Терапевтическая генная модуляция - Therapeutic gene modulation
Терапевтическая генная модуляция относится к практике изменения экспрессия гена на одной из различных стадий с целью облегчения какой-либо формы недуга. Он отличается от генная терапия при этом модуляция гена направлена на изменение экспрессии эндогенного гена (возможно, путем введения гена, кодирующего новый модулирующий белок), тогда как генная терапия касается введения гена, продукт которого напрямую помогает реципиенту.
Модуляция экспрессии генов может быть опосредована на уровне транскрипция ДНК-связывающими агентами (которые могут быть факторы искусственной транскрипции ), маленькие молекулы, или же синтетические олигонуклеотиды. Он также может опосредоваться посттранскрипционно через РНК-интерференция.
Модуляция транскрипционного гена
Подход к терапевтической модуляции использует агенты, которые модулируют эндогенную транскрипцию, специфически воздействуя на эти гены в гДНК уровень. Преимущество этого подхода перед модуляцией на уровне мРНК или белка состоит в том, что каждая клетка содержит только одну копию гДНК. Таким образом, количество копий-мишеней значительно ниже, что позволяет теоретически вводить лекарства в гораздо более низких дозах.
Этот подход также предлагает несколько преимуществ по сравнению с традиционными генная терапия. Прямой таргетинг эндогенный транскрипция должна давать правильное относительное выражение сращивание варианты. Напротив, традиционная генная терапия обычно вводит ген, который может экспрессировать только один транскрипт, а не набор стехиометрически экспрессируемых вариантов сплайсированных транскриптов. Кроме того, введенные вирусом гены могут быть нацелены на подавление генов путем метилирования, что может противодействовать эффекту традиционной генной терапии.[1] Ожидается, что это не будет проблемой для модуляции транскрипции, поскольку она действует на эндогенную ДНК.
Существует три основные категории агентов, которые действуют как модуляторы транскрипционных генов: триплексообразующие олигонуклеотиды (TFO), синтетические полиамиды (SPA) и ДНК-связывающие белки.[2]
Триплекс-образующие олигонуклеотиды
Кто они такие
Образующие триплекс олигонуклеотиды (TFO) являются одним из возможных способов достижения терапевтической модуляции генов. TFO имеют длину примерно 10-40 пар оснований и могут связываться с большая канавка в дуплексной ДНК, которая образует третью цепь или тройную спираль.[2][3] Связывание происходит в областях полипурина или полипиримидина через водородные связи Хугстина с пуриновыми (A / G) основаниями на двухцепочечной ДНК, которая уже находится в форме Спираль Уотсона-Крика.[4]
Как они работают
TFO могут быть молекулами полипурина или полипиримидина и связываться с одной из двух цепей двойной спирали либо параллельно, либо параллельно. антипараллельный ориентация на участки полипурина или полипиримидина. Поскольку коды распознавания ДНК различны для параллельного и антипараллельного способа связывания TFO, TFO, состоящие из пиримидины (C / T) связываются с богатой пуринами цепью целевой двойной спирали через Hoogsteen водородные связи параллельно.[3] TFO в составе пурины (A / G) или смешанный пурин и пиримидин связываются с одной и той же богатой пурином цепью через обратные связи Хугстина антипараллельным образом. TFO могут распознавать богатые пуринами целевые цепи для дуплексной ДНК.[2]

Осложнения и ограничения
Чтобы мотивы TFO связались параллельным образом и создали водородные связи, атом азота в положении 3 остатка цитозина должен быть протонированный, но на физиологический pH уровней это не так, что может предотвратить параллельное связывание.[2]
Другое ограничение состоит в том, что TFO могут связываться только с богатыми пуринами целевыми цепями, и это ограничивает выбор эндогенный сайты-мишени генов для полипурин-полипиримидиновых участков в дуплексной ДНК. Если бы был сгенерирован метод, позволяющий также связать TFO с пиримидиновыми основаниями, это позволило бы TFO нацеливаться на любую часть геном. Также геном человека богат последовательностями полипурина и полипиримидина, которые могут влиять на специфичность TFO для связывания с целевой областью ДНК. Подход к преодолению этого ограничения заключается в разработке TFO с модифицированными нуклеотидами, которые действуют как заблокированные нуклеиновые кислоты увеличить близость TFO для конкретных целевых последовательностей.[5]
Другие ограничения включают опасения относительно связывающая аффинность и специфичность, стабильность in vivo и проникновение в клетки. Исследователи пытаются преодолеть эти ограничения, улучшая характеристики TFO за счет химические модификации, например, изменение магистрали TFO для уменьшения электростатическое отталкивание между TFO и дуплексом ДНК. Кроме того, из-за их высокой молекулярной массы поглощение клетками ограничено, и некоторые стратегии для преодоления этого включают Конденсирующие агенты ДНК, связывание TFO с гидрофобными остатками, такими как холестерин или агенты, повышающие проницаемость клеток.[2]
Что они могут сделать
Ученые все еще совершенствуют технологию, чтобы превратить TFO в терапевтический продукт и большая часть этого вращается вокруг их потенциального применения в антигенной терапии. В частности, они использовались как индукторы сайт-специфические мутации, реагенты, которые избирательно и специфично раскалывать ДНК-мишень, и как модуляторы экспрессия гена.[6] Один из таких методов модификации последовательности гена заключается в нацеливании ДНК с помощью TFO для активации целевой ген. Если целевая последовательность расположена между двумя неактивными копиями гена, лиганды ДНК, такие как TFO, могут связываться с целевым сайтом и распознаваться как повреждения ДНК. Чтобы исправить эти поражения, Ремонт ДНК комплексы собираются на целевой последовательности, ДНК восстанавливается. Повреждение субстрата внутримолекулярной рекомбинации затем может быть восстановлено и обнаружено, если резекция заходит достаточно далеко, чтобы получить совместимые концы с обеих сторон от сайта расщепления, а затем лигируют 3'-выступы, что приводит к образованию единственной активной копии гена и потере всех последовательностей между двумя копиями гена.[4]
В модельных системах TFO могут ингибировать экспрессию генов на уровне ДНК, а также вызывать целевой мутагенез в модели.[6] Индуцированное ТФО ингибирование удлинения транскрипции на эндогенных мишенях было успешно протестировано на культурах клеток.[7] Однако, несмотря на многое in vitro Успех в сотовых приложениях был ограничен, возможно, из-за целевой доступности.
TFO могут заставить замолчать ген молчания путем нацеливания на инициацию или удлинение транскрипции, остановку на сайтах связывания триплекса или внесение постоянных изменений в последовательность-мишень посредством стимуляции собственных путей репарации клетки. Эти приложения могут быть актуальны при создании лечение рака которые подавляют экспрессию генов на уровне ДНК. Поскольку аберрантная экспрессия генов является признаком рака, регулирование уровней экспрессии этих эндогенных генов может потенциально действовать как терапия для нескольких рак типы.
Синтетические полиамиды
Синтетические полиамиды представляют собой набор небольших молекул, которые образуют специфические водородные связи с малая бороздка ДНК. Они могут оказывать действие либо напрямую, связывая регуляторную область или транскрибируемую область гена для модификации транскрипции, либо косвенно, путем спроектированной конъюгации с другим агентом, который вносит изменения в целевой сайт ДНК.
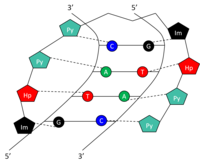
Структура
Определенные основания в малой бороздке ДНК могут распознаваться и связываться небольшими синтетическими полиамидами (SPA). ДНК-связывающие СПА содержат три полиамидно-аминокислотных компонента: гидроксипиррол (Hp), имидазол (Im), и пиррол (Py).[10] Цепочки этих аминокислот образуют петлю в виде шпильки. Аминокислоты по обе стороны от шпильки образуют пару, которая может распознавать обе стороны шпильки. Базовая пара Уотсона-Крика. Это происходит за счет водородных связей внутри малой бороздки ДНК. Амидные пары Py / Im, Py / Hp, Hp / Py и Im / Py распознают пары оснований Уотсона-Крика C-G, A-T, T-A и G-C, соответственно (таблица 1). См. Рисунок для графического представления распознавания 5'-GTAC-3 'SPA. СПА обладают низкой токсичностью, но еще не использовались для модуляции генов человека.
| Амидная пара | Нуклеотидная пара |
|---|---|
| Py / Im | C-G |
| Py / Hp | В |
| Л.с. / Py | Т-А |
| Im / Py | G-C |
Ограничения и обходные пути
Главный структурный недостаток немодифицированных SPA как модуляторов генов состоит в том, что их последовательность распознавания не может быть расширена за пределы 5 пар оснований Уотсона-Крика. Естественная кривизна малой бороздки ДНК слишком крута для структуры шпильки. Есть несколько групп, предлагающих обходные пути к этой проблеме.[8][11][12][13][14] SPA можно сделать так, чтобы оно лучше соответствовало кривизне малой канавки, вставив бета-аланин который расслабляет структуру.[10] Другой подход к увеличению длины распознавания - последовательное использование нескольких коротких шпилек.[15][16] Этот подход увеличил длину распознавания до одиннадцати пар оснований Watson-Crick.


Прямая модуляция
SPA могут ингибировать транскрипцию за счет связывания в транскрибируемой области гена-мишени. Это ингибирование происходит за счет блокирования удлинения РНК-полимеразой.
SPA также могут модулировать транскрипцию, воздействуя на сайт связывания регулятора транскрипции. Если регулятор является активатором транскрипции, это снизит уровни транскрипции. В качестве примера было продемонстрировано, что нацеливание SPA на сайт связывания для активирующего фактора транскрипции TFIIIA ингибирует транскрипцию расположенной ниже 5S РНК.[17] Напротив, если регулятор является репрессором, это повысит уровни транскрипции. В качестве примера, нацеливание SPA на фактор хозяина LSF, который подавляет экспрессию длинного концевого повтора (LTR) вируса иммунодефицита человека (ВИЧ) типа 1, блокирует связывание LSF и, следовательно, подавляет экспрессию LTR.[18].

Конъюгированная модуляция
Не было показано, что СПА напрямую модифицируют ДНК или обладают другой активностью, кроме прямого блокирования других факторов или процессов. Однако модифицирующие агенты могут быть связаны с хвостовыми концами конструкции шпильки. Специфическое связывание SPA с ДНК обеспечивает сайт-специфическое нацеливание конъюгированного модифицирующего агента.
SPA были объединены с ДНК-алкилирующими фрагментами циклопропилпирролоиндола.[19] и хлорамбуцил[20] которые были способны повредить и сшить ДНК SV40. Этот эффект ингибировал клеточный цикл и рост. Хлорамбуцил, химиотерапевтический агент, был более эффективен в сочетании с SPA, чем без него.
В 2012 году SPA были конъюгированы с SAHA, мощным ингибитором гистондеацетилазы (HDAC).[21] SPA с конъюгированными SAHA были нацелены на Oct-3/4 и Nanog, которые вызывали эпигенетическое ремоделирование и, следовательно, увеличивали экспрессию множественных генов, связанных с плюрипотентностью, в эмбриональных фибробластах мыши.
Дизайнерские протеины с цинковыми пальцами
Какие они есть / структура
Созданы дизайнерские протеины с цинковыми пальцами белки используется для нацеливания на определенные области ДНК. Эти белки используют Связывание с ДНК емкость природных цинковый палец домены для модуляции определенных целевых областей геном.[22] Как в авторском, так и в натуральном мотиве «цинковые пальцы» белок состоит из двух β-листы и один α-спираль. Два гистидин остатки на α-спирали и два цистеин остатки на β-листах связаны с цинк атом, который служит для стабилизации белкового домена в целом. Эта стабилизация особенно полезна для α-спирали в ее функции как домена распознавания и связывания ДНК. Фактор транскрипции TFIIIA является примером встречающегося в природе белка с мотивами «цинковые пальцы».[23]

Как они работают
Мотивы цинковых пальцев встраиваются в большая канавка спиральной ДНК,[23] где аминокислота остаток последовательность на α-спирали придает мотиву специфичность его последовательности-мишени. Домен привязан к семи-нуклеотид последовательность ДНК (позиции с 1 по 6 на первичная прядь ДНК, плюс позиции 0 и 3 на дополнительная нить ), тем самым обеспечивая высокую селективность белкового мотива по отношению к своей мишени.[22] При разработке дизайнерского белка из цинковых пальцев исследователи могут использовать такие методы, как сайт-направленный мутагенез с последующим рандомизированные испытания на связующую способность,[22][24] или рекомбинация in vitro мотивов с известной целевой специфичностью для получения библиотеки конечных белков, специфичных для последовательности.[25]


Эффекты и воздействия на модуляцию генов
Дизайнерские белки с цинковыми пальцами могут модулировать экспрессия генома несколькими способами. В конечном итоге за конечный результат экспрессии в первую очередь отвечают два фактора: является ли целевая последовательность регулирующий регион или кодирующая область ДНК, и какие типы эффекторные домены связаны с доменом цинкового пальца. Если целевая последовательность для инженерного дизайнерский протеин является регуляторной областью - например, промоутер или репрессор из репликация - сайт связывания для естественных факторов транскрипции будет скрыт, что приведет к соответствующему уменьшению или увеличению, соответственно, транскрипция для связанных ген.[26] Точно так же, если целевая последовательность является экзон, дизайнерский цинковый палец заслонит последовательность от РНК-полимераза комплексы транскрипции, приводящие к усеченному или нефункциональному продукту гена.[22]
Эффекторные домены, связанные с цинковым пальцем, также могут иметь сопоставимые эффекты. Это функция этих эффекторных доменов, которые, возможно, являются наиболее важными в отношении использования дизайнерских белков «цинковые пальцы» для терапевтической модуляции генов. Если метилаза домен связывается с дизайнерским белком цинкового пальца, когда белок цинкового пальца связывается с целевой последовательностью ДНК, увеличение состояние метилирования ДНК в этом регионе впоследствии будет результатом. Скорость транскрипции пораженных таким образом генов будет снижена.[27] Многие из эффекторных доменов функционируют, чтобы напрямую модулировать ДНК - например, через метилирование, расщепление,[28] или рекомбинация целевой последовательности ДНК[29] - или изменяя скорость его транскрипции - например, ингибирование транскрипции через репрессорные домены, которые блокируют транскрипционный аппарат,[30] продвижение транскрипции с помощью доменов активации, которые привлекают на сайт транскрипционный аппарат,[31] или же гистон - или другой эпигенетический -модификации доменов, влияющих на хроматин состояние и способность транскрипционного аппарата получать доступ к затронутым генам.[32] Эпигенетическая модификация является основной темой при определении различных уровней экспрессии генов, что объясняется идеей о том, насколько плотно запутана цепь ДНК - от гистонов на локальном уровне до хроматина на локальном уровне. хромосомный уровень - может влиять на доступность последовательностей ДНК для аппарата транскрипции, тем самым влияя на скорость, с которой она может быть транскрибирована.[23] Если вместо прямого воздействия на нить ДНК, как описано выше, дизайнерский белок с цинковыми пальцами вместо этого влияет на состояние эпигенетической модификации для целевой области ДНК, аналогичным образом может осуществляться модуляция экспрессии гена.
В первом случае успешно продемонстрировать использование дизайнерских белков «цинковые пальцы» для модуляции экспрессии генов. in vivo, Чу и другие[26] разработали белок, состоящий из трех доменов с цинковыми пальцами, которые нацелены на конкретную последовательность на BCR-ABL слияние онкоген. Этот специфический онкоген участвует в острый лимфобластный лейкоз. Онкоген обычно позволяет лейкемия клетки размножаться в отсутствие специфических факторов роста, что является признаком рак. Включив сигнал ядерной локализации с трехдоменным белком «цинковые пальцы» для облегчения связывания белка с геномной ДНК в ядре, Choo и другие смогли продемонстрировать, что созданный ими белок может блокировать транскрипцию онкогена in vivo. Клетки лейкемии стали зависимыми от обычных факторов роста, вернув клеточный цикл под контроль нормальное регулирование.[26]
Посттранскрипционная модуляция генов
Основной подход к посттранскрипционной модуляции генов - через РНК-интерференция (РНКи). Основная проблема с использованием РНКи в модуляции генов - это доставка лекарства к клеткам-мишеням.[33][34] Модуляция гена РНКи была успешно применена к мышам для лечения воспалительного заболевания кишечника на мышиной модели.[35] В этом лечении использовались стабилизированные наночастицы, нацеленные на интегрин бета-7, на основе липосом, захватывающие короткие интерферирующие РНК (миРНК). Существует несколько других форм доставки РНКи, в том числе: доставка полиплекса, конъюгаты лиганд-миРНК, «голая» доставка, доставка неорганических частиц с использованием наночастиц золота и сайт-специфическая локальная доставка.[36]
Клиническое значение
С другой стороны, дизайнерские белки с цинковыми пальцами прошли некоторые испытания в клиническая арена. Эффективность и безопасность EW-A-401, сконструированного фактора транскрипции цинкового пальца, в качестве фармакологическое средство для лечения хромота, а сердечно-сосудистый недуг, был исследован в клинических испытаниях.[37] Белок состоит из модифицированного плазмидная ДНК который побуждает пациента продуцировать сконструированный фактор транскрипции, мишенью которого является фактор роста эндотелия сосудов-A (VEGF-A) ген, положительно влияющий кровеносный сосуд разработка. Хотя еще не одобрено США. Управление по контролю за продуктами и лекарствами (FDA), два Клинические исследования фазы I были завершены, которые идентифицируют этот белок цинкового пальца как многообещающий и безопасный потенциальный терапевтический агент для лечения заболевание периферических артерий в людях.[38]
Смотрите также
Рекомендации
- ^ Молодой WB, Link CJ (2000). «Химерный ретровирусный вирус-помощник и последовательность IRES пикорнавируса для устранения метилирования ДНК для улучшения ретровирусной упаковки клеток». Журнал вирусологии. 74 (11): 5242–5249. Дои:10.1128 / JVI.74.11.5242-5249.2000. ЧВК 110878. PMID 10799600.
- ^ а б c d е Уил Т.Г., Хайма Х.Дж., Ротс М.Г. (2003). «Терапевтическая модуляция функции эндогенных генов с помощью агентов с заданной специфичностью последовательности ДНК». Исследования нуклеиновых кислот. 31 (21): 6064–6078. Дои:10.1093 / нар / gkg815. ЧВК 275457. PMID 14576293.
- ^ а б Sargent, RG .; Kim, S .; Грюнерт, округ Колумбия. (2011). «Модификация генов на основе олиго / полинуклеотидов: стратегии и терапевтический потенциал». Олигонуклеотиды. 21 (2): 55–75. Дои:10.1089 / oli.2010.0273. ЧВК 3078494. PMID 21417933.
- ^ а б Саймон, П .; Cannata, F .; Concordet, JP; Джованнанджели, К. (август 2008 г.). «Нацеливание ДНК с образующими триплекс олигонуклеотидами для модификации последовательности гена». Биохимия. 90 (8): 1109–16. Дои:10.1016 / j.biochi.2008.04.004. PMID 18460344.
- ^ Zhou, Y .; Kierzek, E .; Лоо, ЗП .; Антонио, М .; Яу, YH .; Чуах, YW .; Гейфман-Шохат, С .; Kierzek, R .; Чен, Г. (июль 2013 г.). «Распознавание дуплексов РНК химически модифицированными триплексообразующими олигонуклеотидами». Нуклеиновые кислоты Res. 41 (13): 6664–73. Дои:10.1093 / nar / gkt352. ЧВК 3711454. PMID 23658228.
- ^ а б Guntaka, RV .; Варма, BR .; Вебер, К.Т. (Январь 2003 г.). «Триплекс-образующие олигонуклеотиды как модуляторы экспрессии генов». Int J Biochem Cell Biol. 35 (1): 22–31. Дои:10.1016 / с 1357-2725 (02) 00165-6. PMID 12467644.
- ^ Faria, M .; Дерево, CD .; Perrouault, L .; Nelson, JS .; Winter, A .; White, MR .; Helene, C .; Джованнанджели, К. (апрель 2000 г.). «Направленное ингибирование удлинения транскрипции в клетках, опосредованное триплекс-образующими олигонуклеотидами». Proc Natl Acad Sci U S A. 97 (8): 3862–7. Дои:10.1073 / pnas.97.8.3862. ЧВК 18107. PMID 10760257.
- ^ а б Редди Б.С., Шарма С.К., Лоун Д.В. (2001). «Последние разработки в области селективных эффекторов ДНК малых бороздок». Curr. Med. Chem. 8 (5): 475–508. Дои:10.2174/0929867003373292. PMID 11281837.
- ^ Дерван ПБ (2001). «Молекулярное распознавание ДНК малыми молекулами». Биоорг. Med. Chem. 9 (9): 2215–2235. Дои:10.1016 / s0968-0896 (01) 00262-0. PMID 11553460.
- ^ а б White S, Szewcxyk JW, Turner JM, Baird EE, Dervan PB (1998). «Распознавание четырех пар оснований Уотсона-Крика в малой бороздке ДНК синтетическими лигандами» (PDF). Природа. 391 (6666): 468–471. Дои:10.1038/35106. PMID 9461213. S2CID 205023593.
- ^ Дерван ПБ, Эдельсон Б.С. (2003). «Распознавание малой бороздки ДНК пирролимидазольными полиамидами». Curr. Мнение. Struct. Биол. 13 (3): 284–299. Дои:10.1016 / s0959-440x (03) 00081-2. PMID 12831879.
- ^ Лаун Дж. В. (1988). «Лекситропсины: рациональный дизайн агентов чтения последовательности ДНК в качестве новых противораковых агентов и потенциальных клеточных зондов». Противораковый препарат Des. 3 (1): 25–40. PMID 2838035.
- ^ Траугер Дж. У., Бэрд Э., Дерван ПБ (1996). «Распознавание ДНК сконструированными лигандами при субнаномолярных концентрациях». Природа. 382 (6591): 559–561. Дои:10.1038 / 382559a0. PMID 8700233. S2CID 4335955.
- ^ Wemmer DE (2000). «Разработаны лиганды малых бороздок, специфичные для последовательности». Анну. Rev. Biophys. Biomol. Struct. 29: 439–461. Дои:10.1146 / annurev.biophys.29.1.439. PMID 10940255.
- ^ Керс I, Дерван ПБ (2002). «Поиск оптимального линкера в полиамидах шпильки тандемного типа». Биоорг. Med. Chem. 10 (10): 3339–3349. Дои:10.1016 / s0968-0896 (02) 00221-3. PMID 12150881.
- ^ Вейерманн П., Дерван ПБ (2002). «Распознавание десяти пар оснований ДНК с помощью димеров шпильки голова к голове» (PDF). Варенье. Chem. Soc. 124 (24): 6872–6878. Дои:10.1021 / ja020258k. PMID 12059208.
- ^ Gottesfeld JM, Neely L, Trauger JW, Baird EE, Dervan PB (1997). «Регуляция экспрессии генов небольшими молекулами». Природа. 387 (6629): 202–205. Дои:10.1038 / 387202a0. PMID 9144294. S2CID 4358491.
- ^ Coull JJ, He G, Melander C, Rucker VC, Dervan PB, Margolis DM (2002). «Направленная дерепрессия длинного концевого повтора вируса иммунодефицита человека 1 типа пирролимидазольными полиамидами». Дж. Вирол. 76 (23): 12349–12354. Дои:10.1128 / jvi.76.23.12349-12354.2002. ЧВК 136904. PMID 12414976.
- ^ Ван Ю.Д., Дзигелевски Дж., Вюрц Н.Р., Дзигелевска Б., Дерван ПБ, Берман Т.А. (2003). «Сшивание ДНК и биологическая активность шпильки полиамид-хлорамбуцил конъюгата». Исследования нуклеиновых кислот. 31 (21): 1208–1215. Дои:10,1093 / нар / гкг215. ЧВК 150233. PMID 12582240.
- ^ Ван Ю.Д., Дзигелевски Дж., Чанг А.Ю., Дерван П.Б., Бирман Т.А. (2002). «Внеклеточная и клеточная активность конъюгата полиамид-CBI, селективного по последовательности ДНК». J. Biol. Chem. 277 (45): 42431–42437. Дои:10.1074 / jbc.M207179200. PMID 12196541.
- ^ Пандиан Н.Г., Накано Й., Сато С., Моринага Х., Бандо Т, Нагасе Х., Сугияма Х. (2012). «Синтетическая малая молекула для быстрой индукции множественных генов плюрипотентности в эмбриональных фибробластах мыши». Научные отчеты. 2 (544): 544. Дои:10.1038 / srep00544. ЧВК 3408130. PMID 22848790.
- ^ а б c d Papworth, M .; Колашинская, П .; Минчук, М. (январь 2006 г.). «Дизайнерские протеины с цинковыми пальцами и их применение». Ген. 366 (1): 27–38. Дои:10.1016 / j.gene.2005.09.011. PMID 16298089.
- ^ а б c Уотсон, Джеймс Д. (2008). Молекулярная биология гена. Сан-Франциско: Пирсон / Бенджамин Каммингс. п. 595. ISBN 978-0-8053-9592-1.
- ^ Desjarlais, JR .; Берг, JM. (Март 1993 г.). «Использование структуры консенсусной последовательности цинкового пальца и правил специфичности для разработки специфических ДНК-связывающих белков». Proc Natl Acad Sci U S A. 90 (6): 2256–60. Дои:10.1073 / пнас.90.6.2256. ЧВК 46065. PMID 8460130.
- ^ Исалан, М .; Klug, A .; Чу, Ю. (июль 2001 г.). «Быстрый, общеприменимый метод создания цинковых пальцев, иллюстрируемый нацеливанием на промотор ВИЧ-1». Nat Biotechnol. 19 (7): 656–60. Дои:10.1038/90264. ЧВК 2677679. PMID 11433278.
- ^ а б c Чу, Й .; Sánchez-García, I .; Клуг, А. (декабрь 1994 г.). «Репрессия in vivo сайт-специфическим ДНК-связывающим белком, разработанным против онкогенной последовательности» (PDF). Природа. 372 (6507): 642–5. Дои:10.1038 / 372642a0. HDL:10261/6295. PMID 7990954. S2CID 12701336.
- ^ Карвин, CD .; Parr, RD .; Кладде, депутат. (Ноябрь 2003 г.). «Сайт-селективное нацеливание in vivo на метилирование ДНК цитозина-5 белками цинкового пальца». Нуклеиновые кислоты Res. 31 (22): 6493–501. Дои:10.1093 / нар / gkg853. ЧВК 275549. PMID 14602907.
- ^ Kim, YG .; Cha, J .; Чандрасегаран, С. (февраль 1996 г.). «Гибридные рестрикционные ферменты: слияния цинковых пальцев с доменом расщепления Fok I». Proc Natl Acad Sci U S A. 93 (3): 1156–60. Дои:10.1073 / пнас.93.3.1156. ЧВК 40048. PMID 8577732.
- ^ Урнов, ФД .; Miller, JC .; Lee, YL .; Beausejour, CM .; Rock, JM .; Август, S .; Джеймисон, AC .; Porteus, MH .; и другие. (Июнь 2005 г.). «Высокоэффективная эндогенная коррекция генов человека с использованием разработанных нуклеаз типа« цинковые пальцы »». Природа. 435 (7042): 646–51. Дои:10.1038 / природа03556. PMID 15806097. S2CID 4390010.
- ^ Beerli, RR .; Dreier, B .; Барбас, CF. (Февраль 2000 г.). «Положительная и отрицательная регуляция эндогенных генов с помощью сконструированных факторов транскрипции». Proc Natl Acad Sci U S A. 97 (4): 1495–500. Дои:10.1073 / pnas.040552697. ЧВК 26462. PMID 10660690.
- ^ Lara, H .; Wang, Y .; Beltran, AS .; Хуарес-Морено, К .; Юань, X .; Като, S .; Leisewitz, AV .; Cuello Fredes, M .; и другие. (Август 2012 г.). «Противодействие серозному эпителиальному раку яичников с помощью дизайнерских факторов транскрипции цинкового пальца». J Biol Chem. 287 (35): 29873–86. Дои:10.1074 / jbc.M112.360768. ЧВК 3436144. PMID 22782891.
- ^ Сноуден, AW .; Zhang, L .; Урнов, Ф .; Dent, C .; Jouvenot, Y .; Чжун, X .; Rebar, EJ .; Джеймисон, AC .; и другие. (Декабрь 2003 г.). «Подавление фактора роста эндотелия сосудов А в клетках глиобластомы с использованием сконструированных факторов транскрипции цинкового пальца». Рак Res. 63 (24): 8968–76. PMID 14695215.
- ^ Бельке, Массачусетс. (Апрель 2006 г.). «Прогресс в использовании миРНК in vivo». Мол Тер. 13 (4): 644–70. Дои:10.1016 / j.ymthe.2006.01.001. ЧВК 7106286. PMID 16481219.
- ^ Dykxhoorn, DM .; Либерман, Дж. (2006). «Запуск интерференции: перспективы и препятствия для использования малых интерферирующих РНК в качестве низкомолекулярных лекарств». Анну Рев Биомед анг. 8: 377–402. CiteSeerX 10.1.1.418.758. Дои:10.1146 / annurev.bioeng.8.061505.095848. PMID 16834561.
- ^ Peer, D .; Парк, EJ .; Morishita, Y .; Карман, CV .; Симаока, М. (февраль 2008 г.). «Системная лейкоцит-направленная доставка siRNA, показывающая, что циклин D1 является противовоспалительной мишенью». Наука. 319 (5863): 627–30. Дои:10.1126 / science.1149859. ЧВК 2490797. PMID 18239128.
- ^ Rettig, GR .; Бельке, Массачусетс. (Март 2012 г.). «Прогресс в использовании миРНК-II in vivo». Мол Тер. 20 (3): 483–512. Дои:10.1038 / мт.2011.263. ЧВК 3293614. PMID 22186795.
- ^ «Идентификатор NCT00080392. Модуляция фактора роста эндотелия сосудов (VEGF) с использованием сконструированного фактора транскрипции цинкового пальца для лечения перемежающейся хромоты нижних конечностей». ClinicalTrials.gov. Национальные институты здравоохранения США. 30 декабря 2011 г.. Получено 25 июля 2013.
- ^ Giacca, M .; Закчинья, С. (июнь 2012 г.). «Генная терапия VEGF: терапевтический ангиогенез в клинике и за ее пределами». Джин Тер. 19 (6): 622–9. Дои:10.1038 / gt.2012.17. PMID 22378343.