Эволюция берёзовой пяденицы - Peppered moth evolution


В эволюция пяденицы является эволюционный случай направленного изменения окраски популяции моли в результате загрязнения воздуха во время Индустриальная революция. Частота бабочек темного цвета в то время увеличилась, пример промышленный меланизм. Позже, когда загрязнение было уменьшено, снова преобладала светлая форма. Промышленный меланизм в моль перечная был ранним испытанием Чарльз Дарвин с естественный отбор в действии и остается классическим примером в учение эволюции.[1][2] В 1978 г. Сьюэлл Райт описал это как «самый ясный случай, когда действительно наблюдался заметный эволюционный процесс».[3][4]
Темный цвет или меланический форма пяденицы перечной (вар. карбонария) не был известен до 1811 г. После полевого сбора в 1848 г. Манчестер, промышленный город в Англии, частота встречаемости этого сорта резко возросла. К концу XIX века он почти полностью превосходил первоначальный светлый шрифт (вар. типика), с рекордом 98% в 1895 г.[5] Об эволюционной важности мотылька размышляли только при жизни Дарвина. Спустя 14 лет после смерти Дарвина, в 1896 году, J.W. Тутт представил это как случай естественного отбора.[6] Благодаря этому идея получила широкое распространение, и все больше людей верили в теорию Дарвина.
Бернард Кеттлуэлл был первым, кто исследовал эволюционный механизм адаптации берёзовой бабочки в период с 1953 по 1956 год. Он обнаружил, что тело светлого цвета является эффективным камуфляж в чистой среде, например, в Дорсет, в то время как темный цвет был полезен в загрязненной среде, например, в Бирмингем. Такое избирательное выживание было связано с птицами, которые легко ловили темных бабочек на чистых деревьях и белых бабочек на деревьях, затемненных копотью. История, поддержанная Кеттлвелл эксперимент, стал каноническим примером Дарвиновский эволюция и свидетельство естественного отбора используется в стандартных учебниках.[7]
Однако неудача в воспроизведении эксперимента и критика методов Кеттлвелла Теодором Дэвидом Сарджентом в конце 1960-х годов привели к общему скептицизму. Когда Джудит Хупер с Мотыльков и людей был опубликован в 2002 году, история Кеттлвелла была подвергнута более суровой критике, обвинена в мошенничестве и широко игнорировалась. Критика стала главным аргументом для креационистов. Майкл Майерус был главным защитником. Его семилетний эксперимент, начатый в 2001 году, был самым сложным в своем роде в популяционная биология, результаты которого были опубликованы посмертно в 2012 году, очень подробно подтвердили работу Кеттлвелла. Это восстановило эволюцию берёзовой бабочки как «самое прямое свидетельство» и «один из самых ярких и легко понимаемых примеров дарвиновской эволюции в действии».[8]
Происхождение и эволюция

Перед Индустриальная революция, моль перечная была редкостью. Первый черный экземпляр (неизвестного происхождения) хранился в Оксфордский университет в 1811 г.[9][10][11] Первый живой экземпляр был пойман Р.С. Эдлстон в Манчестер, Англия в 1848 году, но он сообщил об этом только 16 лет спустя, в 1864 году в журнале Энтомолог.[12] Эдлстон отмечает, что к 1864 году это был наиболее распространенный вид мотылька в его саду в Манчестере. Легкие бабочки смогли слиться со светлыми. лишайники и кору деревьев, а менее распространенная черная моль с большей вероятностью была съедена птицами. Таким образом, из-за обычных светлых лишайников и английских деревьев, светлые бабочки были гораздо более эффективны в укрывании от хищников, а частота темного аллеля составляла около 0,01%.[13]
В первые десятилетия промышленной революции в Англии сельская местность между Лондоном и Манчестером была покрыта сажа от новых угольных заводов. Многие легкие лишайники погибли от Диоксид серы выбросы, и деревья потемнели. Это привело к увеличению охоты птиц на светлые бабочки, так как они больше не смешивались с их загрязненный экосистема: действительно, их тела теперь резко контрастируют с цветом коры. С другой стороны, бабочки темного цвета были замаскированный очень хорошо у почерневших деревьев.[7] Быстро увеличивалась популяция темной бабочки. К середине 19 века количество бабочек темного цвета заметно возросло, и к 1895 году процент темных бабочек в Манчестере составлял 98%, что резко изменилось (почти на 100%) по сравнению с первоначальной частотой. .[7] Этот эффект индустриализации цвета кузова привел к появлению термина "промышленный меланизм ".[2]
Подтверждение того, что промышленный меланизм может быть доказательством Чарльз Дарвин теория естественный отбор был замечен еще при жизни. Альберт Бриджес Фарн (1841–1921), британский энтомолог, 18 ноября 1878 г. написал Дарвину, в котором обсудил свои наблюдения за изменениями окраски моли Annulet (тогда Гнофос обскурата, сейчас же Charissa obscurata ). Он отметил наличие темной бабочки в торфе в New Forest, бурые бабочки на глине и красной почве в Херефордшир, и белые мотыльки на меловых скалах в Льюис, затем предположил, что этот вариант является примером «выживания наиболее приспособленных». Он сказал Дарвину, что нашел темных бабочек на меловом склоне, где листва почернела от дыма от дыма. известковые печи, и он также слышал, что белые мотыльки стали менее распространены в Льюисе после того, как печи для обжига извести проработали несколько лет.[14] Дарвин, похоже, не ответил на эту информацию, возможно, потому, что он думал, что естественный отбор будет намного более медленным процессом.[15] Научное объяснение окраски моли было опубликовано только в 1896 году, через 14 лет после смерти Дарвина, когда J.W. Тутт явно связали меланизм бёдра с естественным отбором.[13]
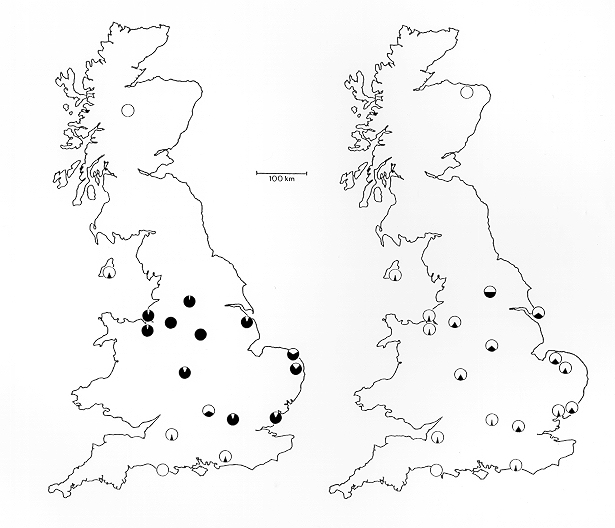
Рост и падение частоты фенотипов
Меланизм появился в популяциях берёзовой пяденицы Европы и Северной Америки. Информации о росте частоты очень мало. Намного больше известно о последующем падении фенотип частота, как было измерено лепидоптерологи с помощью ловушки для моли.
Стюард собрал данные для первых записей перечной мотылька по местности и пришел к выводу, что морфа карбонария была результатом единственного мутация которые впоследствии распространились. К 1895 году он достиг 98% в Манчестере.[16]
Примерно с 1962 г. по настоящее время частота фенотипа карбонария неуклонно приближается к более чистому воздуху в промышленных городах. Его снижение было измерено более точно, чем его рост, посредством более тщательных научных исследований. Примечательно, что Бернард Кеттлуэлл провел общенациональное исследование в 1956 году. Брюс Грант провели аналогичный в начале 1996 г.,[17] и Л.М.Кук в 2003 году.[18]
Подобные результаты были получены в Америке. Формы меланика в Японии не обнаружены. Считается, что это связано с тем, что пяденица в Японии не обитает в промышленно развитых регионах.[18]
Генетика
J.W. Тутт был первым, кто предложил «гипотезу дифференциального хищничества птиц» в 1896 году как механизм естественный отбор. Меланические морфы лучше маскировались на фоне коры деревьев без лиственных лишайников, тогда как типика морфы лучше маскировались на фоне деревьев с лишайниками. В результате птицы будут чаще находить и есть те морфы, которые не были замаскированы.[19]
В 1924 г. J.B.S. Холдейн рассчитывается, используя простой общая модель выбора - селективное преимущество, необходимое для зарегистрированной естественной эволюции пядениц, основанное на предположении, что в 1848 г. частота появления пядениц темной окраски составляла 2%, а к 1895 г. - 95%. Темная или меланическая форма должна была быть на 50% более подходящей, чем типичная светлая форма. Даже с учетом возможных ошибок в модели это разумно исключало стохастический процесс генетического дрейфа, потому что изменения были слишком быстрыми.[20] Статистический анализ Холдейна отбора меланического варианта у берёзовых мотыльков стал хорошо известной частью его усилий по демонстрации того, что математические модели, сочетающие естественный отбор с менделевской генетикой, могут объяснить эволюцию - усилия, которые сыграли ключевую роль в основании дисциплины популяционная генетика, и начало современный синтез эволюционной теории с генетикой.[21]
У пяденицы аллель темной пяденицы доминирующий, а аллель легкой бабочки - рецессивный, что означает, что типика у мотыльков есть фенотип (видимая или обнаруживаемая характеристика), которая видна только в гомозиготный генотип (организм, имеющий две копии одного и того же аллеля), и никогда гетерозиготный один. Это помогает объяснить, насколько быстро изменилось население, когда выбранный для темного окрашивания.[нужна цитата ]
Моль перечная Biston Betularia также модель параллельная эволюция в заболеваемости меланизмом в британской форме (f. карбонария) и американская форма (f. Swettaria), поскольку они неотличимы по внешнему виду. Генетический анализ показывает, что оба фенотипа наследуются как аутосомно-доминантные. Перекрестная гибридизация показывает, что фенотипы продуцируются изоаллелями в одном локусе.[22]
Ген для карбонария в B. betularia считалось, что это район хромосома 17, но позже был сделан вывод, что он не может содержать его, потому что ни один из генов в хромосоме не кодирует ни рисунок крыльев, ни меланинизацию.[23][24] Область, которая использовалась для его обнаружения, была первым интроном ортолога кора ген в Дрозофила. Путем исключения кандидатов в пределах области на основании редкости осталась вставка из 21 925 пар оснований.[24]
Вкладыш с надписью карбюратор-TE, это II класс сменный элемент который имеет неповторяющуюся последовательность размером приблизительно 9 т.п.н., которая повторяется тандемно два и третий раз.[24] В целевом сайте 6 пар оснований инвертированных повторов и 4 дублированных пары оснований, которых нет в типика моль. УглеводыTE имеет более высокую экспрессию на стадии быстрого морфогенеза крыльевых дисков.[24] Механизм того, как он увеличивает экспрессию или является ли он единственным вовлеченным геном, до сих пор неизвестен.[24]
Альтернативные гипотезы
Несколько альтернативных гипотез естественному отбору как движущей силе эволюции были предложены в течение 1920-х и 1930-х годов. Случайная мутация, миграция или генетический дрейф также рассматривались как основные силы эволюции.[25] Однако масштабы наблюдаемых изменений можно объяснить только естественным отбором. Это видно из популяционная генетика что недифференциальное изменение не вызовет эволюции. Если частоты аллелей обозначены алгебраический термины п и q, и скажи) п = 0,6 и q = 0,4, то недифференциальное сокращение численности популяции, скажем, с 2000 до 100 особей, все равно даст те же значения (приблизительно) п = 0,6 и q = 0.4.
П.А. Райли предложил дополнительный фактор отбора, в котором хэви-метал хелатирование с помощью меланина может защитить бёдер от токсического воздействия тяжелых металлов, связанного с индустриализацией. Это селективное преимущество могло бы дополнить основной селективный механизм дифференцированного хищничества птиц.[26]
Фенотипическая индукция
Джон Уильям Хеслоп-Харрисон (1920) отверг гипотезу Тутта о различном хищничестве птиц на том основании, что он не верил в то, что птицы питаются молью. Вместо этого он отстаивал идею о том, что загрязнители могут вызывать изменения в соме и зародышевой плазме организма. Эта гипотеза, вероятно, уходит корнями в 1890-е годы, когда она была предложена как форма Ламаркизм. Важно отметить его исторический контекст.[нужна цитата ]
Хасебрук (1925) был первым, кто попытался доказать эту гипотезу. Он утверждал, что загрязнение воздуха изменило физиологию чешуекрылых, в результате чего образовался избыток черного пигмента. Он подвергал куколок чешуекрылых воздействию различных доз загрязняющих газов, а именно: сероводород (ЧАС2S), аммиак (NH3) и «пыредин» (предположительно, его написание пиридин ). В своих исследованиях он использовал восемь видов, четыре из которых были видами бабочек, не проявлявших меланизма. Форд (1964) утверждает, что иллюстрации Хасебрука показали, что появившиеся аномальные формы не были меланиками, и Хасебрук не смог изучить их генетику.[нужна цитата ]
Хеслоп-Харрисон (Харрисон и Гаррет, 1926; Харрисон, 1928) предположили, что увеличение количества меланиновой моли в промышленно развитых регионах было связано с "давление мутации ", а не отбору хищниками, которых он считал незначительными. Соли вести и марганец присутствовали в частицах загрязняющих веществ, переносимых по воздуху, и он предположил, что они вызывают мутацию генов производства меланина, но не других. Он использовал Selenia bilunaria и Tephrosia bistortata как материал. Личинок кормили листьями, которые содержали эти соли, и впоследствии появились меланики. Подобные эксперименты Хьюза МакКенни (1932) и Томасена и Лемче (1933) не смогли воспроизвести эти результаты. Однако статистик и генетик Рональд Фишер показали, что средства контроля Хеслоп-Харрисон были неадекватными.[27] Эта гипотеза, однако, оказалась опровергнута экспериментами по разведению.
Кеттлвелл эксперимент
Первые важные опыты на бёдровой моли были проведены Бернард Кеттлуэлл в Оксфордском университете под руководством Э. Б. Форд, который помог ему получить грант от Фонд Наффилда для проведения экспериментов. В 1953 году Кеттлвелл начал предварительный эксперимент, в ходе которого бабочки были выпущены в большую (18 м × 6 м) вольер, где их кормили большие сиськи (Парус майор). Его главный эксперимент на Природный заповедник Кэдбери в Бирмингем, Англия, включал маркировку, выпуск и повторный отлов помеченных бабочек. Он обнаружил, что в этом загрязненном лесу типика Предпочтительно охотились на моль. Таким образом, он показал, что меланический фенотип важен для выживания перечной бабочки в такой среде обитания. Кеттлуэлл повторил эксперимент в 1955 году в незагрязненных лесах в Дорсет и снова в грязном лесу Бирмингема. Его сопровождал Нико Тинберген, и они вместе сняли фильм.[2][19] В 1956 году он повторил эксперименты и получил аналогичные результаты; в Бирмингеме птицы съели большую часть белой бабочки (75%), тогда как в Дорсете большая часть темной бабочки (86%) была съедена.[1][28]
Критика
Теодор Дэвид Сарджент проводил эксперименты между 1965 и 1969 годами, из которых он пришел к выводу, что невозможно воспроизвести результаты Кеттлвелла, и сказал, что птицы не отдают предпочтения мотыльку ни на черных, ни на белых стволах деревьев.[29][30] Он предположил, что Кеттлвелл обучил птиц собирать бабочек на стволах деревьев для получения желаемых результатов.[3][31]
Майкл Майерус книга 1998 года Меланизм: эволюция в действии является адаптацией произведений Кеттлвелла Эволюция меланизма, в котором обсуждалась критика оригинальных экспериментальных методов Кеттлвелла.[19] Рецензируя книгу, Джерри Койн пришел к выводу, что «пока мы должны отказаться от Бистон как хорошо понятый пример естественного отбора в действии, хотя, несомненно, это случай эволюции. Есть много исследований, более подходящих для использования в классе ».[32]
Джудит Хупер книга Мотыльков и людей (2002) - одна из самых суровых критик эксперимента Кеттлвелла.[33] Хупер утверждал, что полевые записи Кеттлвелла не могут быть найдены, и предположил, что его эксперимент был мошенничеством, на основе критики Сарджента, утверждающего, что фотографии бабочек были сделаны из мертвых бабочек, помещенных на бревно. Она сказала, что Э. Б. Форд был «дарвиновским фанатиком»,[34] и утверждал, что использовал наивного с научной точки зрения Кеттлвелла для получения желаемых экспериментальных результатов. Затем она заявила, что ученые в целом проявили «легковерное и предвзятое» принятие эволюции.[35] Прием книги привел к утверждениям о том, что историю эволюции берёзовой мотыльки следует исключить из учебников.[36][37] Ученые изучили утверждения Хупера и пришли к выводу, что они необоснованны.[18][38][39]

Филип Э. Джонсон, соучредитель креационистов умный дизайн движение, заявило, что бабочки «не сидят на стволах деревьев», «бабочки должны быть приклеены к стволам» для фотографий и что эксперименты были «мошенническими» и «мошенническими».[40] Сторонник разумного замысла Джонатан Уэллс написал эссе на эту тему, сокращенная версия которого появилась в номере журнала от 24 мая 1999 г. Ученый, утверждая, что «За 25 лет полевых исследований К.А.Кларк и его коллеги нашли только одну пяденицу на стволе дерева», и пришли к выводу, что «Тот факт, что пяденица обыкновенная обычно не отдыхает на стволах деревьев, опровергает эксперименты Кеттлвелла».[41] Далее Уэллс писал в 2000 г. Иконы эволюции, в котором он утверждает: «Однако учебники не объясняют того, что биологи с 1980-х годов знали, что у классической истории есть некоторые серьезные недостатки. Самый серьезный из них заключается в том, что бородавчатая моль в дикой природе даже не отдыхает на ней. стволы деревьев. Учебные фотографии, оказывается, инсценированы ».[42] Однако пяденицы иногда отдыхают на стволах деревьев, и разница между «постановочными» и «постановочными» фотографиями невелика.[43]
Эксперимент Майеруса
С 2001 по 2007 год Майерус проводил эксперименты в Кембридже, чтобы разрешить различные обоснованные критические замечания по поводу эксперимента Кеттлвелла. Во время своего эксперимента он отметил естественное положение покоя перечной бабочки. Из 135 исследованных мотыльков более половины находились на ветвях деревьев, в основном на нижней половине ветви, 37% - на стволах деревьев, в основном на северной стороне, и только 12,6% отдыхали на ветках или под ними. После переписки с Хупером он добавил эксперимент, чтобы выяснить, летучие мыши, а не птицы, могли быть главными хищниками. Он наблюдал, как несколько видов птиц фактически охотятся на бабочек, и это различное хищничество птиц было основным фактором, ответственным за сокращение численности бабочек. карбонария частота по сравнению с типика.[19] Он описал свои результаты как полное подтверждение теории естественного отбора эволюции берёзовой бабочки и сказал: «Если взлет и падение берёзовой бабочки является одним из наиболее впечатляющих и легко понимаемых примеров дарвиновской эволюции в действии, это должно быть учил. В конце концов, он предоставляет доказательства эволюции ".[44]
Майерус умер до того, как успел завершить запись своих экспериментов, поэтому работа была продолжена Куком, Грантом, Саккери и Маллетом и опубликована 8 февраля 2012 года как «Избирательное истребление птичьей мотылькой: последний эксперимент Майкла Майеруса. . "[45] Эксперимент стал крупнейшим за всю историю изучения промышленного меланизма, в котором участвовало 4864 человека за шестилетнее исследование, и он подтвердил, что меланизм у бабочек является подлинным примером естественного отбора, включающего камуфляж и хищничество. Их заключительное замечание гласит: «Эти данные предоставляют самые прямые доказательства того, что маскировка и хищничество птиц являются основным объяснением роста и падения меланизма у бабочек».[8]
Койн ответил: «Несмотря на оборонительную позицию британских эволюционистов, я думаю, что моя критика имела определенный вес, потому что кембриджский биолог Майкл Майерус решил повторить эксперименты Кеттлвелла, но на этот раз сделал их правильно». Он процитировал Кука и другие. вывод о том, что «Эти новые данные отвечают на критику более ранней работы и подтверждают методологию, применявшуюся во многих предыдущих экспериментах с хищниками, в которых стволы деревьев использовались в качестве мест отдыха. Новые данные в сочетании с весомостью ранее существовавших данных убедительно показывают, что« промышленный меланизм в засеянных перцем » мотылек по-прежнему остается одним из наиболее ярких и понятных примеров дарвиновской эволюции в действии ». Койн сказал, что он «рад согласиться с этим выводом, который отвечает на мои предыдущие критические замечания по поводу Бистон сказка."[46]
Смотрите также
использованная литература
- ^ а б Радж, Дэвид В. (2005). "Красота классической экспериментальной демонстрации естественного отбора Кеттлуэлла". Бионаука. 55 (4): 369–375. Дои:10.1641 / 0006-3568 (2005) 055 [0369: TBOKCE] 2.0.CO; 2.
- ^ а б c Майерус, Майкл Э. Н. (2008). «Промышленный меланизм у берёзовой мотылька, Biston betularia: прекрасный обучающий пример дарвиновской эволюции в действии» (PDF). Эволюция: образование и пропаганда. 2 (1): 63–74. Дои:10.1007 / s12052-008-0107-y. S2CID 25407417.
- ^ а б Райс, Стэнли А. (2007). Энциклопедия эволюции. Нью-Йорк: факты в файле. п. 308. ISBN 978-1-4381-1005-9.
- ^ М. Э. Н. Майерус (1998). Меланизм: эволюция в действии. Издательство Оксфордского университета. ISBN 978-0-19-854983-3.
- ^ Clarke, C.A .; Mani, G. S .; Винн, Г. (1985). «Эволюция наоборот: чистый воздух и моль». Биологический журнал Линнеевского общества. 26 (2): 189–199. Дои:10.1111 / j.1095-8312.1985.tb01555.x.
- ^ Майерус, Майкл Э. Н. (2008). «Промышленный меланизм у берёзовой мотылька, Biston Betularia: Превосходный обучающий пример дарвиновской эволюции в действии ». Эволюция: образование и пропаганда. 2 (1): 63–74. Дои:10.1007 / s12052-008-0107-y.
- ^ а б c Миллер, Кен (1999). Мотылек: обновление
- ^ а б Cook, L.M .; Grant, B.S .; Saccheri, I.J .; Маллет, Джеймс (2012). «Избирательное истребление пернатой мотылька птицами: последний эксперимент Майкла Майеруса». Письма о биологии. 8 (4): 609–612. Дои:10.1098 / рсбл.2011.1136. ЧВК 3391436. PMID 22319093.
- ^ Берри, Р. Дж. (1990). «Промышленный меланизм и моль (Biston betularia (L.))». Биологический журнал Линнеевского общества. 39 (4): 301–322. Дои:10.1111 / j.1095-8312.1990.tb00518.x.
- ^ Saccheri, I.J .; Rousset, F .; Watts, P.C .; Брейкфилд, П. М .; Кук, Л. М. (2008). «Отбор и поток генов на убывающей стайке меланической пяденицы». Труды Национальной академии наук. 105 (42): 16212–16217. Bibcode:2008ПНАС..10516212С. Дои:10.1073 / pnas.0803785105. ЧВК 2571026. PMID 18854412.
- ^ Нил, Дик (2004). Введение в популяционную биологию (Перепечатка ред.). Кембридж: Издательство Кембриджского университета. п. 171. ISBN 9780521532235.
- ^ Эдлстон, Р. С. (1864). "[Без названия]". Энтомолог. 2: 150. Cite использует общий заголовок (Помогите)
- ^ а б Харт, Адам Дж .; Стаффорд, Ричард; Smith, Angela L .; Гуденаф, Энн Э. (2010). «Доказательства современной эволюции при жизни Дарвина» (PDF). Текущая биология. 20 (3): R95. Дои:10.1016 / j.cub.2009.12.010. PMID 20144776. S2CID 31093691.
- ^ Фарн, А. Б. (18 ноября 1878 г.). "Фарн, А.Б. Дарвину К.Р., Письмо 11747 о корреспондентском проекте Дарвина". Документы Дарвина. Комната рукописей, библиотека Кембриджского университета, Вест-роуд, Кембридж, Англия. ДАР 164: 26.
- ^ «Как необычное письмо Дарвину обнаружило промышленный меланизм в ночных мотыльках». Научный фокус. 27 февраля 2018 г.. Получено 16 июн 2018.
- ^ Стюард, Р. К. (1977). «Промышленный и непромышленный меланизм у берёзовой мотылька. Biston Betularia (Л.) ". Экологическая энтомология. 2 (3): 231–243. Дои:10.1111 / j.1365-2311.1977.tb00886.x. S2CID 85624115.
- ^ http://www.talkorigins.org/faqs/wells/images/grantfile.jpg
- ^ а б c Кук, Л. М. (2003). «Взлет и падение карбонарийной формы перечной бабочки». Ежеквартальный обзор биологии. 78 (4): 399–417. Дои:10.1086/378925. PMID 14737825. S2CID 26831926.
- ^ а б c d Майерус, Майкл Э. Н. (Август 2007 г.). "Мотылек: доказательство дарвиновской эволюции" (PDF). Архивировано из оригинал (PDF) 15 июня 2011 г.. Получено 9 сентября 2007.
- ^ Холдейн, J.B.S. (1924). Математическая теория естественного и искусственного отбора.
- ^ Боулер, Питер Дж. (2003). Эволюция: история идеи, 3-е издание. Калифорнийский университет Press. стр.331–332. ISBN 0-520-23693-9.
- ^ Грант, Б. С. (2004). «Аллельный меланизм у американских и британских бёдер». Журнал наследственности. 95 (2): 97–102. Дои:10.1093 / jhered / esh022. PMID 15073224.
- ^ ван'т Хоф, Арьен Э .; Эдмондс, Никола; Даликова, Мартина; Марек, Франтишек; Саккери, Илик Дж. (20 мая 2011 г.). «Промышленный меланизм у британских пернатых мотыльков имеет единственное и недавнее мутационное происхождение». Наука. 332 (6032): 958–960. Bibcode:2011Науки ... 332..958В. Дои:10.1126 / science.1203043. PMID 21493823. S2CID 24400858 - через JSTOR.
- ^ а б c d е ван'т Хоф, Арьен Э .; Кампань, Паскаль; Ригден, Дэниел Дж .; Юнг, Карл Дж .; Лингли, Джессика; Перепел, Майкл А .; Холл, Нил; Дарби, Алистер; Саккери, Илик Дж. (2 июня 2016 г.). «Мутация промышленного меланизма у британской пяденицы - переносимый элемент». Природа. 534 (7605): 102–117. Bibcode:2016Натура.534..102H. Дои:10.1038 / природа17951. PMID 27251284.
- ^ Добжанский, Т. (1937). Генетика и эволюционный процесс. Издательство Колумбийского университета. ISBN 0-231-08306-8.
- ^ Райли, П.А. (2013). «Предлагаемый селективный механизм, основанный на хелатировании металлов в промышленных меланических мотыльках» (PDF). Биологический журнал Линнеевского общества. 109 (2): 298–301. Дои:10.1111 / bij.12062.
- ^ Фишер, Р.А. (1933). «О доказательствах против химической индукции меланизма у чешуекрылых». Труды Королевского общества B. 112 (778): 407–416. Bibcode:1933RSPSB.112..407F. Дои:10.1098 / rspb.1933.0018.
- ^ Кеттлвелл, Г. Б. Д (1958). "Обзор частот Biston betularia (L.) (Lep.) И его меланических форм в Великобритании". Наследственность. 12 (1): 51–72. Дои:10.1038 / hdy.1958.4.
- ^ Сарджент, Т. Д. (1968). «Загадочная моль: влияние на выбор фона окраски окологлазных чешуек». Наука. 159 (3810): 100–101. Bibcode:1968Sci ... 159..100S. Дои:10.1126 / science.159.3810.100. PMID 5634373. S2CID 32124765.
- ^ Сарджент, Т. Д. (1969). «Выбор фона бледных и меланических форм скрытой моли, Phigalia titea (Cramer)». Природа. 222 (5193): 585–586. Bibcode:1969Натура.222..585С. Дои:10.1038 / 222585b0. S2CID 4202131.
- ^ Сарджент, Т.Д .; Millar, C.D .; Ламберт, Д. (1988). «Глава 9:« Классическое »объяснение промышленного меланизма: оценка свидетельств». In Hecht, Max K .; Уоллес, Брюс (ред.). Эволюционная биология. 23. Пленум Пресс. ISBN 0306429772.
- ^ Койн, Джерри А. (1998). «Не черно-белое. Обзор Меланизм: эволюция в действии Майкла Э. Majerus ". Природа. 396 (6706): 35–36. Дои:10.1038/23856.
- ^ Кенни, Майкл (22 октября 2002 г.). «О темных мотыльках, людях и эволюции». Чикаго Трибьюн. Получено 10 декабря 2014.
- ^ "О мотыльках и людях". W. W. Norton & Company. Архивировано из оригинал 10 декабря 2014 г.. Получено 10 декабря 2014.
- ^ Смит, Питер Д. (11 мая 2002 г.). «О мотыльках и людях: интриги, трагедия и мотылек». Хранитель. Получено 10 декабря 2014.
- ^ "Мотыльков и человеков". Независимый. 4 сентября 2003 г.. Получено 10 декабря 2014.
- ^ Довер, Габби (2003). "Охотники на мотыльков". Отчеты EMBO. 4 (3): 235. Дои:10.1038 / sj.embor.embor778. ЧВК 1315906.
- ^ Грант, Б.С. (2002), «Кислый виноград гнева», Наука, 297: 940–941, Дои:10.1126 / science.1073593, S2CID 161367302
- ^ Майерус, Майкл Э. Н. (2005). «Мотылек: закат ученика Дарвина». В Fellowes, Марк; Холлоуэй, Грэм; Рольф, Йенс (ред.). Эволюционная экология насекомых. Уоллингфорд, Оксон: CABI Publishing. С. 375–377. ISBN 978-1-84593-140-7.
- ^ Фрак, Дональд (16 апреля 1999 г.). «Эволюция - апрель 1999 г .: мотыльки и креационисты». Архивировано из оригинал 26 августа 2007 г.. Получено 26 августа 2007.
- ^ Уэллс, Дж. (24 мая 1999 г.). «Размышления о берёзовых мотыльках; эту классическую историю эволюции путём естественного отбора необходимо пересмотреть». Ученый. 13 (11): 13.
- ^ Уэллс Дж. (2000). Иконы эволюции: Наука или миф? Почему многое из того, что мы говорим об эволюции, неверно. Regnery Press, Вашингтон, округ Колумбия, стр. 138 (книга доступна по адресу Iconsofevolution.com )
- ^ "Икона затемнения: Глава 7: Мотыльки пернатые".
- ^ Коннор, Стив (25 августа 2007 г.). «Исследование мотыльков подтверждает классический« контрольный пример »теории Дарвина». Независимый. Архивировано из оригинал 7 октября 2008 г.. Получено 9 сентября 2007.
- ^ Мацке, Ник (8 февраля 2012 г.). «Избирательное истребление пернатой мотылька птицами: последний эксперимент Майкла Майеруса». Большой палец панды. Получено 7 марта 2012.
- ^ Койн, Джерри (12 февраля 2012 г.). «История бёдра солидная». Почему эволюция верна. Получено 7 марта 2012.
{kind=link}
внешние ссылки
- Брюс Грант написал несколько работ о меланизме у перечной мотылька, которые перечислены на его домашняя страница.
- Онлайн-лекция: "Взлет и падение меланической пяденицы" представленный Лоуренсом Куком.
- Янг, Мэтт. «Самогон: почему мотылек остается символом эволюции». Департамент физики Колорадской горной школы. Получено 3 января 2009.
- Мотылек: Упадок дарвиновского ученика. Это стенограмма Майкл Майерус 'лекция прочитана Британская гуманистическая ассоциация на День Дарвина 2004.
- Мотылек: доказательство дарвиновской эволюции. Это стенограмма лекции Майеруса, прочитанной на Европейское общество эволюционной биологии митинг 23 августа 2007 г. силовая установка презентация так же доступно.
- Майерус, Майкл Э. Н. (2009). «Промышленный меланизм у берёзовой мотылька, Biston Betularia: Превосходный обучающий пример дарвиновской эволюции в действии ». Эволюция: образование и пропаганда. 2 (1): 63–74. Дои:10.1007 / s12052-008-0107-y.
Обвинения в фальсификации данных и научном мошенничестве в этом деле оказались бессмысленными.
- 19 июня 2009 г. Telegraph.co.uk опубликовано статья об этом эволюционном феномене и призвала читателей из Великобритании посетить Сайт подсчета бабочек и записывать свои наблюдения за местными мотыльками, чтобы помочь исследователям увеличить объем доступных данных.