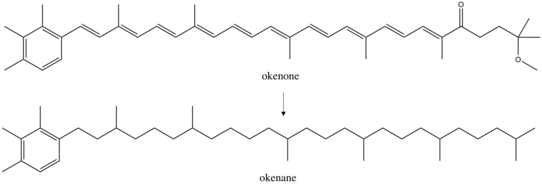
Окенане - Okenane
Окенане, то диагенетический конечный продукт окенона, является биомаркер за Chromatiaceae, то фиолетовые серные бактерии.[1] Эти аноксигенные фототрофы использовать свет для получения энергии и сульфид как их донор электронов и сера источник. Открытие окенана в морские отложения подразумевает прошлое эвксиновый среда, где стояли водяные столбы аноксический и сульфидный. Это потенциально чрезвычайно важно для реконструкции прошлых океанических условий, но до сих пор окенан был идентифицирован только в одном Палеопротерозой Образец породы (возраст 1,6 миллиарда лет) из Северной Австралии.[2][3]
Фон

Океноне - это каротиноид,[4] класс пигменты повсеместно фотосинтетический организмы. Эти сопряженные молекулы действовать как аксессуары в легкий уборочный комплекс. Известно более 600 каротиноидов, каждый из которых имеет множество функциональные группы которые меняют их спектр поглощения. Окенон, по-видимому, лучше всего приспособлен к желто-зеленому переходу (520 нм) видимый спектр, улавливая свет внизу морской планктон В океане. Эта глубина варьируется в зависимости от структуры сообщества водной толщи. Обзор микробное цветение найденный Chromatiaceae где-то на глубине от 1,5 м до 24 м, но более 75% произошло на глубине более 12 метров.[5] Далее планктонные серные бактерии занимают другие ниши: зеленые серные бактерии, то Chlorobiaceae, которые производят каротиноид хлоробактен были обнаружены в наибольшем количестве на высоте более 6 м, в то время как зеленые серные бактерии, производящие изорениератен были преимущественно идентифицированы выше 17 м. Обнаружение любого из этих каротиноидов в древних породах может ограничить глубину кислородно-кислородный переход а также ограничить прошлое экология. Окенан и хлорбактан, обнаруженные в образцах палеопротерозоя Австралии, позволили сделать вывод о временно неглубоком аноксическом переходе, вероятно, между 12 и 25 м.[2]
Окенон синтезируется у 12 видов Chromatiaceae, включая восемь. роды. Другие пурпурные серные бактерии имеют ациклические каротиноидные пигменты, такие как ликопин и Родопин. Однако, геохимики в основном изучают окенон, потому что он структурно уникален. Это единственный пигмент с 2,3,4 триметилом.арил шаблон замещения. Напротив, зеленые серные бактерии производят 2,3,6 триметиларил изопреноиды.[6] Синтез этих структур производит биологическую специфичность, которая может различать экологию окружающей среды прошлого. Окенон, хлоробактен и изорениератен производятся серобактериями путем модификации ликопин. В океноне концевая группа ликопина образует χ-кольцо, а хлоробактен имеет φ-кольцо.[7] Первый шаг в биосинтезе этих двух пигментов похож, образование β-кольца β-циклазой. фермент. Затем синтезы расходятся с каротин десатуразой /метилтрансфераза фермент, преобразующий концевую группу β-кольца в χ-кольцо. Другие реакции завершают синтез до окенона: удлинение конъюгации, добавление метокси группа и вставив кетон. Однако биологически хорошо охарактеризованы только первые стадии синтеза.
Сохранение
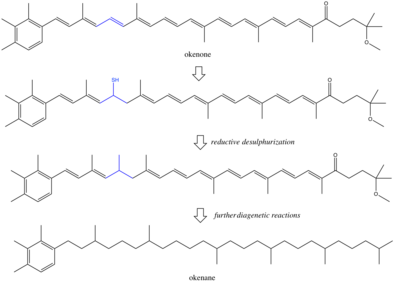
Пигменты и прочее биомаркеры продуцируемые организмами, могут ускользать от микробных и химических деградация и упорствовать в осадочные породы.[8] В условиях хранения окружающая среда часто бывает бескислородной и восстанавливающей, что приводит к химической потере функциональных групп, таких как двойные связи и гидроксильные группы. Точные реакции во время диагенез плохо изучены, хотя некоторые предложили редуктивные десульфуризация как механизм для насыщение окенона в окенан.[9][10] Всегда существует вероятность того, что окенан образуется в результате абиотических реакций, возможно, из-за метильных сдвигов в β-каротин.[11] Если бы эта реакция происходила, окенан имел бы несколько предшественников, и биологическая специфичность биомаркера снизилась бы. Однако маловероятно, что изомерные перегруппировки двух метильных групп происходят без ферментативной активности. Большинство исследований делают вывод, что окенан является истинным биомаркером пурпурных серных бактерий. Однако есть и другие биологические аргументы против этой интерпретации.[12] Прежние организмы, которые синтезировали окенон, могут не быть современными аналогами пурпурных серных бактерий. В современном океане могут быть и другие не охарактеризованные фотосинтезаторы, производящие окенон. Еще одно осложнение: горизонтальный перенос генов.[13] Если Chromatiaceae получили способность создавать окенон совсем недавно, чем в палеопротерозое, то окенан отслеживает не пурпурные серные бактерии, а скорее первоначального донора гена. Эти неоднозначности указывают на то, что интерпретация биомаркеров в породах возрастом в миллиард лет будет ограничиваться пониманием древних метаболизм.
Методы измерения
ГХ / МС
Перед анализом осадочные породы находятся извлеченный за органическая материя. Как правило, извлекается менее одного процента из-за термическая зрелость материнской породы. Органическое содержимое часто разделяют на насыщает, ароматика, и поляры. Газовая хроматография может быть соединен с масс-спектрометрии для анализа экстрагированной ароматической фракции. Соединения элюируются из столбец на основе их отношение массы к заряду (M / Z) и отображаются в зависимости от относительной интенсивности. Пики присваиваются соединениям на основе поиска в библиотеке, стандартов и относительных время удерживания. Некоторые молекулы имеют характерные пики, которые позволяют легко искать при определенном соотношении массы к заряду. Для триметиларилизопреноида окенана этот характерный пик имеет место при M / Z, равном 134.
Изотопные отношения
Соотношения изотопов углерода пурпурных и зеленых серных бактерий значительно отличаются от других фотосинтезирующих организмов. Биомасса пурпурных серных бактерий Chromatiaceae часто обеднена δ13C по сравнению с типичными кислородными фототрофами, в то время как зеленые серные бактерии, Chlorobiaceae, часто бывают обогащены.[14] Это предлагает дополнительную дискриминацию для определения экологических сообществ, сохранившихся в осадочных породах. Для биомаркера окенана δ13C может быть определен Масс-спектрометр изотопного отношения.
Пример использования: Северная Австралия
В современной среде обитают пурпурные серные бактерии. меромиктический (постоянно стратифицированные) озера[15] и Silled Fjords и встречаются в нескольких морских экосистемах. Гиперсоленый воды как Черное море исключения.[16] Однако миллиарды лет назад, когда океаны были бескислородными и сульфидными, у фототрофных серных бактерий было больше места для обитания. Исследователи из Австралийский национальный университет и Массачусетский Институт Технологий исследовали породы возрастом 1,6 миллиарда лет, чтобы изучить химические условия Палеопротерозой океан. Многие считают, что на этот раз из-за исчезновения проникающих столбов кислородной воды полосчатые железные образования примерно 1,8 миллиарда лет назад. Другие, возглавляемые Дональд Кэнфилд 1998 год Природа бумага, считают, что воды были эвксинными. Изучение горных пород того времени выявило биомаркеры как пурпурных, так и зеленых серных бактерий, добавив доказательства, подтверждающие Canfield Ocean гипотеза. Проанализированным обнажением осадочных пород была формация Барни Крик из группы МакАртур в северной Австралии. Анализ образцов выявил как 2,3,6-триметиларлизопреноиды (хлорбактан) Chlorobiaceae, так и 2,3,4-триметиларилизопреноиды (окенан) Chromatiaceae. И хлорбактан, и окенан указывают на эвксиновый океан, с сульфидными и бескислородными условиями на поверхности ниже 12-25 м. Авторы пришли к выводу, что хотя кислород был в атмосфере, палеопротерозойские океаны не были полностью насыщены кислородом.[2]
Смотрите также
- Аноксическое событие
- Аноксигенный фотосинтез
- Биомаркеры
- Каротиноиды
- Зеленые серные бактерии
- Пурпурные серные бактерии
Рекомендации
- ^ Имхофф, Йоханнес Ф. (1 января 1995 г.). "Таксономия и физиология фототрофных пурпурных бактерий и зеленых серных бактерий". В Бланкеншипе Роберт Э .; Мэдиган, Майкл Т .; Бауэр, Карл Э. (ред.). Аноксигенные фотосинтетические бактерии. Достижения в фотосинтезе и дыхании. 2. Springer Нидерланды. С. 1–15. Дои:10.1007/0-306-47954-0_1. ISBN 9780792336815.
- ^ а б c Brocks, Jochen J .; Шеффер, Филипп (2008-03-01). «Окенан, биомаркер пурпурных серных бактерий (Chromatiaceae) и других новых производных каротиноидов из формации Барни-Крик 1640 млн лет». Geochimica et Cosmochimica Acta. 72 (5): 1396–1414. Дои:10.1016 / j.gca.2007.12.006.
- ^ Brocks, Jochen J .; С любовью, Гордон Д .; Вызывает, Роджер Э .; Knoll, Andrew H .; Логан, Грэм А .; Боуден, Стивен А. (2005). «Биомаркеры зеленых и пурпурных серных бактерий в стратифицированном палеопротерозойском море». Природа. 437 (7060): 866–870. Дои:10.1038 / природа04068. PMID 16208367.
- ^ Шеффер, Филипп; Адам, Пьер; Верунг, Патрик; Альбрехт, Пьер (1997-12-01). «Новые производные ароматических каротиноидов из фотосинтетических бактерий серы в отложениях». Буквы Тетраэдра. 38 (48): 8413–8416. Дои:10.1016 / S0040-4039 (97) 10235-0.
- ^ Гемерден, Ханс Ван; Мас, Хорди (1 января 1995 г.). Бланкеншип, Роберт Э .; Мэдиган, Майкл Т .; Бауэр, Карл Э. (ред.). Аноксигенные фотосинтетические бактерии. Достижения в фотосинтезе и дыхании. Springer Нидерланды. С. 49–85. Дои:10.1007/0-306-47954-0_4. ISBN 9780792336815.
- ^ Summons, R.E .; Пауэлл, Т. Г. (1987-03-01). «Идентификация арилизопреноидов в материнских породах и сырой нефти: биологические маркеры зеленых серных бактерий». Geochimica et Cosmochimica Acta. 51 (3): 557–566. Дои:10.1016 / 0016-7037 (87) 90069-X.
- ^ Vogl, K .; Брайант, Д. А. (2012-05-01). «Биосинтез биомаркера окенона: образование χ-кольца». Геобиология. 10 (3): 205–215. Дои:10.1111 / j.1472-4669.2011.00297.x. ISSN 1472-4669. PMID 22070388.
- ^ Brocks, Jochen J .; Грайс, Клити (01.01.2011). Рейтнер, Иоахим; Тиль, Фолькер (ред.). Энциклопедия геобиологии. Энциклопедия серии наук о Земле. Springer Нидерланды. С. 147–167. Дои:10.1007/978-1-4020-9212-1_30. ISBN 9781402092114.
- ^ Hebting, Y .; Schaeffer, P .; Behrens, A .; Adam, P .; Schmitt, G .; Schneckenburger, P .; Бернаскони, С. М .; Альбрехт, П. (16.06.2006). «Свидетельства биомаркеров основного пути сохранения осадочного органического углерода». Наука. 312 (5780): 1627–1631. Дои:10.1126 / science.1126372. ISSN 0036-8075. PMID 16690819.
- ^ Werne, Josef P .; Лайонс, Тимоти У .; Холландер, Дэвид Дж .; Схоутен, Стефан; Hopmans, Ellen C .; Синнингхе Дамсте, Яап С. (15 июля 2008 г.). «Изучение путей диагенетической сульфуризации органических веществ с использованием изотопного анализа серы, специфичного для соединений». Geochimica et Cosmochimica Acta. 72 (14): 3489–3502. Дои:10.1016 / j.gca.2008.04.033.
- ^ Купманс, Мартин П .; Схоутен, Стефан; Kohnen, Math E.L .; Синнингхе Дамсте, Яап С. (1996-12-01). «Ограниченное применение арилизопреноидов в качестве индикаторов аноксии фотической зоны». Geochimica et Cosmochimica Acta. 60 (23): 4873–4876. Дои:10.1016 / S0016-7037 (96) 00303-1. HDL:1874/4427.
- ^ Brocks, Jochen J .; Банфилд, Джиллиан (2009). «Раскрытие древней микробной истории с помощью протеогеномики сообщества и геохимии липидов». Обзоры природы Микробиология. 7 (8): 601–609. Дои:10.1038 / nrmicro2167. PMID 19609261.
- ^ Коббс, Кэссиди; Хит, Джереми; Stireman III, John O .; Аббат, Патрик (1 августа 2013 г.). «Каротиноиды в неожиданных местах: галлицы, латеральный перенос генов и биосинтез каротиноидов у животных». Молекулярная филогенетика и эволюция. 68 (2): 221–228. Дои:10.1016 / j.ympev.2013.03.012. PMID 23542649.
- ^ Зякун, А. М .; Лунина, О. Н .; Прусакова, Т. С .; Пименов, Н. В .; Иванов, М. В. (06.12.2009). «Фракционирование стабильных изотопов углерода фотоавтотрофно растущими аноксигенными пурпурными и зелеными серными бактериями». Микробиология. 78 (6): 757. Дои:10.1134 / S0026261709060137. ISSN 0026-2617.
- ^ Оверманн, Йорг; Битти, Дж. Томас; Холл, Кен Дж .; Пфенниг, Норберт; Норткот, Том Г. (1991-07-01). «Характеристика плотного пурпурного бактериального слоя серы в меромиктовом соленом озере» (PDF). Лимнология и океанография. 36 (5): 846–859. Дои:10.4319 / lo.1991.36.5.0846. ISSN 1939-5590.
- ^ Hashwa, F.A .; Трупер, Х. Г. (1978). «Жизнеспособные фототрофные серные бактерии со дна Черного моря». Helgoländer Wissenschaftliche Meeresuntersuchungen. 31 (1–2): 249–253. Дои:10.1007 / BF02297000. ISSN 0017-9957.