Ботриолепис - Bothriolepis
| Ботриолепис | |
|---|---|
 | |
| Модель B. canadensis | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Класс: | †Плакодерми |
| Порядок: | †Антиархи |
| Семья: | †Ботриолепидиды |
| Род: | †Ботриолепис Эйхвальд, 1840 |
| Виды | |
| |
Ботриолепис (Греческий: «ямчатая чешуя» или «траншейная шкала») была широко распространенной, многочисленной и разнообразной род из антиарх плакодермы которые жили во время Средний к Поздний девон период Палеозойская эра. Исторически, Ботриолепис обитали во множестве палео-сред, разбросанных по каждому палеоконтиненту, включая прибрежные морские и пресноводные условия.[1] Большинство видов Ботриолепис характеризовались как относительно небольшие, бентосный, пресная вода детритофаги (организмы, которые получают питательные вещества, потребляя разлагающийся растительный / животный материал), в среднем около 30 сантиметров (12 дюймов) в длину.[2] Однако самый крупный вид, Б. рекс, имел примерную длину 170 сантиметров (67 дюймов). Хотя по всему миру встречается более 60 видов,[3] сравнительно Ботриолепис не является необычно более разнообразным, чем большинство современных обитающих на дне видов.[4]
Классификация
Ботриолепис это род, помещенный в отряд антиархов плакодермы. Самые ранние антиархические плакодермы впервые появились в Силурийский период палеозойской эры и встречается на каждом палеоконтинент к девонскому периоду.[5] Самые ранние представители Ботриолепис появляются к среднему девону. Антиархи, как и другие плакодермы, морфологически разнообразны и характеризуются костными пластинами, покрывающими их голову и переднюю часть туловища.[5] Ранние онтогенетические стадии плакодерм имели более тонкие костные пластинки как на голове, так и на туловище, что позволяло легко отличить ранние онтогенетические стадии плакодермы в летописи окаменелостей от таксонов, которые обладали полностью развитыми костными пластинами, но были небольшими по характеристикам.[5] Костные пластинки плакодермы обычно состояли из трех слоев, включая компактный базальный пластинчатый костный слой, средний губчатый костный слой и поверхностный слой;[5] Ботриолепис может быть классифицирован как плакодерма, поскольку она обладает этими слоями. Плакодермы вымерли к концу девона.[5] Плакодерми - это парафилетический группа клады Гнатостомы, который включает в себя всех челюстных позвоночных.[5] Неизвестно, когда именно возникли гнатостомы, но скудная летопись окаменелостей указывает на то, что это было когда-то в раннем палеозое.[6] Последний вид Ботриолепис вымер вместе с остальными Placodermi в конце девонского периода.
Общая анатомия

Голова
В головке есть два отверстия. Ботриолепис: отверстие для замочной скважины по средней линии на верхней стороне для глаз и ноздрей и отверстие для рта на нижней стороне около переднего конца головы. Открытие сохранившихся структур, которые выглядят как носовые капсулы, подтверждает веру в то, что внешние носовые отверстия располагаются на дорсальной стороне головы рядом с глазами.[7] Кроме того, положение рта на вентральной стороне черепа соответствует типичной горизонтальной ориентации покоя. Ботриолепис. У него была особенность на черепе - отдельная перегородка кости под отверстием для глаз и ноздри, окружающие носовые капсулы, называемые предглазничным углублением.
Челюсть
Новый образец от Формация Гого в бассейне Каннинг в Западной Австралии предоставили доказательства морфологических особенностей висцеральных элементов челюсти Ботриолепис. Используя образец, очевидно, что подбородочная пластинка (дермальная кость, которая формирует верхнюю часть челюсти) антиархов гомологична суборбитальной пластине, обнаруженной у других плакодерм. Кость нижней челюсти состоит из дифференцированной лопаточной и прикусной частей. Рядом с нижнечелюстным суставом располагаются прелатеральные и подлатеральные пластины, которые являются костями, несущими канал. В палатоквадрат не имеет высокого орбитального отростка и прикреплялся только к вентральной части подбородочной пластинки, что доказывает, что этмоидальная область черепной коробки (область черепа, разделяющая мозг и носовую полость) на самом деле была глубже, чем предполагалось изначально.[8] В дополнение к вышеупомянутому образцу из формации Гого было обнаружено несколько других образцов с ротовым аппаратом, удерживаемым в естественном положении мембраной, которая покрывает ротовую область и прикрепляется к боковому и переднему краям головы.[9] Ботриолепис имеет челюсть, в которой две половинки разделены, а у взрослого человека функционально независимы.[9]
Ствол

Ботриолепис У него было тонкое туловище, которое, вероятно, было покрыто мягкой кожей без чешуек или отметин. Ориентация, которая, по-видимому, была наиболее стабильной для покоя, была дорсальной поверхностью вверх, о чем свидетельствует плоская поверхность на вентральной стороне.[1] Очертание ствола предполагает, что, возможно, здесь был нотохорд присутствует в окружении перепончатой оболочки,[9] однако прямых доказательств этого нет, поскольку хорда состоит из мягких тканей, что обычно не сохраняется в летописи окаменелостей. Подобно другим антиархи, то грудной щит Ботриолепис был прикреплен к его сильно бронированной голове. Его коробчатый корпус был заключен в броневые листы, обеспечивающие защиту от хищников. К брюшной поверхности туловища прикреплена большая тонкая круглая пластинка, отмеченная глубокими линиями и поверхностными гребнями. Эта пластина находится чуть ниже отверстия в клоака.[9]
Кожный скелет
Кожный скелет состоит из трех слоев: поверхностного пластинчатого слоя, губчатого губчатого вещества и компактного базального пластинчатого слоя. Уже в раннем онтогенезе эти слои проявляются у экземпляра Ботриолепис канадский. Сначала развиваются компактные слои.[10] Предполагается, что на поверхностном слое есть зубчики, которые могли быть сделаны из ячеистой кости.[11]
Плавники и хвост

Ботриолепис имел длинную пару шиповидных грудные плавники, соединенный в основании, и снова чуть больше чем на полпути. Эти шиповидные плавники, вероятно, использовались для отрыва тела от дна; его тяжелая броня заставила бы его быстро утонуть, как только он потерял бы инерцию.[2][12] Он также мог использовать свои грудные плавники, чтобы бросать на себя осадок (грязь, песок и т. Д.). Помимо грудных, у него также было два спинных плавника: низкий удлиненный передний спинной плавник и высокий округлый задний спинной плавник. [9]- хотя предполагаемая структура спинных плавников варьируется в зависимости от конкретных видов Ботриолепис и несколько раз изменялся в реконструкциях, выпущенных исследователями по мере появления новой информации. В хвостовой хвост был удлиненным, оканчивающимся узкой полосой, но, к сожалению, редко сохраняется в окаменелостях.[9] Хотя нет согласованного объяснения их функции, Ботриолепис также имели две перепончатые брюшные оборки, расположенные на заднем конце туловищного панциря по обе стороны от хвоста, каждая из которых имеет две отдельные области.[7] Нет никаких доказательств того, что оборки были задействованы в поддержке скелета, но возможно, что они либо функционировали как плавники, либо участвовали в воспроизводстве, и, возможно, даже присутствовали у одного пола, но не у другого.[7]
Мягкая анатомия

Структуры, состоящие из мягких тканей, обычно не сохраняются в окаменелостях, потому что они легко разрушаются и разлагаются намного быстрее, чем твердые ткани, а это означает, что в летописи окаменелостей часто не хватает информации о внутренней анатомии ископаемых видов. Однако сохранение структур мягких тканей может иногда происходить, если отложения заполняют внутренние структуры организма после или после его смерти. В статье Роберта Денисона «Мягкая анатомия Ботриолепис"исследует формы и органы Ботриолепис.[7] Эти внутренние структуры были сохранены, когда различные типы отложений, окружающие внешность животного, заполнили внутренние панцири (таким образом могли быть сохранены только органы, которые сообщаются с внешним миром). На разных участках месторождения были выявлены три различных типа отложений. Ботриолепис: первый - бледный зеленовато-серый песчаник средней текстуры, состоящий в основном из кальцита; второй - подобный, но более мелкий осадок, сохраняющий многие формы органов; и третий - отчетливый мелкозернистый алевролит, состоящий из кварца, слюды и других минералов, но без кальцита.[7] Эти отложения помогли сохранить следующие внутренние элементы:
Пищеварительная система
В целом пищеварительная система из Ботриолепис - который включает органы, участвующие в приеме пищи, пищеварении и удалении отходов - можно охарактеризовать как простое и прямое, в отличие от человеческого. Он начинается на переднем конце организма небольшой ротовой полостью, расположенной над задней частью пластин верхней челюсти. Кзади от рта пищеварительная система простирается в более широкую и сплющенную в дорсовентральном направлении область, называемую глотка, из которых возникают и жабры, и легкие. В пищевод, который также характеризуется как сплющенная в дорсо-вентральном направлении трубка, проходит ото рта в желудок и приводит к уплощенной эллипсоидальной структуре. Эта структура может быть гомологичной переднему концу кишечника других рыб.[7] Плоскостность этих структур могла быть преувеличена, когда ископаемые образцы испытали тектоническая деформация через геологическое время. Кишечник начинается узко на переднем конце, расширяется в поперечном направлении и затем снова сужается кзади к цилиндрической прямой кишке, которая заканчивается как раз в пределах заднего конца панциря туловища. Хотя пищеварительная система примитивна по своей природе и не имеет расширенной области желудка, она специализируется на независимо приобретенном сложном спиральном клапане, сравнимом с таковым у эластожаберные ветви и многие костлявые рыбы, похожие на те, что встречаются у некоторых акул. Единственная складка ткани, свернутая вокруг своей оси, образует этот специализированный спиральный клапан.[7]
Жабры
Предполагается, что жабры Ботриолепис относятся к примитивному типу, хотя их структура до сих пор не изучена. С боков они окружены оперкулярной складкой и находятся в пространстве под боковой частью головного щита, проходя медиально под ним. нейрокраниум. По сравнению с жабрами рыб нормальной формы, жаберная область Ботриолепис считается расположенным дорсальнее, спереди более густо, и в целом относительно короткий и широкий.[7]
Парные вентральные мешки


Кзади от туловищного панциря отходят парные вентральные мешочки, доходящие до переднего конца спиральной кишки. Кажется, что мешочки берут начало в глотке и представляют собой единую срединную трубку, которая затем расширяется кзади и в конечном итоге разделяется на два мешочка, которые могут быть гомологичными. легкие определенных дипноанцы и четвероногие.[7] Была выдвинута гипотеза, что эти легкие в сочетании с сочлененными руками и жестким поддерживающим скелетом позволили бы Ботриолепис путешествовать по суше. Кроме того, как Роберт Денисон[7] заявляет, потому что нет никаких доказательств связи между внешними Нарис и рот, Ботриолепис вероятно, дышал так же, как современные двоякодышащие, то есть помещая рот над поверхностью воды и глотая воздух.
Несмотря на оригинальную интерпретацию, представленную Денисоном в 1941 году, не все палеонтологи согласны с тем, что плакодермы любят Ботриолепис на самом деле обладал легкими. Например, в его статье «Легкие» у плакодерм, стойкий палеобиологический миф, связанный с предвзятыми интерпретациями окружающей среды, D. Goujet предполагает, что, хотя следы некоторых органов пищеварения могут быть очевидны из осадочных структур, нет никаких доказательств, подтверждающих присутствие легких в образцах из формации Эскуминак в Канаде, на которых основывалось первоначальное утверждение. Он отмечает, что мировое распространение Ботриолепис ограничивается строго морской средой, и поэтому считает, что наличие легких в Ботриолепис неопределенно. Вероятно, необходимо дальнейшее исследование окаменелостей, чтобы сделать вывод о наличии легких в Ботриолепис.[13]
Кормление
Ботриолепис, как и все другие антиархы, считается, что они питались, проглатывая глотки грязи и других мягких отложений, чтобы переварить детрит, мелкие или микроорганизмы, водоросли и другие формы органических веществ в проглоченных отложениях. Кроме того, расположение рта на вентральной стороне головы предполагает, что Ботриолепис вероятно, питался снизу. Считается, что регулярное присутствие «углеродистого материала в пищеварительном тракте» указывает на то, что большая часть его рациона состояла из растительного материала.[7]
Распределение
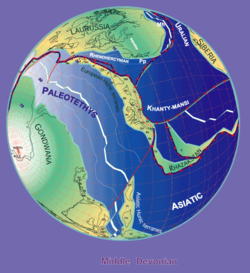
Авторы Stampfli & Borel, 2000 г.
Ботриолепис ископаемые найдены в Среднем и Позднем Девонский страты (от 387 до 360 миллионов лет назад).[12] Поскольку окаменелости находятся в пресноводных отложениях, Ботриолепис предполагается, что провел большую часть своей жизни в пресная вода реки и озера, но, вероятно, смог войти соленая вода а также потому, что его ареал соответствовал континентальным берегам Девона. Большие группы Ботриолепис экземпляры найдены в Азии, Европе, Австралии (Формация Гого ),[9] Пенсильвания (формация Кэтскилл),[1] Квебек (Формация Эскуминака ),[1] Вирджиния (Чемунг),[14] Колорадо,[14] Формация Куч (Boyacá, Колумбия),[15] и по всему миру.
Сайт Catskill Formation
В Формация Кэтскилл (Верхний девон, фаменский ярус), расположенный в Округ Тиога, Пенсильвания, это место обитания большой выборки мелких особей Ботриолепис. Образец был взят из серии каменных плит, которые состояли из частичных или полных сочлененных внешних каркасов. Было обнаружено, что более двухсот особей плотно сбиты вместе, практически не перекрываясь. Из этого образца много информации о характеристиках молоди Ботриолепис можно определить. Морфометрическое исследование, проведенное Джейсоном Даунсом и соавторами, выделяет определенные характеристики, которые указывают на молодость у Ботриолепис, в том числе умеренно крупная голова и умеренно крупная орбитальное отверстие - обе эти характеристики также были признаны Эриком Стенсио в 1948 году в самых маленьких B. canadensis лиц.[1] Несколько других черт, которые Стенсио отметил для молодых особей, также можно увидеть в образце Катскилл. Эти особенности включают «тонкие дермальные кости с орнаментом, состоящим из непрерывных анастомозирующих гребней, а не бугорков, дорсальный щиток туловища уже, чем длинный, и с непрерывным и ярко выраженным дорсальным срединным гребнем, и предсрединную пластину, которая шире, чем длинна».[1]
Виды
Палеонтология позвоночных в значительной степени зависит от способности различать разные виды таким образом, чтобы это было согласовано как в пределах определенного рода, так и среди всех организмов. Род Ботриолепис не является исключением из этого принципа. Ниже перечислены некоторые известные виды в пределах Ботриолепис; всего было названо более шестидесяти видов, и вполне вероятно, что значительная часть из них является действительной из-за космополитической природы Ботриолепис.[3]
Ботриолепис канадский

Ботриолепис канадский - таксон, который часто служит модельным организмом для отряда Antiarchi из-за его огромной выборки полных, неповрежденных экземпляров, найденных в Формация Эскуминака в Квебек, Канада.[1] Из-за огромного размера выборки этот вид часто используется для сравнения данных о росте недавно приобретенных образцов Ботриолепис, в том числе найденные в формации Катскилл, упомянутой выше. Это сравнение позволяет исследователям определить, относятся ли вновь обнаруженные образцы к молодым особям или к новым видам "Bothriolepis".
B. canadensis был впервые описан в 1880 году Дж. Ф. Уайтавсом с использованием ограниченного числа обезображенных образцов. Следующим, кто предложил реконструкцию вида, был В. Паттен, опубликовавший свои открытия в 1904 году после открытия нескольких образцов, хорошо сохранившихся в трехмерном пространстве. В 1948 г. Э. Стенсио опубликовал подробное описание B. canadensis анатомия с использованием большого количества материала, которая в конечном итоге стала наиболее общепринятым описанием этого вида. После публикации Стенсио многие другие представили реконструированные модели B. canadensis с измененными аспектами анатомии, включая модифицированный единственный спинной плавник Везины и недавние реконструкции Арсено и другие из экземпляров с небольшим тафономическим искажением. В настоящее время модель Арсено и другие. считается наиболее точным, хотя до сих пор ведется много споров о различных аспектах внешней анатомии этого вида. Несмотря на неопределенность, B. canadensis до сих пор считается одним из самых известных видов.[16]
Внешний скелет из Ботриолепис канадский состоит из клеточной костной ткани дермы и характеризуется четкой горизонтальной зональностью или расслоением.[10] Модель рыбы имеет среднюю общую длину 43,67 см (17,19 дюйма) и среднюю длину кожной брони 15,53 см (6,11 дюйма), что составляет 35,6% от расчетной общей длины.[16] Как и многие антиархи, B. canadensis также имел узкие грудные плавники, гетероцеркальный хвостовой плавник (то есть хорда простирается в верхнюю долю хвостового хвоста) и большой спинной плавник, который, вероятно, не играет важной роли в движении, а вместо этого действует скорее как стабилизатор [16]
Ботриолепис рекс
Первоначально описано Даунсом и соавт. (2016), Ботриолепис рекс взяты из формации Нордстранд-Пойнт острова Элсмир, Канада. Длина тела B. rex оценивается в 1,7 метра и поэтому является самым крупным из известных видов Ботриолепис. Его броня особенно толстая и плотная даже с учетом его размера. Даунс и др. (2016) предполагают, что это могло как защитить животное от крупных хищников, так и служить балластом для предотвращения всплытия этого крупного донного обитателя на поверхность.
Ботриолепис вирджинский


Первоначально описано Weems et al. в 1981 г. этот вид, Ботриолепис вирджинский, происходит от "Chemung", недалеко от Винчестер, Вирджиния. Несколько черт, обнаруженных в B. virginiensis также можно найти у других видов Ботриолепис, (особенно Б. нитида), включая задние косые борозды сенсорной линии головы, которые встречаются относительно далеко вперед на затылочной пластине, относительно удлиненное глазничное отверстие и низкий передне-срединно-дорсальный гребень.[14] Характеристики, отличающие B. virginiensis от других видов включают, но не ограничиваются ими, сросшиеся головные швы, слитые элементы в дистальных сегментах грудных плавников взрослых особей и длинную премедианную пластину относительно длины головного щита.[14]
В настоящее время ведется много споров о том, B. virginiensis и Б. нитида действительно можно отличить друг от друга. Томсон и Томас утверждают, что пять видов Ботриолепис из Соединенных Штатов (B. nitida, B. minor, B. virginiensis, B. darbiensis и B. coloradensis) невозможно последовательно отличить друг от друга.[4] Наоборот, Вимс утверждает, что существует несколько признаков, которые отличают виды друг от друга, включая несколько перечисленных выше.[14]
Ботриолепис нитида
Этот вид, встречающийся в современных Пенсильвания, был первоначально описан Дж. Лейди в 1856 году. Как упоминалось выше, существует много споров относительно различимости между Б. нитида и B. virginiensis, однако на основании доказательств, представленных Weems (2004),[14] есть несколько отличительных черт, характерных для каждого вида. Б. нитида имеет максимальную длину лобового щита 65 миллиметров (2,6 дюйма), узкую и неглубокую трехраздельную предглазничную выемку, имеет передне-срединно-дорсальную (ВМД) пластину, которая шире, чем ее длина, и вентральный грудной щит с выпуклыми боковыми краями.[14]
Ботриолепис колорадский
Впервые описанный Истманом в 1904 г., этот вид был обнаружен локализованным в современных Колорадо. Есть вероятность, что этот вид похож, если не идентичен, Б. нитида, однако, поскольку имеющиеся материалы относительно B. coloradensis фрагментирован, сравнить эти два вида с какой-либо степенью уверенности невозможно.[14]
Ботриолепис африканский
Ботриолепис африканский [17] это Ботриолепис виды, известные с высших палеоширота, описанные из месторождений, первоначально заложенных в Поздний девон Антарктический круг. Остатки были извлечены исключительно из одного углеродистого сланца вблизи кровли позднего девона. Фаменский, Формация Витпорт (группа Виттеберга), обнаженная в выемке дороги к югу от Маханды /Grahamstown в Южной Африке. Этот сайт, Waterloo Farm lagerstätte интерпретируется как представление прибрежной лагуны с обратным барьером, с морскими и речными влияниями.[18] Гесс заметил, что Ботриолепис на ферме Ватерлоо было меньше, чем в большинстве Ботриолепис-носящие местонахождения, хотя представлен полный онтогенетический ряд. Длина брони головы и туловища варьировалась от 20 до 300 мм, что на основе пропорций двух самых маленьких особей (у которых сохранены отпечатки хвоста) переводится в полную длину тела, варьирующуюся от 52 до 780 мм.[19]Согласно с [20] Ботриолепис африканский считался наиболее похожим на Ботриолепис барретти [21] с конца Живетян из Антарктида. Сходство между ними было использовано, чтобы предположить вывод Ботриолепис африканский с востока Гондванец Окружающая среда. [22]


использованная литература
- ^ а б c d е ж г Даунс, J.P .; Criswell, K.E .; Даешлер, Э. (Октябрь 2011 г.). «Массовая смертность молодых антиархов (Ботриолепис sp.) из формации Кэтскилл (верхний девон, фаменский ярус), графство Тиога, Пенсильвания ". Труды Академии естественных наук Филадельфии (161): 191–203.
- ^ а б Палмер, Д., изд. (1999). Иллюстрированная энциклопедия динозавров и доисторических животных Маршалла. Лондон: Marshall Editions. п. 33. ISBN 978-1-84028-152-1.
- ^ а б Янг, Г. (2010). «Плакодермы (бронированная рыба): доминирующие позвоночные девонского периода». Ежегодный обзор наук о Земле и планетах. 38: 523–550. Дои:10.1146 / аннурьев-земля-040809-152507.
- ^ а б Thomson, K.S .; Томас, Б. (август 2001 г.). «О статусе вида Ботриолепис (Placodermi, Antiarchi) в Северной Америке ». Журнал палеонтологии позвоночных. 21 (4): 679–686. Дои:10.1671 / 0272-4634 (2001) 021 [0679: otsoso] 2.0.co; 2.
- ^ а б c d е ж Йохансон, Зерина; Тринайстич, Кейт (2014). «Ископаемые онтогенезы: вклад онтогенеза плакодермы в наше понимание эволюции ранних гнатостомов». Палеонтология. 57 (3): 505–516. Дои:10.1111 / pala.12093.
- ^ Бразо, М. (2009). «Мозг и челюсти девонского акантода и современные гнатостомы» (PDF). Природа. 457 (7227): 305–308. Дои:10.1038 / природа07436. HDL:10044/1/17971. PMID 19148098.
- ^ а б c d е ж г час я j k Денисон, Р.Х. (сентябрь 1941 г.). "Мягкая анатомия Ботриолепис". Журнал палеонтологии. 15 (5): 553–561.
- ^ Янг, Г. (1984). "Реконструкция челюстей и мозга у девонской плакодермы. Ботриолепис". Палеонтология. 27 (3): 635–661.
- ^ а б c d е ж г Паттен, В. (июль 1904 г.). "Новые факты, касающиеся Ботриолепис". Биологический бюллетень. 7 (2): 113–124. Дои:10.2307/1535537. JSTOR 1535537.
- ^ а б Даунс, J.P .; Донохью, П.С.Дж. (2009). "Гистология скелета Ботриолепис канадский (Placodermi, Antiarchi) и эволюция скелета у происхождения челюстных позвоночных ". Журнал морфологии. 270 (11): 1364–1380. Дои:10.1002 / jmor.10765. PMID 19533688.
- ^ Джайлз, С. (2013). «Гистология дермальных скелетов плакодермы: значение для природы предкового гнатостома». Журнал морфологии. 274 (6): 627–644. Дои:10.1002 / jmor.20119. ЧВК 5176033. PMID 23378262.
- ^ а б «Музей эпохи рыб - окаменелости». Музей эпохи рыб, Новый Южный Уэльс, Австралия.
- ^ Гуже, Д. (2011). ""Легкие «в плакодермах, стойком палеобиологическом мифе, связанном с предвзятыми интерпретациями окружающей среды». Comptes Rendus Palevol. 10 (5–6): 323–329. Дои:10.1016 / j.crpv.2011.03.008.
- ^ а б c d е ж г час Weems, R.E. (Март 2004 г.). "Ботриолепис viginiensis, действительный вид рыб плакодермы, отделяемый от Ботриолепис нитида". Журнал палеонтологии позвоночных. 24 (1): 245–250. Дои:10.1671/20.
- ^ Жанвье, Филипп; Вильярроэль А., Карлос (1998). "Los Peces Devónicos del Macizo de Floresta (Бояка, Колумбия). Учитывает taxonómicas, bioestratigráficas, biogeográficas y ambientales". Geología Colombiana. 23: 3–18. Получено 2017-03-31.
- ^ а б c Bechard, I .; Arsenault, F .; Cloutier, R .; Керр, Дж. (2014). "Девонская рыба Ботриолепис канадский пересмотрены с трехмерными цифровыми изображениями ". Palaeontologia Electronica. 17 (1).
- ^ ЛОНГ, Дж. А., АНДЕРСОН, М. Э., ГЕСС, Р. В. и ХИЛЛЕР, Н. (1997). Новые рыбы-плакодермы из позднего девона в Южной Африке. Журнал палеонтологии позвоночных 17,253–268.
- ^ Гесс, Роберт В .; Уитфилд, Алан К. (14 февраля 2020 г.). «Эволюция эстуарных рыб и четвероногих: выводы из Гондванского устьевого озера позднего девона (фамена) и эквивалента южноафриканского голоцена». Биологические обзоры. DOI: 10.1111 / brv.12590. PMID 32059074
- ^ GESS, R.W. (2011). Образцы биоразнообразия фаменского периода в Гондване на высоких широтах - свидетельства южноафриканской формации Витпорт (Cape Supergroup, WittebergGroup). Диссертация на соискание степени доктора наук: Университет Витватерсранда, Йоханнебург
- ^ ЛОНГ, Дж. А., АНДЕРСОН, М. Э., ГЕСС, Р. В. и ХИЛЛЕР, Н. (1997). Новые рыбы-плакодермы из позднего девона в Южной Африке. Журнал палеонтологии позвоночных 17,253–268.
- ^ ЯНГ, Г. (1984). Реконструкция челюстей и черепа у девонской плакодермы Bothriolepis. Палеонтология 27, 635–661.
- ^ ЛОНГ, Дж. А., АНДЕРСОН, М. Э., ГЕСС, Р. В. и ХИЛЛЕР, Н. (1997). Новые рыбы-плакодермы из позднего девона в Южной Африке. Журнал палеонтологии позвоночных 17,253–268.
внешние ссылки
| |||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||

