Несинаптическая пластичность - Nonsynaptic plasticity

Несинаптическая пластичность это форма нейропластичность это предполагает изменение ионный канал функция в аксон, дендриты, и тело клетки, что приводит к специфическим изменениям в интеграции возбуждающие постсинаптические потенциалы (EPSP) и тормозящие постсинаптические потенциалы (IPSP). Несинаптическая пластичность - это модификация внутренней возбудимость нейрона. Он взаимодействует с синаптическая пластичность, но это считается отдельной сущностью от синаптической пластичности. Собственная модификация электрических свойств нейроны играет роль во многих аспектах пластичности из гомеостатическая пластичность к учусь и объем памяти сам. Несинаптическая пластичность влияет синаптическая интеграция, подпороговый распространение, генерация шипа и другие фундаментальные механизмы нейронов на клеточном уровне. Эти индивидуальные нейрональные изменения могут привести к изменениям в высший мозг функция, особенно обучение и память. Однако в качестве новой области в нейробиология, большая часть знаний о несинаптической пластичности является неопределенной и все еще требует дальнейшего изучения, чтобы лучше определить ее роль в функции и поведении мозга.
Против. синаптическая пластичность
Нейропластичность это способность определенной части или области нейрона изменяться в силе с течением времени. Существует две общепризнанные категории пластичности: синаптическая и несинаптическая. Синаптическая пластичность имеет дело непосредственно с силой связи между двумя нейронами, включая количество нейротрансмиттер освобожден от пресинаптический нейрон, и ответ, сгенерированный в постсинаптический нейрон. Несинаптическая пластичность включает изменение возбудимости нейронов в аксон, дендриты, и сома отдельного нейрона, удаленного от синапса.
Синаптическая пластичность
Синаптическая пластичность - это способность синапса между двумя нейронами изменять силу с течением времени. Синаптическая пластичность вызывается изменениями в использовании синаптического пути, а именно, частоты синаптических потенциалов и рецепторов, используемых для передачи химических сигналов. Синаптическая пластичность играет большую роль в обучении и памяти мозга. Синаптическая пластичность может происходить посредством внутренних механизмов, в которых изменения силы синапсов происходят из-за его собственной активности, или посредством внешних механизмов, в которых изменения силы синапсов происходят через другие нервные пути. Кратковременная тормозная синаптическая пластичность часто возникает из-за ограниченного нейротрансмиттер снабжение синапсом, и длительное ингибирование может происходить из-за снижения экспрессии рецептора в постсинаптическая клетка. Краткосрочная комплементарная синаптическая пластичность часто возникает из-за остаточного или увеличенного ионного потока в пресинаптическом или постсинаптическом окончании, в то время как долговременная синаптическая пластичность может происходить из-за повышенной продукции AMPA и NMDA рецепторы глутамата, среди прочего, в постсинаптической клетке.[1]
Несинаптическая пластичность
Для сравнения, несинаптическая пластичность - менее известная и несколько новая и продолжающаяся область исследований в нейробиологии. Это проявляется в изменении характеристик несинаптических структур, таких как сома (биология), аксон или дендриты. Несинаптическая пластичность может иметь краткосрочные или долгосрочные последствия. Один из способов возникновения этих изменений - модификация стробированные каналы в дендритах и аксоне, что изменяет интерпретацию возбуждающих или тормозных потенциалов, передаваемых в клетку. Например, аксональная несинаптическая пластичность может наблюдаться, когда потенциал действия не достигает пресинаптического терминала из-за низкой проводимости или накопления ионов. [2]

Синергетические эффекты
Общие возбуждающие эффекты
Было показано, что несинаптическая и синаптическая пластичность работают одновременно различными способами, вызывая стимулирующие эффекты в нейроне. Это включает генерацию спайков, продукт несинаптической регуляции калиевых и других пресинаптических ионных каналов, которые увеличивают реакцию возбуждающий постсинаптический потенциал через высвобождение нейротрансмиттера и увеличение потенциала действия.[3] Несинаптическая дендритная пластичность также усиливает эффекты синаптической пластичности за счет расширения потенциала действия. Как будет сказано ниже, мозговой нейротрофический фактор (BNDF) вырабатывается нейронами для координации несинаптической и синаптической пластичности.[4] Несинаптические изменения в сомальном теле, аксоне или дендритах нейрона неразрывно связаны с синаптической силой.
Интеграция в памяти и обучении
Хотя о роли синаптической пластичности в памяти и обучении известно гораздо больше, синаптическая и несинаптическая пластичность важны для объем памяти и учусь в мозгу. Есть много свидетельств того, что оба механизма работают для достижения наблюдаемых эффектов синергетически. Ключевым примером этого является формирование памяти в синапсе, при котором модификация пресинаптических механизмов высвобождения и постсинаптических рецепторов влияет либо на долгосрочную потенциацию, либо на депрессию. Непрерывный сомальный деполяризация, с другой стороны, был предложен как метод обучения поведению и памяти с помощью несинаптической пластичности. Несинаптическая пластичность также увеличивает эффективность формирования синаптической памяти за счет регуляции потенциалзависимые ионные каналы. Несинаптическая пластичность - это механизм, ответственный за модификации этих каналов в аксоне, приводящий к изменению силы потенциала действия нейронов, неизменно влияя на силу синаптических механизмов и, таким образом, на глубину и длину кодирования памяти. [5][6]
Регулирование синаптической пластичности
Несинаптическая пластичность также имеет способность регулировать эффекты синаптической пластичности посредством негативный отзыв механизмы. Изменение количества и свойств ионных каналов в аксоне или дендритах может уменьшить эффекты гиперстимулированного синапса.[5][6] В случае чрезмерного перевозбуждения этих ионных каналов произойдет обратный поток ионов в ячейку, что приведет к эксайтотоксичность и гибель клеток апоптоз или же некроз.[7]
Внутренние механизмы
Несинаптические нейрональные области, такие как аксон, также обладают качествами, которые влияют на синапс. Эти важные механизмы включают задержку деполяризации, которой подвергается потенциал действия при движении вниз по аксону. Это внутреннее качество замедляет распространение потенциалов действия и связано с движением деполяризующего тока по цитоплазме и периодическим размещением натриевых каналов на Узлы Ранвье. Эти механизмы существуют всегда, но могут меняться в зависимости от состояния клеточной сомы, аксона и дендритов в данный момент. Следовательно, латентность или задержка распространения потенциалов действия или EPSP могут быть переменными. Каждый возбуждающий постсинаптический потенциал который передается в постсинаптическую клетку, сначала передается через потенциал действия вниз по аксону в пресинаптической клетке, и, таким образом, несинаптическая пластичность по своей сути влияет на синаптическую пластичность.[1]
Типы
Собственная возбудимость нейрона
Возбудимость нейрона в любой момент зависит от внутренних и внешних условий клетки во время стимуляции. Поскольку нейрон обычно получает одновременно несколько входящих сигналов, распространение потенциала действия зависит от интеграции всех входящих ВПСП и IPSP прибытие в аксональный бугорок. Если сумма всех возбуждающих и тормозных сигналов деполяризовать клеточная мембрана до порогового напряжения, запускается потенциал действия. Изменение собственной возбудимости нейрона изменит функцию этого нейрона.
Генерация шипа
Несинаптическая пластичность оказывает возбуждающее действие на генерацию шипы. Увеличение генерации спайков коррелировало с уменьшением пиковый порог,[3] ответ от несинаптической пластичности. Этот ответ может быть результатом модуляции некоторых пресинаптических K+ (ион калия) токи (IА,ЯК, Са, и яKs), которые увеличивают возбудимость сенсорных нейронов, расширяют потенциал действия и усиливают нейротрансмиттер релиз. Эти модуляции K+ проводимости служат общими механизмами регулирования возбудимости и синаптическая сила.[5]
Регулирование синаптической пластичности
Несинаптическая пластичность была связана с синаптической пластичностью как через синергетические, так и через регуляторные механизмы. Степень синаптической модификации определяет полярность несинаптических изменений, влияющих на изменение клеточной возбудимости. Умеренные уровни синаптической пластичности вызывают несинаптические изменения, которые будут действовать синергетически с синаптическими механизмами, усиливая реакцию. И наоборот, более устойчивые уровни синаптической пластичности будут вызывать несинаптические ответы, которые будут действовать как механизм отрицательной обратной связи. Механизмы отрицательной обратной связи работают для защиты от насыщения или подавления активности цепи в целом.[5]
Аксональная модуляция
Аксональная модуляция - это тип пластичности, при котором количество, активность или расположение ионные каналы в аксоне изменения. Это заставляет нейрон вести себя по-разному при стимуляции. Модуляция ионных каналов - это ответ на изменение частот стимуляции нейрона.
Пластичность распространения
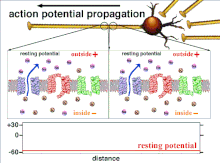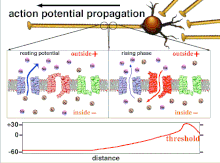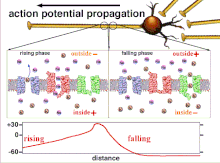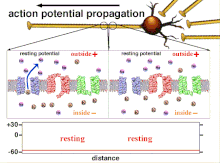
Поскольку это суммирование потенциалов действия, которое в конечном итоге приводит к пересечению пороговой поляризации, временные отношения различных входных сигналов очень важны для определения того, сработает ли постсинаптический нейрон и когда. Со временем время, необходимое потенциалу действия для распространения по длине конкретного аксона, может измениться. В одном эксперименте многоэлектродные решетки были использованы для измерения времени, необходимого для перехода потенциалов действия от одного электрода к другому, называемого задержкой. Затем нейроны стимулировали, и значение задержки регистрировалось с течением времени. Значения латентности менялись со временем, предполагая, что аксональная пластичность влияет на распространение потенциалов действия.[8]
Маневровые
Шунтирование - это процесс, при котором каналы аксональных ионов открываются во время пассивного потока (не требуя ионный насос ) подпорога деполяризация вниз по аксону. Обычно возникает в точках ветвления аксонов,[9] время открытия этих каналов по мере поступления подпорогового сигнала в область вызывает гиперполяризацию, которая вносится в пассивно протекающую деполяризацию. Следовательно, клетка способна контролировать, через какие ветви аксона протекает ток подпороговой деполяризации, в результате чего некоторые ветви аксона становятся более гиперполяризованными, чем другие. Эти разные мембранные потенциалы заставляют определенные области нейрона быть более возбудимыми, чем другие, в зависимости от конкретного местоположения и возникновения шунтирования.
Высокочастотная стимуляция
Краткосрочные эффекты: высокочастотная стимуляция нейрона в течение короткого периода времени увеличивает возбудимость нейрона за счет снижения количества Напряжение требуется для срабатывания потенциала действия.[3]Высокочастотная стимуляция приводит к увеличению внутриклеточной концентрации ионов натрия и кальция за счет многократного открытия потенциалзависимые натриевые и кальциевые каналы в аксоне и терминале. Чем больше частота стимулов, тем меньше времени между каждым стимулом у клетки переполяризовать и вернуться к нормальной жизни потенциал покоя. Следовательно, потенциал покоя становится более деполяризованным, а это означает, что для активации потенциала действия требуется меньший ток деполяризации.
Однако эта модуляция обычно длится очень недолго. Если стимуляция прекратится, нейрон вернется к своему первоначальному потенциалу покоя, так как ионные каналы и насосы иметь достаточно времени, чтобы оправиться от последнего раздражителя.
Долгосрочные эффекты: высокочастотная стимуляция нейрона в течение длительного периода времени вызывает два результирующих нейронных изменения. Вначале нейрон реагирует так же, как и при кратковременной стимуляции, повышением возбудимости. Продолжение высокочастотной стимуляции после этого момента приводит к резкому, необратимому изменению возбудимости. Когда концентрация натрия в аксоне достигает достаточно высокого уровня, натриевые / кальциевые насосы меняют направление потока, в результате чего кальций попадает в клетку по мере того, как натрий выводится наружу. Повышенная концентрация кальция (и последующая деполяризация мембраны) инактивирует натриевые каналы и нацелены на них эндоцитоз и лизосомный гидролиз.[10]Это приводит к значительному уменьшению аксональных натриевых каналов, которые необходимы для распространения потенциала действия. Если стимуляция продолжается, в конце концов нейрон перестанет передавать потенциалы действия и умрет. Смерть нейронов из-за чрезмерной стимуляции называется эксайтотоксичность.
Низкочастотная стимуляция
Краткосрочные эффекты: все живые нейроны имеют базальную скорость распространения потенциала действия и синаптического высвобождения. Таким образом, низкочастотная стимуляция нейрона в краткосрочной перспективе аналогична активности нейрона в состоянии покоя в головном мозге. Никаких серьезных изменений во внутренней возбудимости нейрона не происходит.
Долгосрочные эффекты: низкочастотная стимуляция нейрона в течение длительного периода времени снижает возбудимость нейрона за счет активации кальций-зависимых фосфатаз, которые маркируют Рецепторы AMPA для интернализации.[11] Низкочастотная стимуляция приводит к снижению уровня кальция в клетке. При низких концентрациях кальция активные кальций-зависимые фосфатазы преобладают над кальций-зависимыми киназами. По мере активации большего количества фосфатаз они маркируют большее количество рецепторов AMPA для интернализации через эндоцитоз. Поскольку рецепторы AMPA являются одними из основных возбуждающих рецепторов на нейронах, удаление их с клеточной мембраны эффективно подавляет клетку (если клетка не может реагировать на возбуждающие сигналы, она не может генерировать собственный потенциал действия). Таким образом, низкочастотная стимуляция может полностью изменить эффекты долгосрочное потенцирование,[12] однако эти концепции обычно считаются типами синаптической пластичности.
Гомеостатическая и геббовская пластичность
Центральная нервная система (ЦНС) нейроны интегрируют сигналы от многих нейронов. В краткосрочной перспективе важно иметь изменения в активности нейрона, потому что именно так информация передается в нервной системе (Хеббийская пластичность ). Однако для долгосрочной устойчивости дрейф в сторону возбудимости или невозбудимости нарушит способность схемы передавать информацию (гомеостатическая пластичность ). Долгосрочная потенциация (ДП) вызывает более высокую частоту возбуждения постсинаптических нейронов. Была выдвинута гипотеза, что внутренние свойства нейрона должны быть организованы таким образом, чтобы максимально использовать динамический диапазон, действуя как гомеостатический механизм.[13] Однако было показано, что внутренняя возбудимость следует логнормальному распределению, которое требует активного обучения Хебба.[14] Исследования in vitro показали, что когда спонтанная активность нейронных культур подавляется, нейроны становятся гипервозбудимыми, и, когда увеличение активности индуцируется в течение длительного периода, скорость возбуждения культуры падает.[15][16] Напротив, существует множество доказательств того, что противоположная форма регулирования, обучение по Хебби, или LTP-IE / LTD-IE, также имеет место.[17] и теоретические аргументы показывают, что Хеббийская пластичность должна быть доминирующей формой пластичности и для внутренней возбудимости.[14] С гомеостатическая пластичность также происходит между отдельными синапсами,[18] более ранняя точка зрения, предполагающая, что гомеостатическая пластичность и внутренняя пластичность взаимосвязаны, оказалась несовместимой с доказательствами.
Механизм
Одним из механизмов сохранения динамического диапазона нейрона является синаптическое масштабирование, гомеостатическая форма пластичности, которая восстанавливает активность нейронов до нормальных «исходных» уровней путем изменения постсинаптического ответа синапсов нейрона в зависимости от активности. Гомеостатическая модуляция собственной возбудимости нейрона - еще один способ поддержания стабильности. Регулирование ионная проводимость может быть достигнуто несколькими способами, в основном за счет выпуска нейромодуляторов, таких как дофамин, серотонин и Т. Д.[19] Другой способ - это контролируемое высвобождение мозговых нейротрофический фактор (БДНФ). Также было обнаружено, что BDNF влияет на синаптическое масштабирование, предполагая, что этот нейротрофический фактор может быть ответственным за координацию синаптических и несинаптических механизмов в гомеостатической пластичности.[4]
Дендритная возбудимость
Дендриты - это регионы, ответственные за интеграцию входов от других нейроны. Один из способов, которым нейроны манипулируют интеграционными свойствами дендритов, - это изменение количества и свойств потенциалзависимых ионных каналов. Вызывание долгосрочной потенциации (ДП) в определенных синапс, приводит к увеличению возбудимости дендритных ветвей, специфичных для этого синапса.[20] Возбудимость дендритов важна для распространения и интеграции синаптических сигналов. Считается, что дендритная возбудимость способствует E-S-потенцированию или увеличению вероятности того, что данный вход приведет к срабатыванию потенциала действия.[21]
Известно, что изменения возбудимости дендритов влияют на обратное распространение потенциала действия. Потенциалы действия начинаются около бугра аксона и распространяются по длине аксона, но они также распространяются назад через сому в дендритную ветвь. Активное обратное распространение зависит от ионных каналов, и изменение плотности или свойств этих каналов может повлиять на степень распространения сигнала. ослабленный.[21] Пластичность обратного распространения в дендритах происходит менее чем за одну минуту и продолжается более 25 минут.[22] Обратное распространение - это метод передачи сигналов в синапсы о том, что был активирован потенциал действия. Это важно для пластичность, зависящая от времени всплеска.
Собственная пластичность
Внутренняя пластичность - это форма зависимой от деятельности пластичности, отличной от синаптическая пластичность, который включает изменения в синапсе между двумя нейронами, а не изменения электрических свойств внутри одного нейрона.[23][24] Есть несколько тесно связанных явлений, которые могут повлиять на возбудимость нейрона, например: нейромодуляция, структурная пластичность, кратковременная пластичность, обусловленная кинетикой канала, и нейронное развитие.[25][26] Нет единого мнения относительно количества, которое регулирует внутренняя пластичность, например частота возбуждения нейрона, его усиление или внутренняя концентрация кальция. С функциональной точки зрения внутренняя пластичность может позволить нейронам узнавать интенсивность стимулов и представлять эту статистику интенсивности в их возбудимости.[27][28] Внутренняя пластичность способствует кодированию памяти и дополняет другие формы зависимой от активности пластичности, включая синаптическая пластичность.[29]
Высшая функция мозга
Долговременная ассоциативная память
Экспериментальные доказательства
Эксперимент Кеменеса и другие.[2] продемонстрировал, что во внешнем модуляторный нейрон, несинаптическая пластичность влияет на выражение долговременной ассоциативной памяти. Взаимосвязь между несинаптической пластичностью и памятью оценивалась с использованием гигантских клеток головного мозга (CGC). Деполяризация от условных раздражителей усиливается ответ нейронной сети. Эта деполяризация длилась до тех пор, пока Долгосрочная память. Устойчивая деполяризация и проявление поведенческой памяти произошли более чем через 24 часа после тренировки, что указывает на долгосрочные эффекты. В этом эксперименте электрофизиологический выражение долговременной памяти было вызвано условным стимулом кормовой реакцией. CGC были значительно более деполяризованы у обученных организмов, чем в контрольной группе, что указывает на связь с изменениями в обучении и возбудимости. Когда CGC были деполяризованы, они показали повышенный ответ на условные стимулы и более сильный ответ на фиктивное питание. Это показало, что деполяризации достаточно, чтобы вызвать значительную пищевую реакцию на условные раздражители. Кроме того, не наблюдалось значительной разницы в скорости питания между кондиционированными организмами и организмами, которые были искусственно деполяризованы, что подтверждает, что деполяризации достаточно для формирования поведения, связанного с долговременной памятью.[2]
Память для хранения
Несинаптическая активность клетки обычно выражается в изменении возбудимости нейронов. Это происходит за счет модуляции компонентов мембраны, таких как покой и стробированные каналы и ионные насосы. Считается, что несинаптические процессы участвуют в хранении в памяти. Один из возможных механизмов этого действия заключается в маркировке нейрона, который недавно был активен, изменениями возбудимости. Это поможет связать стимулы, разделенные во времени. Другой потенциальный механизм исходит из вычислительной модели, которая указывает, что несинаптическая пластичность может инициировать модификации в обучении, поскольку изменения возбудимости могут регулировать порог синаптической пластичности.[5]
Емкость систем хранения синаптической памяти очень велика, что делает их привлекательным механизмом для изучения. Примерно 104 синапсов на нейрон и 1011 нейроны человеческого мозга.[23] Несинаптическую пластичность часто упускают из виду просто потому, что ее емкость хранения не так высока. Регулирование плотности ионных каналов в аксоне и соме нейрона изменит пропускную способность и повлияет на все синапсы. Следовательно, его емкость хранения будет значительно меньше, чем у синаптической пластичности.
Хотя его емкость хранения слишком мала, чтобы сделать его единственным механизмом хранения, несинаптическая пластичность может способствовать способам синаптического хранения. Было показано, что модуляция ионных каналов может происходить в таких небольших областях, как конкретные дендриты.[20] Эта специфичность делает емкость несинаптической пластичности большей, чем если бы это была модуляция целого нейрона. Процедурные воспоминания хорошо подходят для этого типа системы хранения, потому что они не требуют высокой специфичности, как декларативная память. Обобщение двигательных задач и обусловленный стимулы могут быть эффективным способом хранения этой информации.[23]
Учусь
Изменения возбудимости от обучения, которые действуют как часть след памяти делают это в качестве праймеров для инициирования дальнейших изменений в нейронах или с помощью механизма кратковременного хранения кратковременной памяти. Несинаптическая пластичность может возникать во время обучения в результате клеточных процессов, хотя время, постоянство и взаимосвязь между несинаптической пластичностью и синаптическим выходом плохо изучены. Исследования показали, что несинаптическая пластичность играет косвенную, но важную роль в формировании воспоминаний. Несинаптическая пластичность, вызванная обучением, связана с деполяризацией сомы.[5]
Классическое кондиционирование
Эксперименты показали, что несинаптические изменения происходят во время условное обучение. Вуди и другие.[30] продемонстрировал, что моргание (EBC), форма классического кондиционирования для изучения нейронных структур и механизмов, лежащих в основе обучения и памяти, у кошек связана с повышенной возбудимостью и повышением активности нейронов в нейронах. сенсомоторные области коры и в лицевое ядро. Было замечено, что повышение возбудимости от классического кондиционирования продолжалось после прекращения реакции. Это говорит о том, что повышенная возбудимость может функционировать как механизм хранения памяти.[5]
При кондиционировании моргания у кроликов несинаптические изменения происходили на протяжении всего периода жизни. дорсальный гиппокамп. Это указывает на то, что хотя одних только изменений возбудимости недостаточно для объяснения процессов хранения в памяти, несинаптическая пластичность может быть механизмом хранения для фаз памяти, ограниченных временем. Несинаптические изменения влияют на другие типы пластичности, связанные с памятью. Например, несинаптическое изменение, такое как деполяризация мембранный потенциал покоя в результате условного обучения может вызвать синаптическую пластичность в будущем обучении.[5]
Обучение правилам и экономия
Способность изучать правила зависит от несинаптической пластичности. Одно исследование было направлено на то, чтобы научить крыс различать различные запахи, и потребовалось несколько дней, чтобы научить их различать первую пару запахов. Однако, узнав это, крыса смогла научиться различать разные запахи намного быстрее. Изменения возбудимости пирамидных нейронов у этих крыс наблюдались в течение трех дней после обучения. Эти изменения со временем исчезли, что свидетельствует о том, что нейроны участвовали в обучении правилам, а не в хранении памяти.[5] Даудал и Дебанн попытались определить, применимы ли те же правила обучения и механизмы индукции, определенные для синаптической пластичности, к несинаптической пластичности, влияющей на ионные каналы. Они определили, что несинаптическая и синаптическая пластичность имеют общие правила обучения и пути индукции, например, Рецептор NMDA зависимая долговременная потенциация (ДП) и длительная депрессия (ООО). Они также показали, что несинаптическая и синаптическая пластичность синергетически образуют когерентный инграмма для хранения следов памяти.[22]
Экономия - это способность повторно изучать забытую информацию намного быстрее, чем она была изучена изначально. Несинаптическая пластичность - возможный механизм этого эффекта экономии. Во время тренировочных процедур многие нейроны испытывают повышение собственной возбудимости. Это повышение возбудимости сохраняется даже после исчезновения памяти.[5][23]
Вещественная зависимость
Злоупотребление наркотиками обычно влияет на мезолимбическая система, а точнее, путь вознаграждения нервной системы. Среди распространенных наркотиков никотин является одним из сильнейших агонистов никотинового холинергического синапса.[31] Никотин, конкурируя с ацетилхолином (ACh), действует через несинаптические, претерминальные, никотиновый рецептор ацетилхолина (nAChR), чтобы инициировать изменение мембранного потенциала и размножать внутриклеточный Ca2+ сигнал, тем самым стимулируя высвобождение нейротрансмиттеров. Специфическая и характерная роль активности nAChR, опосредованной кальциевым током, имеет зависимость от напряжения, отличную от другой Ca2+ проницаемые ионные каналы, а также различное временное и пространственное распределение, и в результате несинаптическая активность nAChR усиливает индукцию синаптической потенциации, способствуя обучению зависимости от веществ.[32]
Приложения к болезни
После повреждения
Несинаптическая пластичность может смягчать эффекты повреждение мозга. Когда один из вестибулярные нервы поврежден, несоответствие в скорости возбуждения нейронов в вестибулярные ядра вызывает ненужные вестибулярные рефлексы. Симптомы этого повреждения со временем исчезают. Вероятно, это связано с модификациями собственной возбудимости нейронов вестибулярного ядра.[23][33]
Судорожная активность
Несинаптическая пластичность также играет ключевую роль в захват Мероприятия. Лихорадочные припадки, припадки, вызванные лихорадкой в раннем возрасте, могут привести к повышенной возбудимости нейронов гиппокампа. Эти нейроны становятся очень чувствительными к возбудителям конвульсий. Было показано, что судороги в раннем возрасте могут предрасполагать один к большему количеству приступов через несинаптические механизмы.[34]
Травмы, в том числе Инсульт что приводит к корковый травма, часто приводит к эпилепсия. Повышенная возбудимость и NMDA Проведение приводит к эпилептической активности, предполагая, что несинаптическая пластичность может быть механизмом, посредством которого эпилепсия индуцируется после травмы.[35]
Аутизм
Вальпроевая кислота (VPA) - это средство от эпилепсия, мигрень, и биполярное расстройство это было связано со многими условиями, включая аутизм. An животная модель аутизма существует, в котором беременным крысам вводят VPA. У потомства есть черты, похожие на черты людей с аутизмом. Вскоре после рождения у этих животных наблюдается снижение возбудимости и повышенная возбудимость. NMDA токи. Эти эффекты исправляются на более поздних этапах жизни. Изменения собственной возбудимости у этих животных помогли нейтрализовать эффекты повышенных токов NMDA на сетевую активность, форму гомеостатической пластичности. Считается, что это помогает смягчить пагубные последствия, которые могут иметь повышенные токи NMDA.[36]
Текущие и будущие исследования
Необходимы дополнительные исследования, чтобы получить более широкое представление о несинаптической пластичности. Темы, требующие дальнейшего изучения с января 2010 г.[Обновить] включают:
- Изменения локальной возбудимости по сравнению с глобальными в нейронных сетях и поддержание следов памяти[5]
- Специфика индукции изменений возбудимости, зависящей от обучения[5]
- Манипуляции с изменениями возбудимости, зависящими от обучения, с помощью фармацевтических препаратов или генетических мутаций и их влияния на следы памяти[5]
- Сходства между молекулярными механизмами синаптической и несинаптической пластичности[5]
- Сравнение паттернов несинаптической пластичности in vivo с результатами in vitro[5]
- Изменения в экспрессии генов, вызванные нейронной активностью[37]
Рекомендации
- ^ а б Бирн, Джон Х. (1997). «Синаптическая пластичность». Неврология онлайн. Медицинская школа UT в Хьюстоне. Получено 28 октября, 2011.
- ^ а б c Кеменес И., Штрауб В.А., Никитин Е.С., Старас К., О'Ши М., Кеменес Г., Бенджамин П.Р. (июль 2006 г.). «Роль отсроченной несинаптической нейрональной пластичности в долговременной ассоциативной памяти». Текущая биология. 16 (13): 1269–79. Дои:10.1016 / j.cub.2006.05.049. PMID 16824916. S2CID 16726488.
- ^ а б c Hansel C, Linden DJ, D'Angelo E (май 2001 г.). «Beyond parallel fiber LTD: разнообразие синаптической и несинаптической пластичности в мозжечке». Природа Неврология. 4 (5): 467–75. Дои:10.1038/87419. PMID 11319554. S2CID 13919286.
- ^ а б Десаи Н.С., Резерфорд Л.С., Турриджиано Г.Г. (1999). «BDNF регулирует внутреннюю возбудимость кортикальных нейронов». Обучение и память. 6 (3): 284–91. Дои:10.1101 / лм. 6.3.284 (неактивно 2020-10-22). ЧВК 311300. PMID 10492010.CS1 maint: DOI неактивен по состоянию на октябрь 2020 г. (связь)
- ^ а б c d е ж грамм час я j k л м п о Mozzachiodi R, Бирн JH (январь 2010 г.). «Больше, чем синаптическая пластичность: роль несинаптической пластичности в обучении и памяти». Тенденции в неврологии. 33 (1): 17–26. Дои:10.1016 / j.tins.2009.10.001. ЧВК 2815214. PMID 19889466.
- ^ а б Дебанн Д., Копысова И.Л., Браш Х., Ферран Н. (сентябрь 1999 г.). «Стробирование распространения потенциала действия с помощью аксональной А-подобной калиевой проводимости в гиппокампе: новый тип несинаптической пластичности». Журнал физиологии, Париж. 93 (4): 285–96. Дои:10.1016 / S0928-4257 (00) 80057-1. PMID 10574118. S2CID 30737916.
- ^ Szydlowska K, Tymianski M (февраль 2010 г.). «Кальций, ишемия и эксайтотоксичность». Клеточный кальций. 47 (2): 122–9. Дои:10.1016 / j.ceca.2010.01.003. PMID 20167368.
- ^ Баккум DJ, Чао З.С., Поттер С.М. (май 2008 г.). «Долговременная зависимая от активности пластичность задержки и амплитуды распространения потенциала действия в корковых сетях». PLOS ONE. 3 (5): e2088. Bibcode:2008PLoSO ... 3.2088B. Дои:10.1371 / journal.pone.0002088. ЧВК 2324202. PMID 18461127.

- ^ Дебанн Д., Гевилер Б.Х., Томпсон С.М. (1996). «Синаптическая и несинаптическая пластичность между отдельными пирамидными клетками в гиппокампе крысы in vitro». Журнал физиологии, Париж. 90 (5–6): 307–9. Дои:10.1016 / s0928-4257 (97) 87903-х. PMID 9089497. S2CID 31639170.
- ^ Ахмед З., Верашко А. (январь 2009 г.). «Зависимая от активности аксональная пластичность: эффекты электрической стимуляции на потенциалы сложного действия, регистрируемые нервной системой мыши in vitro». Открытый журнал неврологии. 3 (1): 6. Bibcode:2009ONMJ .... 1 .... 6P. Дои:10.2174/1874082000903010001. S2CID 14646213.
- ^ Lin JW, Ju W, Foster K, Lee SH, Ahmadian G, Wyszynski M и др. (Декабрь 2000 г.). «Отчетливые молекулярные механизмы и различные эндоцитотические пути интернализации рецептора AMPA». Природа Неврология. 3 (12): 1282–90. Дои:10.1038/81814. PMID 11100149. S2CID 18109725.
- ^ О'Делл Т.Дж., Кандел Э.Р. (1994). «Низкочастотная стимуляция стирает LTP за счет активации протеинфосфатаз, опосредованной рецептором NMDA». Обучение и память. 1 (2): 129–139. Дои:10.1101 / лм. 1.2.129 (неактивно 2020-10-22). PMID 10467591.CS1 maint: DOI неактивен по состоянию на октябрь 2020 г. (связь)
- ^ Стеммлер М., Кох С. (июнь 1999 г.). «Как напряжение, зависящее от проводимости, может адаптироваться, чтобы максимизировать информацию, закодированную частотой нейрональных импульсов». Природа Неврология. 2 (6): 521–7. Дои:10.1038/9173. PMID 10448216. S2CID 15134486.
- ^ а б Шелер Г (2017). «Логарифмические распределения доказывают, что внутреннее обучение - хеббийское». F1000 Исследования. 6: 1222. arXiv:1410.5610. Дои:10.12688 / f1000research.12130.2. ЧВК 5639933. PMID 29071065.
- ^ Corner MA, Ramakers GJ (январь 1992 г.). «Самопроизвольное возбуждение как эпигенетический фактор в развитии мозга - физиологические последствия хронического воздействия тетродотоксина и пикротоксина на культивируемые нейроны неокортекса крысы». Исследование мозга. Развитие мозга. 65 (1): 57–64. Дои:10.1016 / 0165-3806 (92) 90008-К. PMID 1551233.
- ^ Турриджиано Г.Г., Лесли К.Р., Десаи Н.С., Резерфорд Л.С., Нельсон С.Б. (февраль 1998 г.). «Активно-зависимое масштабирование квантовой амплитуды в нейронах неокортекса». Природа. 391 (6670): 892–6. Bibcode:1998Натура.391..892Т. Дои:10.1038/36103. PMID 9495341. S2CID 4328177.
- ^ Махон С., Шарпье С. (август 2012 г.). «Двунаправленная пластичность собственной возбудимости контролирует эффективность сенсорных входов в нейронах коры ствола 5 слоя in vivo». Журнал неврологии. 32 (33): 11377–89. Дои:10.1523 / JNEUROSCI.0415-12.2012. ЧВК 6621180. PMID 22895720.
- ^ Кек Т., Келлер Г.Б., Якобсен Р.И., Эйзель Ю.Т., Бонхоффер Т., Хюбенер М. (октябрь 2013 г.). «Синаптическое масштабирование и гомеостатическая пластичность в зрительной коре мышей in vivo». Нейрон. 80 (2): 327–34. Дои:10.1016 / j.neuron.2013.08.018. PMID 24139037. S2CID 13151568.
- ^ Шелер Г (2014). «Изучение собственной возбудимости в нейронах со средними шипами». F1000 Исследования. 2: 88. Дои:10.12688 / f1000research.2-88.v2. ЧВК 4264637. PMID 25520776.
- ^ а б Фрик А., Маги Дж., Джонстон Д. (февраль 2004 г.). «ДП сопровождается повышенной локальной возбудимостью дендритов пирамидных нейронов». Природа Неврология. 7 (2): 126–35. Дои:10.1038 / nn1178. PMID 14730307. S2CID 11964239.
- ^ а б Sjöström PJ, Rancz EA, Roth A, Häusser M (апрель 2008 г.). «Дендритная возбудимость и синаптическая пластичность». Физиологические обзоры. 88 (2): 769–840. Дои:10.1152 / Physrev.00016.2007. PMID 18391179. S2CID 1261675.
- ^ а б Даудал Г., Дебанн Д. (2003). «Долговременная пластичность собственной возбудимости: правила и механизмы обучения». Обучение и память. 10 (6): 456–65. Дои:10,1101 / лм. 64103. PMID 14657257.
- ^ а б c d е Zhang W, Linden DJ (ноябрь 2003 г.). «Другая сторона инграммы: обусловленные опытом изменения внутренней возбудимости нейронов». Обзоры природы. Неврология. 4 (11): 885–900. Дои:10.1038 / номер 1248. PMID 14595400. S2CID 17397545.
- ^ Дебанн Д., Инглеберт Ю., Русье М. (февраль 2019 г.). «Пластичность собственной нейрональной возбудимости» (PDF). Текущее мнение в нейробиологии. 54: 73–82. Дои:10.1016 / j.conb.2018.09.001. PMID 30243042. S2CID 52812190.
- ^ Триеш, Йохен. "Synergies between intrinsic and synaptic plasticity in individual model neurons. " Достижения в системах обработки нейронной информации. 2004.
- ^ W. Zhang and D. J. Linden. The other side of the engram: Experience-driven changes in neuronal intrinsic excitability. Nature Reviews Neuroscience, 4:885-900, 2003.
- ^ T. Monk, C. Savin, and J. Lucke. "Optimal neural inference of stimulus intensities. " Scientific reports, 8:1, 2018.
- ^ T. Monk, C. Savin, and J. Lucke. Neurons equipped with intrinsic plasticity learn stimulus intensity statistics. Достижения в системах обработки нейронной информации. 2016.
- ^ Grasselli G, Boele HJ, Titley HK, Bradford N, van Beers L, Jay L, et al. (Январь 2020 г.). "SK2 channels in cerebellar Purkinje cells contribute to excitability modulation in motor-learning-specific memory traces". PLOS Биология. 18 (1): e3000596. Дои:10.1371/journal.pbio.3000596. ЧВК 6964916. PMID 31905212.
- ^ Woody CD, Black-Cleworth P (November 1973). "Differences in excitability of cortical neurons as a function of motor projection in conditioned cats". Журнал нейрофизиологии. 36 (6): 1104–16. Дои:10.1152/jn.1973.36.6.1104. PMID 4761722.
- ^ Kauer JA, Malenka RC (November 2007). "Synaptic plasticity and addiction". Обзоры природы. Неврология. 8 (11): 844–58. Дои:10.1038/nrn2234. PMID 17948030. S2CID 38811195.
- ^ Dani JA, Ji D, Zhou FM (August 2001). "Synaptic plasticity and nicotine addiction". Нейрон. 31 (3): 349–52. Дои:10.1016/S0896-6273(01)00379-8. PMID 11516393. S2CID 10062998.
- ^ Darlington CL, Dutia MB, Smith PF (June 2002). "The contribution of the intrinsic excitability of vestibular nucleus neurons to recovery from vestibular damage". Европейский журнал нейробиологии. 15 (11): 1719–27. Дои:10.1046/j.1460-9568.2002.02024.x. PMID 12081651. S2CID 19939794.
- ^ Bender RA, Dubé C, Gonzalez-Vega R, Mina EW, Baram TZ (2003). "Mossy fiber plasticity and enhanced hippocampal excitability, without hippocampal cell loss or altered neurogenesis, in an animal model of prolonged febrile seizures". Гиппокамп. 13 (3): 399–412. Дои:10.1002/hipo.10089. ЧВК 2927853. PMID 12722980.
- ^ Bush PC, Prince DA, Miller KD (October 1999). "Increased pyramidal excitability and NMDA conductance can explain posttraumatic epileptogenesis without disinhibition: a model". Журнал нейрофизиологии. 82 (4): 1748–58. Дои:10.1152/jn.1999.82.4.1748. PMID 10515964. S2CID 5927906.
- ^ Walcott EC, Higgins EA, Desai NS (September 2011). "Synaptic and intrinsic balancing during postnatal development in rat pups exposed to valproic acid in utero". Журнал неврологии. 31 (37): 13097–109. Дои:10.1523/JNEUROSCI.1341-11.2011. ЧВК 6623264. PMID 21917793.
- ^ Leslie JH, Nedivi E (August 2011). "Activity-regulated genes as mediators of neural circuit plasticity" (PDF). Прогресс в нейробиологии. 94 (3): 223–37. Дои:10.1016/j.pneurobio.2011.05.002. HDL:1721.1/102275. ЧВК 3134580. PMID 21601615.