Фитоглобин - Phytoglobin
Похоже, что один из основных авторов этой статьи тесная связь со своим предметом. (Апрель 2017 г.) (Узнайте, как и когда удалить этот шаблон сообщения) |
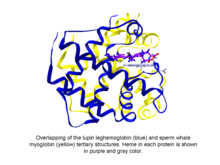
Фитоглобины белки глобулярных растений (водорослей и наземных растений), классифицируемые в глобин суперсемейство, содержащее гем, я.е. протопорфирин IX -Fe, протезная группа. Самые простые известные фитоглобины - это леггемоглобины, обнаруженный в 1939 году Кубо после спектроскопического и химического анализа красного пигмента сои. корневые узелки.[1] Спустя несколько десятилетий после доклада Кубо кристаллизация фитоглобина люпина (известного как леггемоглобин) Вайнштейном и соавторами показала, что третичная структура этого белка и структуры кашалота миоглобин был удивительно похож, что указывало на то, что фитоглобин, открытый Кубо, действительно соответствовал глобину.[2]
Одной из важных функций фитоглобина является его диоксигеназа оксида азота Мероприятия.[3]
Распространение и классификация
Фитоглобины (сокращенно Phytogbs) повсеместно распространены в растениях, поскольку они были идентифицированы в водорослях и наземных растениях, включая примитивные растения. мохообразные и развился однодольные и двудольные. Их можно классифицировать следующим образом:[3][4]
- 2/2 Фитоглобины (семейство усеченных глобинов, подсемейство TrHb2)
- Phytogb3: содержится в водорослях и наземных растениях.
- 3/3 Фитоглобины (миоглобиноподобное семейство)
- Phytogb0: базальный тип, обнаруженный в водорослях, мохообразные и голосеменные.[4]
- Phytogb1, 2: встречается только в покрытосеменные.
Небобовые симбиотические глобины (SymPhytogbs)[4] разбросаны по Phytogb1 и Phytogb2.[3] Симбиотические глобины обычно снабжают кислородом симбиотические бактерии, которые связывают азот. В бобовых культурах ризобия, но у некоторых актиноризных растений актиномицет Frankia вместо этого сделай работу.[3]
Структурные характеристики
Фитогбы кодируются генами, прерванными на 3 интроны (хотя 4 интрона, содержащие фитогб ген был обнаружен в мох Physcomitrella patens ).[5] Первый и третий интрон фитогб гены локализуются в том же положении, что и миоглобин гены, что предполагает, что фитогб и миоглобин гены произошли от общего предка более 600 миллионов лет назад.[6] Наличие второго интрона у фитогб гены были предсказаны Go с помощью теоретического анализа,[7] что было дополнительно подтверждено клонированием и секвенированием соя фунт гены Маркером и соавторами.[8]

Фитогб представляет собой мономерные белки с молекулярной массой от ~ 17 до ~ 19 кДа. Однако при высоких (более 1 мМ) концентрациях фитогб может образовывать димеры. Полипептидная цепь Phytogbs складывается в определенное расположение от 6 до 7 спиралей (обозначенных буквами от A до H), известное как глобиновая складка, которая образует гидрофобный карман, в котором расположен гем. У Phytogbs были идентифицированы два типа глобиновой складки: 3/3 и 2/2 складки,[9][10] где спирали A, E и F перекрываются со спиралями B, G и H, а спирали B и E перекрываются со спиралями G и H соответственно.

Подобно другим глобинам, гем-Fe в Phytogbs координируется в проксимальной области с помощью аминокислоты His (называемой проксимальной His). Дистальная область гем-Fe может быть занята множеством лигандов (таких как кислород и оксид азота ) или дистальная (часто His) аминокислота, уступая место пента- или гексакоординированным Phytogbs соответственно. Phytogbs2, SymPhytogbs и Lbs преимущественно пента-координаты, тогда как Phytogbs1 преимущественно гексакоординаты, а Phytogbs0 и Phytogbs3 представляют собой комбинацию пента- и гексакоординат.[4] Координация гем-Fe важна для функции Phytogb (и других глобинов), поскольку она регулирует скорость связывания лиганда и высвобождения как следствие кинетических констант. kна и kвыключенсоответственно. Например, сродство Lb сои и Phytogb1 риса к O2 (KО2) средний и очень высокий, потому что kна составляет 130 и 68 мМ−1 s−1, kвыключен составляет 5,6 и 0,038 с−1 и KО2 (я.е. О2-родство в результате kна/kвыключен) составляет 23 и 1800 мМ−1соответственно.[11] Это указывает на то, что Lb сои может функционировать как O2-storage или -transport protein, и что функция Phytogb1 риса (и других гексакоординированных Phytogb) может быть отличной от O2-транспорт, потому что высокое сродство этого белка к O2 результат очень низкого kвыключен постоянный.[12]
Синтез и постулируемые функции
Как и другие глобины, пентакоординированные фитогбы обратимо связывают и транспортируют O2. Функция Lbs в узелках была выяснена в 1974 году Виттенбергом, Эпплби и другими.[13] В клубеньках концентрация Lbs очень высока, поскольку они составляют ~ 30% от общего количества растворимых белков. Очевидная функция Lbs в узелках - способствовать диффузии O2 дышащим бактероидам для азотфиксация. В то же время Lb способствует поддержанию низкого уровня O2-уровни (~ 10 нМ), чтобы избежать инактивации O2-чувствительный нитрогеназа фиксирующий атмосферный азот.[14]
Кроме того, Phytogbs связывают другие газообразные лиганды, в первую очередь оксид азота (NO), и проявляют NO диоксигеназа Мероприятия.[15] Работа Хилла с соавторами в течение последних 15 лет показала, что уровни эндогенного NO варьируются в зависимости от концентрации Phytogbs1 в трансгенной кукурузе и люцерне.[16] Основываясь на этих наблюдениях, эти авторы предположили, что функция насыщенных кислородом Phytogbs заключается в модулировании уровней NO посредством активности NO-диоксигеназы и в косвенном регулировании широкого спектра функций клетки, которые модулируются уровнями NO. Окисленные фитоглобины класса 1, реагирующие с NO с образованием нитрата, представляют собой основной механизм, с помощью которого NO улавливается растениями. Цикл, включающий нитратредуктазу, восстановление нитрита до NO, улавливание NO фитоглобином, был определен как цикл фитоглобин-NO[17]. Его работа приводит к поддержанию окислительно-восстановительного и энергетического статуса во время гипоксии и приводит к уменьшению производства этанола и молочной кислоты.[18]
Phytogbs0, 1, 2 и 3 синтезируются в очень низких концентрациях в различных (эмбриональных и вегетативных) органах растений.[19][20][21] Тем не менее, концентрация Phytogbs увеличивается в растениях, подверженных определенным стрессовым условиям, таким как наводнение.[22] и ограничение света.[23] Следовательно, некоторые фитогены считаются чувствительными к стрессу белками растений.
Эволюция
Части этой статьи (те, что связаны с филогенезом Беканы, отвергают общее происхождение Lbs и SymPhytogbs в Phytogb1) должны быть обновлено. (Ноябрь 2020) |
Работа Арредондо-Питера, Виноградова и соавторов прояснила основные события, которые могли произойти во время эволюции Phytogbs.[24][25][26] Хотя фитогбы произошли от общего предка до происхождения эукариоты (я.е. в эубактерии домен), у растений были идентифицированы две эволюционные линии Phytogbs: линия (3 / 3-) Phytogb0, 1 и 2 и линия SymPhytogb и Lb и линия (2 / 2-) Phytogb3. По-видимому, Phytogbs0 были предками Phytogbs1 и 2, а SymPhytogbs и Lbs произошли от предка Phytogb1 как приспособление к симбиотической азотфиксации. Сравнительный анализ мха Phytogb0 со структурой Phytogb1 риса и Lb сои показал, что основные эволюционные изменения, которые, вероятно, произошли во время эволюции Phytogbs0, 1 и 2, SymPhytogbs и Lbs, были (i) переходом гекса-координаты в пента-координату в геме. группы, (ii) уменьшение длины на CD-петле и N- и C-концах, и (iii) уплотнение белка в глобулярную структуру. Более того, структура глобинового домена Phytogbs3 очень похожа на структуру бактериальных усеченных глобинов, что указывает на то, что 2/2-складывание сохраняется во время эволюции Phytogbs3. Однако глобиновый домен наземного растения Phytogbs3 фланкирован внеаминокислотными последовательностями, которые, вероятно, произошли от предка наземного растения Phytogbs3. Функциональное и эволюционное значение внеаминокислотных последовательностей в структуре Phytogbs3 до сих пор не известно.
Технологические приложения
Фитогбс потенциально полезны для ряда биотехнологических применений. Например, вскоре после открытия риса Phytogbs1 предполагаемым применением было использование этих белков в качестве О2-сенсоры в электронных устройствах. Кроме того, чрезмерная экспрессия Phytogbs была предложена в качестве стратегии повышения устойчивости сельскохозяйственных культур к определенным стрессовым условиям, таким как затопление сельскохозяйственных культур. [27] [28], поскольку он связывает ранний сигнал наводнения этилен к низкий кислород реакция растений [29][30]. В последнее время Phytogbs рассматривались кандидатами на разработку кровезаменители и как добавки в вегетарианские бургеры.
Заметки
использованная литература
- ^ Кубо Х., Uber hamoprotein aus den wurzelknollchen von leguminosen, Acta Phytochim. (Токио), 11 (1939) 195-200.
- ^ Вайнштейн Б. К., Арутюнян Э. Х., Куранова И. П., Борисов В. В., Сосфенов Н. И., Павловский А. Г., Гребенко А. И., Конарева Н. В., Структура леггемоглобина из клубеньков корня люпина в разрешении 5 А., Природа, 254 (1975) 163-164
- ^ а б c d е ж Бекана, Мануэль; Юруэла, Инмакулада; Сарат, Гаутама; Каталон, Пилар; Харгроув, Марк С. (сентябрь 2020 г.). «Гемоглобины растений: путь от одноклеточных зеленых водорослей к сосудистым растениям». Новый Фитолог. 227 (6): 1618–1635. Дои:10.1111 / nph.16444.
- ^ а б c d Хилл Р., Харгроув М. С., Арредондо-Питер Р. (2016). «Фитоглобин: новая номенклатура глобинов растений, принятая глобиновым сообществом на XVIII конференции 2014 года по связыванию кислорода и чувствительным белкам». F1000 Исследования. 5: 212. Дои:10.12688 / f1000research.8133.1.CS1 maint: использует параметр авторов (ссылка на сайт)
- ^ Мартинес-Окампо Ф., Васкес-Лимон К., Арредондо-Петер Р., Обнаружение, ПЦР-амплификация и характеристика несимбиотического гена гемоглобина, содержащего 4 интрона, из мха Physcomitrella patens., Global J. Biochem., 3 (2012) 12
- ^ Хардисон Р. К. Краткая история гемоглобинов: растений, животных, протистов и бактерий., Proc. Natl. Акад. Sci. USA., 93 (1996) 5675-5679
- ^ Го М., Корреляция экзонных областей ДНК со структурными единицами белка в гемоглобине., Nature, 291 (1981) 90-92.
- ^ Хилдиг-Нильсен Дж. Дж., Дженсен Э., Палудан К., Виборг О., Гаррет Р., Йоргерсен П. Л., Маркер К. А., Первичные структуры двух генов леггемоглобина сои, Nucleic Acids Res., 10 (1982) 689-701.
- ^ Харгроув М., Брукер Э. А., Стек Б., Сарат Г., Арредондо-Питер Р., Клукас Р. В., Олсон Дж. С., Филипс-младший. Г. Н., Кристаллическая структура несимбиотического гемоглобина., Структура, 8 (2000) 1005-1014.
- ^ Ридер Б. Дж., Хаф М. А., Структура несимбиотического растительного гемоглобина класса 3 из Arabidopsis thaliana раскрывает новое N-концевое удлинение спирали., Acta Crystallogr., D70 (2014) 1411-1418.
- ^ Арредондо-Питер Р., Харгроув М. С., Моран Дж. Ф., Сарат Г., Клукас Р. В., Гемоглобины растений, Физиология растений, 118 (1998) 1121-1126.
- ^ Арредондо-Питер Р., Харгроув М. С., Сарат Г., Моран Дж. Ф., Лорман Дж., Олсон Дж. С., Клукас Р. В., Гемоглобины риса: клонирование генов, анализ и кинетика связывания кислорода рекомбинантного белка, синтезированного в кишечная палочка., Plant Physiol., 115 (1997) 1259-1266.
- ^ Виттенберг Дж. Б., Бергерсен Ф. Дж., Эпплби К. А., Тернер Г. Л., Облегченная диффузия кислорода: роль леггемоглобина в фиксации азота бактероидами, выделенными из клубеньков корня сои., J. Biol. Chem., 249 (1974) 4057-4066
- ^ Эпплби К. А., Леггемоглобин и Ризобий дыхание., Annu. Rev. Plant Physiol., 35 (1984) 443-478.
- ^ Смагх Б. Дж., Трент Дж. Т., Харгроув М. С. Активность диоксигеназы NO в гемоглобинах является повсеместной in vitro, но ограничивается сокращением in vivo., PLoS One, 3 (2008) DOI: 10.1371 / journal.pone.0002039.
- ^ Хилл Р. Д., Несимбиотические гемоглобины - Что происходит помимо поглощения оксида азота ?, AoB Plants, Pls004 (2012) doi: 10.1093 / aobpla / pls1004
- ^ Игамбердиев, А.У .; Барон, К .; Manac'h-Little, N .; Стойменова, М .; Хилл, Р. Д. (2005). «Цикл гемоглобина / оксида азота: участие в стрессе наводнения и влияние на передачу сигналов гормонов». Анналы ботаники. 96 (4): 557–564. Дои:10.1093 / aob / mci210. ISSN 0305-7364. ЧВК 4247025. PMID 16027133.
- ^ Игамбердиев, А.У .; Хилл, Р. Д. (2004). «Нитраты, NO и гемоглобин в адаптации растений к гипоксии: альтернатива классическим путям ферментации». Журнал экспериментальной ботаники. 55 (408): 2473–2482. Дои:10.1093 / jxb / erh272. ISSN 0022-0957. PMID 15448180.
- ^ Гаррочо-Виллегас В., Арредондо-Петер Р., Молекулярное клонирование и характеристика мха (Ceratodon purpureus) несимбиотический гемоглобин дает представление о ранней эволюции несимбиотических гемоглобинов растений., Mol. Биол. Evol., 25 (2008) 1482-1487.
- ^ Росс Э.Дж., Шеарман Л., Матизен М., Чжоу Дж., Арредондо-Питер Р., Сарат Г., Клукас Р.В., Несимбиотические гемоглобины синтезируются во время прорастания и в дифференцирующихся типах клеток., Protoplasma, 218 (2001) 125 -133.
- ^ Уоттс Р. А., Хант П. В., Хвитвед А. Н., Харгроув М. С., Пикок В. Дж., Деннис Э. С., Гемоглобин растений, гомологичных усеченным гемоглобинам микроорганизмов., Proc. Natl. Акад. Sci. USA., 98 (2001) 10119-10124.
- ^ Тейлор Э. Р., Ни Х. З., МакГрегор А. В., Хилл Р. Д. Ген гемоглобина злаков экспрессируется в тканях семян и корней в анаэробных условиях, Plant Mol. Биол., 24 (1994) 853-862
- ^ Лира-Руан В., Сарат Г., Клукас Р. В., Арредондо-Питер Р., Синтез гемоглобинов в рисе (Oryza sativa var. Jackson) растения, растущие в нормальных и стрессовых условиях., Plant Sci., 161 (2001) 279-287.
- ^ Васкес-Лимон К., Хугевис Д., Виноградов С. Н., Арредондо-Петер Р., Эволюция гемоглобинов наземных растений, Plant Sci., 191-192 (2012) 71-81.
- ^ Виноградов С. Н., Фернандес И., Хугевийс Д., Арредондо-Петер Р. Филогенетические взаимоотношения гемоглобинов 3/3 и 2/2 в геномах Archaeplastida с гемоглобинами бактерий и других эукариот, Мол. Завод, 4 (2011) 42-58.
- ^ Виноградов С. Н., Хугевийс Д., Арредондо-Петер Р., Каковы происхождение и филогения гемоглобинов растений ?, Commun. Интегрировать. Биол., 4 (2011) 443-445.
- ^ Анджейчак, О.А.; Havelund, JF; Ван, WQ; Ковальчук, С; Хагенсен, CE; Hasler-Sheetal, H; Дженсен, Онтарио; Rogowska-Wrzesinska, A; Møller, IM; Hebelstrup, KH (24 февраля 2020 г.). «Гипоксический протеом и метаболом ячменя (Hordeum vulgare L.) с добавлением фитоглобина и без него ». Международный журнал молекулярных наук. 21 (4): 1546. Дои:10.3390 / ijms21041546. PMID 32102473.
- ^ Мира, Мохамед М .; Хилл, Роберт Д.; Стасолла, Клаудио (ноябрь 2016 г.). «Фитоглобины улучшают гипоксический рост корней, уменьшая гибель клеток апикальной меристемы». Физиология растений. 172 (3): 2044–2056. Дои:10.1104 / стр. 16.01150. ЧВК 5100795. PMID 27702845.
- ^ Хартман, Иоганнес Герард Вильгельм (22 января 2020 г.). Сигнал раннего затопления этиленом позволяет растениям выдерживать кислородный стресс. Universiteit Utrecht. HDL:1874/387763. ISBN 978-90-393-7231-9.
- ^ Хартман, S; Лю, Z; van Veen, H; Vicente, J; Reinen, E; Martopawiro, S; Чжан, Х; ван Донген, N; Босман, Ф; Бассель, GW; Виссер, EJW; Бейли-Серрес, Дж; Теодулу, Флорида; Hebelstrup, KH; Гиббс, диджей; Холдсворт, MJ; Sasidharan, R; Voesenek, LACJ (5 сентября 2019 г.). «Этилен-опосредованное истощение оксида азота предварительно адаптирует растения к стрессу гипоксии». Nature Communications. 10 (1): 4020. Bibcode:2019НатКо..10.4020H. Дои:10.1038 / s41467-019-12045-4. ЧВК 6728379. PMID 31488841.