мозга.[49]
Нейрофилософы, такие как Патрисия Черчленд выразили как научные, так и философские возражения против теории о том, что зеркальные нейроны отвечают за понимание намерений других. В главе 5 своей книги «Braintrust» 2011 года Черчленд отмечает, что утверждение о том, что зеркальные нейроны участвуют в понимании намерений (посредством моделирования наблюдаемых действий), основано на предположениях, которые омрачены нерешенными философскими проблемами. Она утверждает, что намерения понимаются (кодируются) на более сложном уровне нейронной активности, чем у отдельных нейронов. Черчленд утверждает, что «нейрон, хотя и сложный в вычислительном отношении, это просто нейрон. Это не интеллектуальный гомункул. Если нейронная сеть представляет собой нечто сложное, например намерение [оскорбить], она должна иметь правильный ввод и быть в нужное место в нейронной схеме для этого ". [50]
Недавно Сесилия Хейес (профессор экспериментальной психологии, Оксфорд) выдвинула теорию о том, что зеркальные нейроны являются побочным продуктом ассоциативного обучения, а не эволюционной адаптации. Она утверждает, что зеркальные нейроны у людей являются продуктом социального взаимодействия, а не эволюционной адаптацией к пониманию действия. В частности, Хейес отвергает теорию, выдвинутую В.С. Рамачандран, зеркальные нейроны, были «движущей силой большого скачка в эволюции человека».[10][51]
Разработка
Данные о младенцах человека, использующие меры отслеживания взгляда, показывают, что система зеркальных нейронов развивается до 12-месячного возраста и что эта система может помочь человеческим младенцам понимать действия других людей.[52] Критический вопрос касается того, как зеркальные нейроны приобретают зеркальные свойства. Две тесно связанные модели постулируют, что зеркальные нейроны обучаются через Hebbian[53] или же Ассоциативное обучение[54][55][56] (видеть Ассоциативное последовательное обучение ). Однако, если премоторные нейроны необходимо обучать действием, чтобы приобрести зеркальные свойства, неясно, как новорожденные младенцы могут имитировать мимические жесты другого человека (имитация невидимых действий), как предполагает работа Мельцов и Мур. Одна из возможностей состоит в том, что при виде выпячивания языка врожденный механизм выпуска у новорожденных. Тщательный анализ показывает, что «имитация» этого единственного жеста может быть причиной почти всех сообщений о мимике лица новорожденных.[57]
Возможные функции
Понимание намерений
Многие исследования связывают зеркальные нейроны с пониманием целей и намерений. Fogassi et al. (2005)[58] зарегистрировали активность 41 зеркального нейрона в нижней теменной доле (IPL) двух макак-резусов. IPL долгое время считалась ассоциативной корой головного мозга, которая объединяет сенсорную информацию. Обезьяны наблюдали, как экспериментатор либо хватал яблоко и подносил его ко рту, либо хватал объект и помещал его в чашку.
- В общей сложности 15 зеркальных нейронов активно активировались, когда обезьяна наблюдала движение «хватать, чтобы есть», но не регистрировала никакой активности при воздействии условия «хватание на место».
- Для четырех других зеркальных нейронов справедливо обратное: они активировались в ответ на то, что экспериментатор в конце концов поместил яблоко в чашку, но не съел его.
Только тип действия, а не кинематическая сила, с которой модели управляют объектами, определяют активность нейрона. Также важно отметить, что нейроны сработали до того, как обезьяна увидела, как модель человека запускает второй двигательный акт (поднося объект ко рту или помещая его в чашку). Следовательно, нейроны IPL «кодируют одно и то же действие (захват) по-разному в зависимости от конечной цели действия, в которое он встроен». [58] Они могут предоставить нейронную основу для предсказания последующих действий другого человека и вывода намерений.[58]
Содействие обучению
Другая возможная функция зеркальных нейронов - облегчение обучения. Зеркальные нейроны кодируют конкретное представление действия, то есть представление, которое активировалось бы, если бы наблюдатель действовал. Это позволило бы нам моделировать (внутренне повторять) наблюдаемое действие неявно (в мозгу), чтобы собрать наши собственные моторные программы наблюдаемых действий и подготовиться к воспроизведению этих действий позже. Это неявное обучение. Благодаря этому, наблюдатель будет производить действие явно (в своем поведении) с ловкостью и ловкостью. Это происходит за счет процессов ассоциативного обучения. Чем чаще активируется синаптическая связь, тем сильнее она становится.[49]
Сочувствие
Стефани Престон и Франс де Ваал,[59] Жан Десети,[60][61] и Витторио Галлезе[62][63] и Кристиан Кейзерс[3] независимо утверждали, что система зеркальных нейронов участвует в сочувствие. Большое количество экспериментов с использованием фМРТ, электроэнцефалография (ЭЭГ) и магнитоэнцефалография (МЭГ) показали, что определенные области мозга (в частности, передняя островок, передняя поясная кора, и нижняя лобная кора) активны, когда люди испытывают эмоции (отвращение, счастье, боль и т. д.) и когда видят, что другой человек испытывает эмоции.[64][65][66][67][68][69][70]Дэвид Фридберг и Витторио Галлезе также выдвинули идею, что эта функция системы зеркальных нейронов имеет решающее значение для эстетический опыты.[71] Однако эти области мозга не совсем такие, как те, которые отражают действия рук, а зеркальные нейроны для эмоциональных состояний или эмпатии у обезьян еще не описаны.
Кристиан Кейзерс в Социальная лаборатория мозга и коллеги показали, что люди, которые более эмпатичны в соответствии с анкетами самооценки, имеют более сильную активацию как в зеркальной системе для действий руками[72] и зеркальная система эмоций,[69] Обеспечивая более прямую поддержку идеи о том, что зеркальная система связана с эмпатией. Некоторые исследователи отметили, что человеческая зеркальная система не пассивно реагирует на наблюдение за действиями, а находится под влиянием мышления наблюдателя.[73] Исследователи наблюдали связь зеркальных нейронов во время чуткого взаимодействия с пациентом.[74]
Исследования на крысах показали, что передняя поясная кора содержит зеркальные нейроны для боли, то есть нейроны, реагирующие как на собственное переживание боли, так и при наблюдении за болью других,[44] и ингибирование этой области приводит к уменьшению эмоциональное заражение у крыс[44] и мыши,[43] и снижение отвращения к причинению вреда другим.[75] Это дает причинные доказательства связи между зеркальными нейронами боли и эмоциональное заражение и просоциальное поведение, два явления, связанные с эмпатией, у грызунов. Эта активность мозга в гомологичной области мозга связана с индивидуальной вариабельностью эмпатии у людей.[45] предполагает, что аналогичный механизм может действовать у млекопитающих.
Самосознание человека
В. С. Рамачандран предположил, что зеркальные нейроны могут обеспечить неврологическую основу человеческого самосознания.[76] В эссе, написанном для Edge Foundation в 2009 году, Рамачандран дал следующее объяснение своей теории: «... Я также предположил, что эти нейроны могут не только помогать имитировать поведение других людей, но и могут быть обращены« внутрь », так сказать, на создавать представления второго порядка или мета-представления вашего собственный более ранние мозговые процессы. Это может быть нейронная основа самоанализа и взаимности самосознания и другого осознания. Очевидно, здесь возникает вопрос о курице или яйце относительно того, что эволюционировало первым, но ... Суть в том, что двое эволюционировали вместе, взаимно обогащая друг друга, чтобы создать зрелое представление о себе, которое характеризует современных людей ». [77]
Язык
В исследованиях функциональной МРТ у людей были обнаружены области, гомологичные системе зеркальных нейронов обезьян, в нижней лобной коре, недалеко от области Брока, одной из предполагаемых языковых областей мозга. Это привело к предположению, что человеческий язык эволюционировал из системы представления / понимания жестов, реализованной в зеркальных нейронах. Считается, что зеркальные нейроны могут обеспечить механизм понимания действий, имитационного обучения и моделирования поведения других людей.[78] Эта гипотеза подтверждается некоторыми цитоархитектонический гомологии между премоторной областью F5 обезьяны и областью Брока человека.[79] Ставки словарный запас ссылка расширения на способность дети чтобы вокально отражать не-слова и, таким образом, усваивать новое произношение. Такой повторение речи происходит автоматически, быстро[80] и отдельно в мозгу восприятие речи.[81][82] Более того, такая вокальная имитация может происходить без понимания, например, в затенение речи[83] и эхолалия.[84]
Дополнительное доказательство этой связи получено в недавнем исследовании, в котором активность мозга двух участников была измерена с помощью фМРТ, когда они жестикулировали словами друг другу, используя жесты рук с игрой в шарады - модальность, которую некоторые предлагали, может представлять собой эволюционного предшественника человеческого языка. Анализ данных с использованием Причинность Грейнджера показали, что зеркально-нейронная система наблюдателя действительно отражает паттерн активности моторной системы отправителя, подтверждая идею о том, что моторная концепция, связанная со словами, действительно передается от одного мозга к другому с использованием зеркальной системы.[85]
Система зеркальных нейронов кажется по своей сути неадекватной, чтобы играть какую-либо роль в синтаксис, учитывая, что это определяющее свойство человеческих языков, которое реализовано в иерархической рекурсивной структуре, уплощено в линейные последовательности фонем, что делает рекурсивную структуру недоступной для сенсорного обнаружения[86]
Автоматическая имитация
Этот термин обычно используется для обозначения случаев, когда человек, наблюдая за движением тела, непреднамеренно выполняет подобное движение тела или изменяет способ выполнения движения тела. Автоматическая имитация редко включает в себя открытое выполнение соответствующих ответов. Вместо этого эффекты обычно состоят из времени реакции, а не точности, различий между совместимыми и несовместимыми испытаниями. Исследования показывают, что существование автоматической имитации, которая является скрытой формой имитации, отличается от пространственной совместимости. Это также указывает на то, что, хотя автоматическая имитация подвержена входной модуляции процессами внимания и выходной модуляции тормозными процессами, она опосредуется усвоенными долгосрочными сенсомоторными ассоциациями, которые не могут быть изменены непосредственно намеренными процессами. Многие исследователи полагают, что автоматическая имитация опосредована системой зеркальных нейронов.[87] Кроме того, есть данные, которые демонстрируют, что наш контроль позы нарушается, когда люди слушают предложения о других действиях. Например, если задача - сохранить осанку, люди делают это хуже, когда слушают такие предложения: «Я встаю, надеваю тапочки, иду в ванную». Это явление может быть связано с тем, что при восприятии действия происходит аналогичная активация моторной коры, как если бы такое же действие выполнял человек (система зеркальных нейронов).[88]
Мимикрия мотора
В отличие от автоматической имитации, двигательная мимика наблюдается в (1) натуралистических социальных ситуациях и (2) посредством измерения частоты действий в течение сеанса, а не измерения скорости и / или точности в рамках испытаний.[89]
Объединение исследований моторной мимикрии и автоматической имитации может выявить правдоподобные признаки того, что эти явления зависят от одних и тех же психологических и нервных процессов. Однако предварительные данные получены из исследований, показывающих, что социальная прайминг оказывает аналогичное влияние на моторную мимикрию.[90][91]
Тем не менее, сходство между автоматической имитацией, зеркальными эффектами и моторной мимикрией побудило некоторых исследователей предположить, что автоматическая имитация опосредуется системой зеркальных нейронов и что это строго контролируемый лабораторный эквивалент моторной мимикрии, наблюдаемой в натуралистических социальных контекстах. Если это правда, то автоматическая имитация может использоваться как инструмент для исследования того, как система зеркальных нейронов способствует когнитивному функционированию и как моторная мимикрия способствует просоциальным установкам и поведению.[92][93]
Мета-анализ имитационных исследований на людях показывает, что существует достаточно доказательств активации зеркальной системы во время имитации, что вероятно вовлечение зеркальных нейронов, даже несмотря на то, что никакие опубликованные исследования не зафиксировали активность отдельных нейронов. Однако для имитации двигателя этого, вероятно, недостаточно. Исследования показывают, что области лобной и теменной долей, выходящие за пределы классической системы зеркал, одинаково активируются во время имитации. Это говорит о том, что другие области, наряду с системой зеркал, имеют решающее значение для имитации поведения.[6]
Аутизм
Также было высказано предположение, что проблемы с системой зеркальных нейронов могут лежать в основе когнитивных расстройств, в частности аутизм.[94][95] Однако связь между дисфункцией зеркальных нейронов и аутизмом является предварительной, и еще предстоит продемонстрировать, как зеркальные нейроны связаны со многими важными характеристиками аутизма.[8]
Некоторые исследователи утверждают, что существует связь между дефицитом зеркальных нейронов и аутизм. Записи ЭЭГ из моторных областей подавляются, когда кто-то наблюдает за движением другого человека, сигнал, который может относиться к системе зеркальных нейронов. Это подавление было меньше у детей с аутизмом.[94] Хотя эти результаты были воспроизведены несколькими группами,[96][97] другие исследования не обнаружили доказательств дисфункциональной системы зеркальных нейронов при аутизме.[8] В 2008 году Оберман и др. опубликовал исследовательскую работу, в которой представлены противоречивые данные ЭЭГ. Оберман и Рамачандран обнаружили типичное подавление мю для знакомых стимулов, но не для незнакомых, что привело их к выводу, что система зеркальных нейронов детей с РАС (расстройством аутистического спектра) функционирует, но менее чувствительна, чем у обычных детей.[98] Основываясь на противоречивых доказательствах, представленных экспериментами по подавлению мю-волн, Патрисия Черчленд предупредил, что результаты подавления мю-волн нельзя использовать в качестве действительного показателя для измерения производительности систем зеркальных нейронов.[99] Недавние исследования показывают, что зеркальные нейроны не играют роли при аутизме:
... нет явных доказательств фундаментального дефицита зеркальной системы при аутизме. Поведенческие исследования показали, что люди с аутизмом хорошо понимают цели действий. Кроме того, два независимых исследования нейровизуализации сообщили, что париетальный компонент зеркальной системы обычно функционирует у людей с аутизмом.[100]
Некоторые анатомические различия были обнаружены в областях мозга, связанных с зеркальными нейронами, у взрослых с расстройствами аутистического спектра по сравнению со взрослыми, не страдающими аутизмом. Все эти корковые области были тоньше, и степень истончения коррелировала с тяжестью симптомов аутизма, корреляция почти ограничивалась этими областями мозга.[101] Основываясь на этих результатах, некоторые исследователи утверждают, что аутизм вызывается нарушениями в системе зеркальных нейронов, что приводит к нарушениям социальных навыков, имитации, сочувствия и теории разума.[ВОЗ? ]
Многие исследователи отмечают, что теория аутизма о «разбитых зеркалах» слишком упрощена, и одни только зеркальные нейроны не могут объяснить различия, обнаруживаемые у людей с аутизмом. Прежде всего, как отмечалось выше, ни одно из этих исследований не было прямым измерением активности зеркальных нейронов - другими словами, активность фМРТ или подавление ритма ЭЭГ не однозначно индексируют зеркальные нейроны. Динштейн и его коллеги обнаружили нормальную активность зеркальных нейронов у людей с аутизмом с помощью фМРТ.[102] У людей с аутизмом не всегда обнаруживается дефицит понимания намерения, понимания действий и биологического восприятия движения (ключевые функции зеркальных нейронов).[103][104] или зависят от задачи.[105][106] Сегодня очень немногие люди верят, что проблема «все или ничего» с зеркальной системой может лежать в основе аутизма. Вместо этого «необходимо провести дополнительное исследование и проявлять большую осторожность при обращении к СМИ». [107]
Исследования 2010 г.[108] пришли к выводу, что аутичные люди не проявляют дисфункции зеркальных нейронов, хотя небольшой размер выборки ограничивает степень, в которой эти результаты могут быть обобщены.
Теория разума
В Философия разума, зеркальные нейроны стали главным призывом к сплочению теоретики моделирования относительно нашего "теория разума. »« Теория разума »относится к нашей способности делать выводы о психическом состоянии другого человека (то есть, убеждениях и желаниях) на основании опыта или их поведения.
Есть несколько конкурирующих моделей, которые пытаются объяснить нашу теорию разума; Наиболее примечательным в отношении зеркальных нейронов является теория моделирования. Согласно теории симуляции, теория разума доступна, потому что мы подсознательно сопереживайте человеку, за которым мы наблюдаем, и, учитывая соответствующие различия, представляйте, чего бы мы хотели и во что верили в этом сценарии.[109][110] Зеркальные нейроны интерпретировались как механизм, с помощью которого мы моделируем других, чтобы лучше понять их, и поэтому их открытие было воспринято некоторыми как подтверждение теории моделирования (которая появилась за десять лет до открытия зеркальных нейронов).[62] Совсем недавно теория разума и моделирование рассматривались как взаимодополняющие системы с разными временными курсами развития.[111][112][113]
На уровне нейронов, в исследовании 2015 года, проведенном Кереном Харушем и Зивом Уильямсом с использованием совместно взаимодействующих приматов, выполняющих повторяющуюся игру «дилемма заключенного», авторы идентифицировали нейроны в передняя поясная кора которая выборочно предсказывала еще неизвестные решения или скрытое состояние оппонента. Эти «нейроны, предсказывающие других», различали собственные решения и другие решения и были уникально чувствительны к социальному контексту, но они не кодировали наблюдаемые действия оппонента или получение вознаграждения. Таким образом, эти поясные клетки могут существенно дополнять функцию зеркальных нейронов, предоставляя дополнительную информацию о других социальных агентах, которая не является незамедлительной наблюдаемой или известной.[114]
Половые различия
Серия недавних исследований, проведенных Yawei Cheng с использованием различных нейрофизиологических показателей, в том числе: МЭГ,[115] спинномозговая рефлекторная возбудимость,[116] электроэнцефалография,[117][118] документально подтвердили наличие гендерных различий в системе зеркальных нейронов человека: участники женского пола демонстрируют более сильный моторный резонанс, чем участники мужского пола.
В другом исследовании половые различия между механизмами зеркальных нейронов были подтверждены тем, что данные показали повышенную способность к сочувствию у женщин по сравнению с мужчинами. Во время эмоционального социального взаимодействия женщины продемонстрировали большую способность воспринимать эмоциональную перспективу, чем мужчины, при общении с другим человеком лицом к лицу. Однако данные исследования показали, что когда дело дошло до распознавания эмоций других, способности всех участников были очень похожи, и не было ключевой разницы между мужчинами и женщинами.[119]
Сонный паралич / Призрачные нарушители спальни
Баланд Джалал и В. С. Рамачандран выдвинули гипотезу, что система зеркальных нейронов играет важную роль в возникновении галлюцинаций злоумышленников и внетелесных переживаний во время сонный паралич.[120] Согласно этой теории, сонный паралич приводит к растормаживанию системы зеркальных нейронов, открывая путь для галлюцинаций человекоподобных теневых существ. В качестве механизма такого растормаживания зеркального нейрона предлагается деафферентация сенсорной информации во время паралича сна.[120] Авторы предполагают, что их гипотеза о роли системы зеркальных нейронов может быть проверена:
«Эти идеи можно было бы изучить с помощью нейровизуализации, чтобы изучить избирательную активацию областей мозга, связанных с активностью зеркальных нейронов, когда человек галлюцинирует злоумышленника или испытывает внетелесный опыт во время сонного паралича».[120]
Функция зеркального нейрона, психоз и эмпатия при шизофрении
Недавнее исследование, в котором измерялось подавление мю-волн, предполагает, что активность зеркальных нейронов положительно коррелирует с психотическими симптомами (то есть большее подавление мю / активность зеркальных нейронов было самым высоким среди субъектов с большей серьезностью психотических симптомов). Исследователи пришли к выводу, что «более высокая активность зеркальных нейронов может лежать в основе дефицита сенсорного стробирования при шизофрении и может способствовать сенсорной неправильной атрибуции, особенно в ответ на социально значимые стимулы, и быть предполагаемым механизмом бреда и галлюцинаций».[121]
Смотрите также
- Ассоциативное последовательное обучение
- Общая теория кодирования
- Эмоциональное заражение
- Сочувствие
- Синестезия с зеркальным касанием
- Зеркальное отображение (психология)
- Моторное познание
- Двигательная теория восприятия речи
- Об интеллекте
- Позитронно-эмиссионная томография
- Имитационная теория эмпатии
- Повторение речи
- Нейрон веретена
Рекомендации
- ^ а б c d Риццолатти, Джакомо; Крейгеро, Лейла (2004). «Зеркало-нейронная система» (PDF). Ежегодный обзор нейробиологии. 27 (1): 169–192. Дои:10.1146 / annurev.neuro.27.070203.144230. PMID 15217330.
- ^ Кейзерс, Кристиан (2010). «Зеркальные нейроны» (PDF). Текущая биология. 19 (21): R971–973. Дои:10.1016 / j.cub.2009.08.026. PMID 19922849. S2CID 12668046. Архивировано из оригинал (PDF) 19 января 2013 г.
- ^ а б c d Кейзерс, Кристиан (23.06.2011). Эмпатический мозг. Kindle.
- ^ а б Риццолатти, Джакомо; Фадига, Лучано (1999). «Резонансное поведение и зеркальные нейроны». Italiennes de Biologie. 137 (2–3): 85–100. PMID 10349488.
- ^ Prather, J. F .; Peters, S .; Новицки, С .; Муни, Р. (2008). «Точное слухово-голосовое зеркальное отображение в нейронах для обучения голосовой коммуникации». Природа. 451 (7176): 305–310. Bibcode:2008Натура.451..305П. Дои:10.1038 / природа06492. PMID 18202651. S2CID 4344150.
- ^ а б Моленбергс П., Каннингтон Р., Маттингли Дж. Б. (июль 2009 г.). «Участвует ли система зеркальных нейронов в имитации? Краткий обзор и метаанализ». Неврология и биоповеденческие обзоры. 33 (7): 975–80. Дои:10.1016 / j.neubiorev.2009.03.010. PMID 19580913. S2CID 25620637.
- ^ Акинс, Хана; Кляйн, Эдвард (2002). «Имитационное обучение японского перепела с использованием процедуры двунаправленного контроля». Обучение и поведение животных. 30 (3): 275–281. Дои:10.3758 / bf03192836. PMID 12391793.
- ^ а б c d е Динштейн I, Томас С., Берманн М., Хигер Д.Д. (2008). «Зеркало природы». Curr Biol. 18 (1): R13–8. Дои:10.1016 / j.cub.2007.11.004. ЧВК 2517574. PMID 18177704.
- ^ а б c d Хикок Г (2009). «Восемь проблем для понимания теории действия зеркальных нейронов у обезьян и людей». J Cogn Neurosci. 21 (7): 1229–1243. Дои:10.1162 / jocn.2009.21189. ЧВК 2773693. PMID 19199415.
- ^ а б Привет, Сесилия (2009). "Откуда берутся зеркальные нейроны?" (PDF). Неврология и биоповеденческие обзоры. Архивировано из оригинал (PDF) на 2012-04-26. Получено 2011-12-20.
- ^ Феррари, ПФ; Риццолатти, G (2014). «Исследование зеркальных нейронов: прошлое и будущее». Philos Trans R Soc Lond B Biol Sci. 369 (1644): 20130169. Дои:10.1098 / rstb.2013.0169. ЧВК 4006175. PMID 24778369.
- ^ Кейзерс, Кристиан; Газзола, Валерия (2006). «Прогресс в исследованиях мозга» (PDF). Bcn-nic.nl. Архивировано из оригинал (PDF) 30 июня 2007 г.
- ^ Михаил Арбиб, Гипотеза зеркальной системы. Связь языка с теорией разума, 2005, получено 17 февраля 2006 г. В архиве 29 марта 2009 г. Wayback Machine
- ^ Теорет, Гюго; Паскуаль-Леоне, Альваро (2002). «Приобретение языка: делай, как слышишь». Текущая биология. 12 (21): R736–7. Дои:10.1016 / S0960-9822 (02) 01251-4. PMID 12419204.
- ^ Якобони, Марко (22 февраля 2005 г.). "Улавливание чужих намерений с помощью собственной зеркальной нейронной системы". PLOS Биология. 3 (3): e79. Дои:10.1371 / journal.pbio.0030079. ЧВК 1044835. PMID 15736981.
- ^ Блейксли, Сандра (10 января 2006 г.). «Клетки, читающие мысли». Нью-Йорк Таймс | Наука.
- ^ Джарретт, Кристиан, Спокойный взгляд на самую распространенную концепцию нейронауки - зеркальные нейроны, Wired, 12.13.13,[1]
- ^ Ди Пеллегрино, G .; Fadiga, L .; Fogassi, L .; Галлезе, В .; Риццолатти, G (1992). «Понимание моторных событий: нейрофизиологическое исследование». Экспериментальное исследование мозга. 91 (1): 176–180. Дои:10.1007 / bf00230027. PMID 1301372. S2CID 206772150.
- ^ Риццолатти, Джакомо; Фадига, Лучано; Галлезе, Витторио; Фогасси, Леонардо (1996). «Премоторная кора и распознавание двигательных действий». Когнитивные исследования мозга. 3 (2): 131–141. Дои:10.1016/0926-6410(95)00038-0. PMID 8713554.
- ^ Rizzolatti G, Fabbri-Destro M (январь 2010 г.). «Зеркальные нейроны: от открытия до аутизма». Экспериментальное исследование мозга. 200 (3–4): 223–37. Дои:10.1007 / s00221-009-2002-3. PMID 19760408. S2CID 3342808.
- ^ Галлезе, В .; Fadiga, L .; Fogassi, L .; Риццолатти, Джакомо (1996). «Распознавание действий в премоторной коре». Мозг. 119 (2): 593–609. Дои:10.1093 / мозг / 119.2.593. PMID 8800951.
- ^ Феррари; и другие. (2003). «Зеркальные нейроны, отвечающие на наблюдение за пищевыми и коммуникативными действиями во рту в вентральной премоторной коре». Европейский журнал нейробиологии. 17 (8): 1703–1714. Дои:10.1046 / j.1460-9568.2003.02601.x. PMID 12752388. S2CID 1915143.
- ^ Колер; и другие. (2002). "Наука" (PDF). Bcn-nic.nl. Архивировано из оригинал (PDF) 30 июня 2007 г.
- ^ Газзола; и другие. (2006). «Современная биология» (PDF). Bcn-nic.nl. Архивировано из оригинал (PDF) 30 июня 2007 г.
- ^ Галлезе, Витторио; Фадига, Лучано; Фогасси, Леонардо; Риццолатти, Джакомо (1996). «Распознавание действий в премоторной коре». Мозг. 119 (2): 593–609. Дои:10.1093 / мозг / 119.2.593. PMID 8800951.
- ^ Fogassi et al., Теменная доля: от организации действия к пониманию намерений, Наука, 2005
- ^ а б c Gazzola, V .; Кейзерс, К. (2009). «Наблюдение и выполнение действий имеют общие моторные и соматосенсорные вокселы у всех испытуемых: анализ несглаженных данных фМРТ по отдельности». Кора головного мозга. 19 (6): 1239–1255. Дои:10.1093 / cercor / bhn181. ЧВК 2677653. PMID 19020203.
- ^ Кейзерс, Кристиан; Каас, Джон; Газзола, Валерия (2010). «Соматосенсация в социальном познании» (PDF). Обзоры природы Неврология. 11 (6): 417–28. Дои:10.1038 / номер 2833. PMID 20445542. S2CID 12221575. Архивировано из оригинал (PDF) на 16.08.2010.
- ^ а б Кук Р., Птица Дж., Катмур С., Пресс С., Привет С. (2014). «Зеркальные нейроны: от истоков к функциям». Behav Brain Sci. 37 (2): 177–92. Дои:10.1017 / S0140525X13000903. PMID 24775147.CS1 maint: несколько имен: список авторов (связь)
- ^ Кейзерс С., Газзола В. (2014). «Хеббийское обучение и прогнозирующие зеркальные нейроны для действий, ощущений и эмоций». Philos Trans R Soc Lond B Biol Sci. 369 (1644): 20130175. Дои:10.1098 / rstb.2013.0175. ЧВК 4006178. PMID 24778372.
- ^ Дель Джудис М., Манера В., Кейзерс К. (2009). «Запрограммирован на обучение? Онтогенез зеркальных нейронов». Dev Sci. 12 (2): 350–63. Дои:10.1111 / j.1467-7687.2008.00783.x. HDL:2318/133096. PMID 19143807.CS1 maint: несколько имен: список авторов (связь)
- ^ Ferrari, P. F .; Visalberghi, E .; Paukner, A .; Fogassi, L .; Ruggiero, A .; Суоми, SJ; и другие. (2006). «Неонатальная имитация у макак-резусов». PLOS Биология. 4 (9): e302. Дои:10.1371 / journal.pbio.0040302. ЧВК 1560174. PMID 16953662.
- ^ Феррари П.Ф., Бонини Л., Фогасси Л. (2009) От зеркальных нейронов обезьян к зеркальному поведению: возможные прямые и косвенные пути. Философские труды Королевского общества, Б. .364, 2311-2323.
- ^ Риццолатти, Джакомо; Арбиб, Майкл А. (1998). «Язык в пределах нашей досягаемости». Тенденции в неврологии. 21 (5): 188–194. Дои:10.1016 / S0166-2236 (98) 01260-0. HDL:11858 / 00-001M-0000-002C-4B59-6. PMID 9610880. S2CID 679023.
- ^ Якобони, Марко; Вудс, Роджер П .; Латунь, Марсель; Беккеринг, Гарольд; Mazziotta, John C .; Риццолатти, Джакомо (1999). «Корковые механизмы имитации человека» (PDF). Наука. 286 (5449): 2526–2528. Дои:10.1126 / science.286.5449.2526. PMID 10617472.
- ^ Сайгин А.П., Уилсон С.М., Дронкерс Н.Ф., Бейтс Э. (2004). «Понимание действий при афазии: лингвистические и неязыковые дефициты и их взаимосвязи». Нейропсихология. 42 (13): 1788–804. CiteSeerX 10.1.1.544.9071. Дои:10.1016 / j.neuropsychologia.2004.04.016. PMID 15351628. S2CID 11622224.
- ^ а б Tranel, D .; Kemmerer, D .; Adolphs, R .; Damasio, H .; Дамасио, А. Р. (01.05.2003). «Нейронные корреляты концептуального знания для действий». Когнитивная нейропсихология. 20 (3): 409–432. Дои:10.1080/02643290244000248. PMID 20957578. S2CID 16131.
- ^ а б Сайгин, А. П. (2007). «Верхние височные и премоторные области мозга, необходимые для биологического восприятия движения». Мозг: журнал неврологии. 130 (Pt 9): 2452–2461. Дои:10.1093 / мозг / awm162. PMID 17660183.
- ^ а б Pobric, G .; De c. Гамильтон, А. Ф. Д. С. (07.03.2006). «Понимание действий требует левой нижней лобной коры». Текущая биология. 16 (5): 524–529. Дои:10.1016 / j.cub.2006.01.033. PMID 16527749.
- ^ Candidi M; Urgesi C; Ionta S; Аглиоти С.М. (2008). «Виртуальное поражение вентральной премоторной коры ухудшает зрительное восприятие биомеханически возможных, но не невозможных действий». Социальная неврология. 3 (3–4): 388–400. Дои:10.1080/17470910701676269. PMID 18979387. S2CID 37390465.
- ^ Кейзерс, Кристиан; Газзола, Валерия (2010). "Социальная неврология: зеркальные нейроны, зарегистрированные у людей" (PDF). Текущая биология. 20 (8): R353–354. Дои:10.1016 / j.cub.2010.03.013. HDL:20.500.11755 / 351f0172-b06b-41de-a013-852c64e197fa. PMID 21749952. S2CID 3609747. Архивировано из оригинал (PDF) на 16.08.2010.
- ^ Моленбергс П., Каннингтон Р., Маттингли Дж. Б. (январь 2012 г.). «Области мозга с зеркальными свойствами: метаанализ 125 исследований человеческой фМРТ» (PDF). Неврология и биоповеденческие обзоры. 36 (1): 341–9. Дои:10.1016 / j.neubiorev.2011.07.004. PMID 21782846. S2CID 37871374.
- ^ а б Кеум С., Шин Х.С. (2019). «Нейронная основа изучения страха при наблюдении: потенциальная модель аффективной эмпатии». Нейрон. 104 (1): 78–86. Дои:10.1016 / j.neuron.2019.09.013. PMID 31600517. S2CID 203932127.
- ^ а б c d Каррильо М., Хан Й., Миглиорати Ф, Лю М., Газзола В., Кейзерс С. (2019). «Эмоциональные зеркальные нейроны в передней поясной коре головного мозга крысы». Curr Biol. 29 (8): 1301–1312.e6. Дои:10.1016 / j.cub.2019.03.024. ЧВК 6488290. PMID 30982647.CS1 maint: несколько имен: список авторов (связь)
- ^ а б Ламм К., Десети Дж., Певец Т. (2011). «Метааналитические данные об общих и различных нейронных сетях, связанных с непосредственным переживанием боли и сочувствием к боли». NeuroImage. 54 (3): 2492–502. Дои:10.1016 / j.neuroimage.2010.10.014. PMID 20946964. S2CID 6021487.CS1 maint: несколько имен: список авторов (связь)
- ^ а б Пасколо ПБ, Рагогна Р., Росси Р. (2009). «Парадигма системы зеркало-нейрон и ее последовательность». Походка и поза. 30 (Прил. 1): 65. Дои:10.1016 / j.gaitpost.2009.07.064.
- ^ Lingnau, A .; Gesierich, B .; Карамазза, А. (28 февраля 2009 г.). «Асимметричная адаптация фМРТ не обнаруживает доказательств наличия зеркальных нейронов у людей». Труды Национальной академии наук. 106 (24): 9925–9930. Bibcode:2009PNAS..106.9925L. Дои:10.1073 / pnas.0902262106. ЧВК 2701024. PMID 19497880.
- ^ Килнер, JM .; Нил, А. (12 августа 2009 г.). «Доказательства наличия зеркальных нейронов в нижней лобной извилине человека». J Neurosci. 29 (32): 10153–10159. Дои:10.1523 / jneurosci.2668-09.2009. ЧВК 2788150. PMID 19675249.
- ^ а б Косоногов, В. (2012). «Почему зеркальные нейроны не могут поддерживать понимание действий». Нейрофизиология. 44 (6): 499–502. Дои:10.1007 / s11062-012-9327-4. ISSN 0090-2977. S2CID 6315381.
- ^ Черчленд, Патрисия, Braintrust (2011), Глава 6, стр. 142
- ^ Рамачандран, VS (2000). «Зеркальные нейроны и имитационное обучение как движущая сила« большого скачка вперед »в эволюции человека». Край. Получено 13 апреля 2013.
- ^ Falck-Ytter T, Gredebäck G, von Hofsten C (2006). «Младенцы предсказывают цели действий других людей» (PDF). Природа Неврология 9. Psyk.uu.se. Архивировано из оригинал (PDF) на 25 марта 2009 г.
- ^ Кейзерс и Перретт, Тенденции в когнитивных науках 8 (2004)
- ^ Хейес К. М. (2001). «Причины и последствия имитации». Тенденции в когнитивных науках. 5 (6): 253–261. Дои:10.1016 / S1364-6613 (00) 01661-2. PMID 11390296. S2CID 15602731.
- ^ Брасс, М., и Хейес, К. Тенденции в когнитивных науках 9 (2005)
- ^ Хейес К. М. (2010). «Откуда берутся зеркальные нейроны?». Неврология и биоповеденческие обзоры. 34 (4): 575–583. Дои:10.1016 / j.neubiorev.2009.11.007. PMID 19914284. S2CID 2578537.
- ^ Анисфельд М (1996). «Новорожденным подходит только моделирование оттопыренного языка». Обзор развития. 16 (2): 149–161. Дои:10.1006 / древ.1996.0006.
- ^ а б c Fogassi L, Ferrari PF, Gesierich B, Rozzi S, Chersi F, Rizzolatti G (апрель 2005 г.). «Теменная доля: от организации действия к пониманию намерения». Наука. 308 (5722): 662–7. Bibcode:2005Наука ... 308..662F. Дои:10.1126 / science.1106138. PMID 15860620. S2CID 5720234.
- ^ Preston, S.D .; де Ваал, F.B.M. (2002). «Сочувствие: его конечная и ближайшая основы». Поведенческие науки и науки о мозге. 25 (1): 1–72. CiteSeerX 10.1.1.554.2794. Дои:10.1017 / s0140525x02000018. PMID 12625087.
- ^ Десети, Дж. (2002). Naturaliser l’empathie [Сочувствие натурализованное]. L’Encéphale, 28, 9–20.
- ^ Десети Дж, Джексон П.Л. (2004). «Функциональная архитектура человеческого сочувствия». Обзоры поведенческой и когнитивной нейробиологии. 3 (2): 71–100. Дои:10.1177/1534582304267187. PMID 15537986.
- ^ а б Галлезе В., Гольдман А. (декабрь 1998 г.). «Зеркальные нейроны и имитационная теория чтения мыслей». Тенденции в когнитивных науках. 2 (12): 493–501. Дои:10.1016 / с 1364-6613 (98) 01262-5. PMID 21227300. S2CID 10108122.
- ^ Галлезе V (2001). «Гипотеза« общего многообразия »: от зеркальных нейронов к эмпатии». Журнал исследований сознания. 8: 33–50.
- ^ Ботвиник М., Джа А.П., Былсма Л.М., Фабиан С.А., Соломон П.Е., Пркачин К.М. (2005). «Наблюдение за выражением боли на лице затрагивает области коры головного мозга, участвующие в непосредственном переживании боли». NeuroImage. 25 (1): 312–319. Дои:10.1016 / j.neuroimage.2004.11.043. PMID 15734365. S2CID 24988672.
- ^ Ченг Й, Ян Си, Линь С.П., Ли П.Л., Десети Дж. (Май 2008 г.). «Восприятие боли другими людьми подавляет соматосенсорные колебания: исследование магнитоэнцефалографии». NeuroImage. 40 (4): 1833–40. Дои:10.1016 / j.neuroimage.2008.01.064. PMID 18353686. S2CID 1827514.
- ^ Моррисон, I .; Lloyd, D .; Ди Пеллегрино, G .; Робертс, Н. (2004). «Заместительные реакции на боль в передней поясной коре головного мозга: сопереживание - это мультисенсорная проблема?». Когнитивная, аффективная и поведенческая нейронауки. 4 (2): 270–8. Дои:10.3758 / CABN.4.2.270. PMID 15460933.
- ^ Плетеные; и другие. (2003). «Нейрон» (PDF). Архивировано из оригинал (PDF) 30 июня 2007 г.
- ^ Певица; и другие. (2004). "Наука". Sciencemag.org. Цитировать журнал требует
| журнал =(помощь) - ^ а б Джабби, Мбемба; Сварт, Марте; Кейзерс, Кристиан (2007). «Сочувствие к положительным и отрицательным эмоциям во вкусовой коре». NeuroImage. 34 (4): 1744–53. Дои:10.1016 / j.neuroimage.2006.10.032. PMID 17175173. S2CID 13988152.
- ^ Ламм С., Batson CD, Decety J (январь 2007 г.). «Нейронный субстрат человеческого сочувствия: эффекты перспективного взгляда и когнитивной оценки». Журнал когнитивной неврологии. 19 (1): 42–58. Дои:10.1162 / jocn.2007.19.1.42. PMID 17214562. S2CID 2828843.
- ^ Фридберг, Дэвид; Галлезе, Витторио (2007). «Движение, эмоции и эмпатия в эстетическом опыте». Тенденции в когнитивных науках. 11 (5): 197–203. Дои:10.1016 / j.tics.2007.02.003. PMID 17347026. S2CID 1996468.
- ^ Газзола, Азиз-Заде и Кейзерс (2006). «Современная биология» (PDF). Архивировано из оригинал (PDF) 30 июня 2007 г.
- ^ Моленбергс; и другие. (2 января 2012 г.). «Образцы активации во время наблюдения за действиями модулируются контекстом в областях зеркальной системы» (PDF). NeuroImage. 59 (1): 608–15. Дои:10.1016 / j.neuroimage.2011.07.080. PMID 21840404. S2CID 13951700.
- ^ Ходжат, Мохаммадреза; и другие. "Американский журнал качества медицины | 28.1 (2013 г.)" (PDF). С. 6–7.
- ^ Эрнандес-Лаллемент Дж., Аттах А.Т., Сойман Э., Пинхал С.М., Газзола В., Кейзерс С. (2020). «Вред другим действует как отрицательное подкрепление у крыс». Curr Biol. 30 (6): 949–961.e7. Дои:10.1016 / j.cub.2020.01.017. PMID 32142701. S2CID 212424287.CS1 maint: несколько имен: список авторов (связь)
- ^ Оберман, Л .; Рамачандран, В. (2009). «Отражения на зеркальной нейронной системе: их эволюционные функции за пределами моторного представления». В Пинеде, Дж. (ред.). Зеркальные нейронные системы: роль зеркальных процессов в социальном познании. Humana Press. стр.39 –62. ISBN 978-1-934115-34-3.
- ^ Рамачандран, В. (1 января 2009 г.). "Самосознание: последний рубеж, веб-эссе Edge Foundation". Получено 26 июля, 2011.
- ^ Скойлс, Джон Р., Жест, происхождение языка и праворукость, Психология: 11, # 24, 2000.
- ^ Петридес, Майкл, Кадорет, Женевьева, Макки, Скотт (2005). Орофациальные соматомоторные реакции у макак-гомологов из области Брока, природа: 435, № 1235
- ^ Porter Jr, R.J .; Любкер, Дж. Ф. (1980). «Быстрое воспроизведение последовательностей гласных и гласных: свидетельство быстрой и прямой акустико-моторной связи в речи». Журнал исследований речи и слуха. 23 (3): 593–602. Дои:10.1044 / jshr.2303.593. PMID 7421161.
- ^ McCarthy, R .; Уоррингтон, Э. К. (1984). «Двухмаршрутная модель речевого производства. Свидетельства афазии». Мозг: журнал неврологии. 107 (2): 463–485. Дои:10.1093 / мозг / 107.2.463. PMID 6722512.
- ^ McCarthy, R.A .; Уоррингтон, Э. К. (2001). «Повторение без семантики: поверхностная дисфазия?». Нейроказ. 7 (1): 77–87. Дои:10.1093 / neucas / 7.1.77. PMID 11239078. S2CID 12988855.
- ^ Марслен-Уилсон, В. (1973). «Лингвистическая структура и слежка за речью с очень короткими задержками». Природа. 244 (5417): 522–523. Bibcode:1973Натура.244..522М. Дои:10.1038 / 244522a0. PMID 4621131. S2CID 4220775.
- ^ Fay, W.H .; Коулман, Р. О. (1977). «Человеческий преобразователь / воспроизводитель звука: временные возможности глубоко эхолического ребенка». Мозг и язык. 4 (3): 396–402. Дои:10.1016 / 0093-934X (77) 90034-7. PMID 907878. S2CID 29492873.
- ^ Шипперс, МБ; Roebroeck, A; Renken, R; Nanetti, L; Кейзерс, С. (2010). «Отображение потока информации от одного мозга к другому во время жестовой коммуникации» (PDF). Proc Natl Acad Sci U S A. 107 (20): 9388–93. Bibcode:2010PNAS..107.9388S. Дои:10.1073 / pnas.1001791107. ЧВК 2889063. PMID 20439736. Архивировано из оригинал (PDF) 27 февраля 2012 г.
- ^ Моро, Андреа (2008). Границы Вавилона. Мозг и загадка невозможных языков. MIT Press. п. 257. ISBN 978-0-262-13498-9.
- ^ Longo, M. R; Кособуд, А .; Бертенталь, Б. И. (апрель 2008 г.). «Автоматическая имитация биомеханически возможных и невозможных действий: эффекты предварительных движений против целей» (PDF). J Exp Psychol Hum Percept Perform. 34 (2): 489–501. Дои:10.1037/0096-1523.34.2.489. PMID 18377184.
- ^ Косоногов, Владимир (2011). «Прослушивание предложений, связанных с действием, ухудшает контроль позы». Журнал электромиографии и кинезиологии. 21 (5): 742–745. Дои:10.1016 / j.jelekin.2011.05.007. ISSN 1050-6411. PMID 21705230.
- ^ Chartrand, T. L .; Барг, Дж. А. (июнь 1999 г.). «Эффект хамелеона: связь между восприятием и поведением и социальное взаимодействие». J Pers Soc Psychol. 76 (6): 893–910. Дои:10.1037/0022-3514.76.6.893. PMID 10402679. S2CID 11818459.
- ^ Lakin, J. L .; Чартран, Т. Л. (июль 2003 г.). «Использование бессознательной поведенческой мимикрии для создания аффилированности и взаимопонимания». Психологическая наука. 14 (4): 334–9. Дои:10.1111/1467-9280.14481. PMID 12807406. S2CID 8458849.
- ^ van Baaren, R. B .; Maddux, W. W .; Chartrand, T. L .; De Bouter, C .; ван Книппенберг, А. (май 2003 г.). «Чтобы имитировать, нужны двое: поведенческие последствия самооценки». Журнал личности и социальной психологии. 84 (5): 1093–102. Дои:10.1037/0022-3514.84.5.1093. PMID 12757151. S2CID 729948.
- ^ Привет, Сесилия (2011). «Автоматическая имитация». Психологический бюллетень. 137 (3): 463–83. Дои:10.1037 / a0022288. PMID 21280938. S2CID 6975248.
- ^ Паукнер А., Суоми С.Дж., Висалберги Э., Ferrari PF (август 2009 г.). «Обезьяны-капуцины демонстрируют привязанность к людям, которые им подражают». Наука. 325 (5942): 880–3. Bibcode:2009Sci ... 325..880P. Дои:10.1126 / science.1176269. ЧВК 2764469. PMID 19679816.
- ^ а б Оберман Л. М., Хаббард Е. М., Макклири Дж. П., Альтшулер Е. Л., Рамачандран В. С., Пинеда Дж. А., ЭЭГ-свидетельства дисфункции зеркальных нейронов при аутистических спектральных расстройствах В архиве 2007-01-03 на Wayback Machine, Brain Res Cogn Brain Res .; 24 (2): 190-8, 2005-06
- ^ Дапретто, М; Дэвис, MS; Pfeifer, JH; Скотт, АА; Сигман, М; Bookheimer, SY; Якобони, М. (2006). «Понимание эмоций в других: дисфункция зеркальных нейронов у детей с расстройствами аутистического спектра». Природа Неврология. 9 (1): 28–30. Дои:10.1038 / nn1611. ЧВК 3713227. PMID 16327784.
- ^ Дапретто М., Дэвис М.С., Пфейфер Дж. Х. и др. (Январь 2006 г.). «Понимание эмоций в других: дисфункция зеркальных нейронов у детей с расстройствами аутистического спектра». Nat. Неврологи. 9 (1): 28–30. Дои:10.1038 / nn1611. ЧВК 3713227. PMID 16327784.
- ^ Перкинс Т., Стоукс М., Макгилливрей Дж., Биттар Р. (октябрь 2010 г.). «Дисфункция зеркального нейрона при расстройствах аутистического спектра». Журнал клинической неврологии. 17 (10): 1239–43. Дои:10.1016 / j.jocn.2010.01.026. PMID 20598548. S2CID 15141982.
- ^ Оберман Л. М., Рамачандран В. С., Пинеда Дж. А. (апрель 2008 г.). «Модуляция подавления мю у детей с расстройствами аутистического спектра в ответ на знакомые или незнакомые стимулы: гипотеза зеркального нейрона». Нейропсихология. 46 (5): 1558–65. Дои:10.1016 / j.neuropsychologia.2008.01.010. PMID 18304590. S2CID 14280719.
- ^ Черкленд, П.С. (2011). «6». Braintrust. Издательство Принстонского университета. п. 156.
- ^ Две системы понимания действий при аутизме: зеркальное отображение и ментализация, Барон-Коэн, Тагер-Флусберг и Ломбардо, глава «Понимание других умов» [2]
- ^ Хаджихани, Н; и другие. (2006). «Анатомические различия в системе зеркального нейрона и сети социального познания при аутизме». Кора головного мозга. 16 (9): 1276–82. Дои:10.1093 / cercor / bhj069. PMID 16306324.
- ^ https://www.newscientist.com/article/dn18837-mirror-neurons-seen-behaving-normally-in-autism.html
- ^ Гамильтон, А. Ф. (август 2009 г.). «Цели, намерения и психические состояния: вызовы теорий аутизма» (PDF). Журнал детской психологии и психиатрии. 50 (8): 881–892. CiteSeerX 10.1.1.621.6275. Дои:10.1111 / j.1469-7610.2009.02098.x. PMID 19508497.
- ^ Мерфи, П; Брэди, N; Фитцджеральд, М; Troje, Н.Ф. (2009). «Нет доказательств нарушения восприятия биологического движения у взрослых с расстройствами аутистического спектра». Нейропсихология. 47 (14): 3225–35. Дои:10.1016 / j.neuropsychologia.2009.07.026. PMID 19666038. S2CID 12495492.
- ^ Сайгин, А .; Cook, J .; Блейкмор, С. (18 октября 2010 г.). Бейкер, Крис I (ред.). «Незатронутые пороги восприятия для биологического и небиологического восприятия формы из движения в условиях аутистического спектра». PLOS One. 5 (10): e13491. Bibcode:2010PLoSO ... 513491S. Дои:10.1371 / journal.pone.0013491. ЧВК 2956672. PMID 20976151.
- ^ Повар, Дженнифер; Сайгин, Айше Пинар; Суэйн, Рэйчел; Блейкмор, Сара-Джейн (2009). «Пониженная чувствительность к биологическому движению с минимальным рывком в условиях аутистического спектра». Нейропсихология. 47 (14): 3275–3278. Дои:10.1016 / j.neuropsychologia.2009.07.010. ЧВК 2779370. PMID 19632248.
- ^ Fan, Y.T .; Decety, J .; Yang, C. Y .; Liu, J. L .; Ченг, Ю. (сентябрь 2010 г.). «Неразрушенные зеркальные нейроны при расстройствах аутистического спектра». Журнал детской психологии и психиатрии. 51 (9): 981–988. Дои:10.1111 / j.1469-7610.2010.02269.x. PMID 20524939.
- ^ https://www.newscientist.com/article/dn18837-mirror-neurons-seen-behaving-normally-in-autism/
- ^ Гордон Р. (1986). «Народная психология как симуляция». Разум и язык. 1 (2): 158–171. Дои:10.1111 / j.1468-0017.1986.tb00324.x.
- ^ Гольдман А. (1989). «Психологизированная интерпретация». Разум и язык. 4 (3): 161–185. Дои:10.1111 / j.1468-0017.1989.tb00249.x.
- ^ Мельцов А.Н., Десети Дж. (Март 2003 г.). «Что подражание говорит нам о социальном познании: сближении психологии развития и когнитивной нейробиологии». Философские труды Лондонского королевского общества. Серия B, Биологические науки. 358 (1431): 491–500. Дои:10.1098 / rstb.2002.1261. ЧВК 1351349. PMID 12689375.
- ^ Соммервилл Дж. А., Десети Дж. (Апрель 2006 г.). «Плетение ткани социального взаимодействия: определение психологии развития и когнитивной нейробиологии в области моторного познания». Психономический бюллетень и обзор. 13 (2): 179–200. Дои:10.3758 / BF03193831. PMID 16892982. S2CID 14689479.
- ^ Кейзерс С., Газзола В. (2007). «Интеграция моделирования и теории разума: от себя к социальному познанию» (PDF). Тенденции в когнитивных науках. 11 (5): 194–196. Дои:10.1016 / j.tics.2007.02.002. PMID 17344090. S2CID 18930071. Архивировано из оригинал (PDF) на 25 марта 2009 г.
- ^ Haroush K, Williams ZM (март 2015 г.). «Нейронное предсказание поведения оппонента при совместном социальном обмене у приматов». Клетка. 160 (6): 1233–45. Дои:10.1016 / j.cell.2015.01.045. ЧВК 4364450. PMID 25728667.
- ^ Cheng YW, Tzeng OJ, Decety J, Imada T, Hsieh JC (июль 2006 г.). «Гендерные различия в системе зеркал человека: исследование магнитоэнцефалографии». NeuroReport. 17 (11): 1115–9. Дои:10.1097 / 01.wnr.0000223393.59328.21. PMID 16837838. S2CID 18811017.
- ^ Cheng Y, Decety J, Lin CP, Hsieh JC, Hung D, Tzeng OJ (июнь 2007 г.). «Половые различия в возбудимости позвоночника при наблюдении за двуногим движением». NeuroReport. 18 (9): 887–90. Дои:10.1097 / WNR.0b013e3280ebb486. PMID 17515795. S2CID 16295878.
- ^ Ян CY, Decety J, Lee S, Chen C, Cheng Y (январь 2009 г.). «Гендерные различия мю-ритма во время сочувствия к боли: электроэнцефалографическое исследование». Исследование мозга. 1251: 176–84. Дои:10.1016 / j.brainres.2008.11.062. PMID 19083993. S2CID 40145972.
- ^ Cheng Y, Lee PL, Yang CY, Lin CP, Hung D, Decety J (май 2008 г.). «Гендерные различия в мю-ритме зеркально-нейронной системы человека». PLOS ONE. 3 (5): e2113. Bibcode:2008PLoSO ... 3.2113C. Дои:10.1371 / journal.pone.0002113. ЧВК 2361218. PMID 18461176.
- ^ Schulte-Rüther, M; Маркович, HJ; Шах, штат Нью-Джерси; Финк, гр .; Пифке, М. (1 августа 2008 г.). «Гендерные различия в мозговых сетях, поддерживающих эмпатию». NeuroImage. 42 (1): 393–403. Дои:10.1016 / j.neuroimage.2008.04.180. PMID 18514546. S2CID 10461927.
- ^ а б c Джалал, Баланд; Рамачандран, Вилаянур С. (2017). «Сонный паралич», «Призрачный нарушитель спальни» и внетелесные переживания: роль зеркальных нейронов ». Границы нейробиологии человека. 11: 92. Дои:10.3389 / fnhum.2017.00092. ISSN 1662-5161. ЧВК 5329044. PMID 28293186.
- ^ Маккормик Л.М., Брамм М.С., Бидл Дж. Н., Парадизо С., Ямада Т., Андреасен Н. (март 2012 г.). «Зеркальная функция нейрона, психоз и эмпатия при шизофрении». Психиатрические исследования. 201 (3): 233–9. Дои:10.1016 / j.pscychresns.2012.01.004. ЧВК 3545445. PMID 22510432.
дальнейшее чтение
- Хикок, Грег (2014). Миф о зеркальных нейронах. ISBN 978-0-393-08961-5.
- Кейзерс, К. (2011). Эмпатический мозг. ISBN 978-1-4637-6906-2.
- Якобони М, Мацциотта Дж.С. (2007). «Зеркальная нейронная система: основные данные и клиническое применение». Энн Нейрол. 62 (3): 213–8. Дои:10.1002 / ana.21198. PMID 17721988. S2CID 3225339.
- Кейзерс, К. и Газзола, В. (2006), На пути к объединяющей нейронной теории социального познания, Прогресс в исследованиях мозга.[3]
- Морселла, Э., Барг, Дж. А., и Голлвитцер, П.М. (Ред.) (2009). Оксфордский справочник по человеческим действиям. Нью-Йорк: Oxford University Press.
- Preston, S.D .; де Ваал, F.B.M. (2002). «Сочувствие: его конечная и ближайшая основы». Поведенческие науки и науки о мозге. 25 (1): 1–72. Дои:10.1017 / s0140525x02000018. PMID 12625087.
- Риццолатти G, Fabbri-Destro M, Cattaneo L (2009). «Зеркальные нейроны и их клиническое значение». Нат Клин Прак Нейрол. 5 (1): 24–34. Дои:10.1038 / ncpneuro0990. PMID 19129788. S2CID 2979216.
- Риццолатти, Г., Синигалья, К. (2008). Зеркала в мозгу. Как мы делимся своими действиями и эмоциями. Оксфорд (Великобритания), Oxford University Press.
внешняя ссылка
- NOVA scienceNOW: зеркальные нейроны (включая 14-минутный эфирный сегмент)
- Ты напоминаешь мне меня в New York Times.
- 7-минутное видео на TED.com :: Нейробиолог Вилаянур Рамачандран описывает функции зеркальных нейронов
- Форум Mirror Neuron, Перспективы психологической науки, сентябрь 2011 г.
- Talking Brains, Грег Хикок и Дэвид Поппель, Новости и взгляды на нейронную организацию языка
- Томас, Бен: Что такого особенного в зеркальных нейронах? Гостевой блог Scientific American, 2012 г. (обзор известных исследовательских подходов, основанный на интервью с Якобони, Хикоком, Хейесом и Галлезе)
- Зеркальные нейроны: все еще открытый вопрос? Паоло Б. Пасколо. Progress in Neuroscience 2013; 1 (1-4): 25-82