Cartorhynchus - Cartorhynchus
| Cartorhynchus | |
|---|---|
 | |
| Трехмерная реконструкция черепа, вид снизу слева, обнажающие зубы и внутреннюю крышу черепа. | |
| Научная классификация | |
| Королевство: | Animalia |
| Тип: | Хордовые |
| Учебный класс: | Рептилии |
| Clade: | †Ихтиозавроморфа |
| Clade: | †Ихтиозаврообразные |
| Clade: | †Насоростра |
| Род: | †Cartorhynchus Мотани и другие., 2014 |
| Разновидность: | †С. lenticarpus |
| Биномиальное имя | |
| †Cartorhynchus lenticarpus Мотани и другие., 2014 | |
Cartorhynchus (что означает «укороченная морда») - вымерший род из рано ихтиозавриформ морская рептилия которые жили во время Ранний триас эпоха около 248 миллионов лет назад. Род состоит из одного вида, Cartorhynchus lenticarpus, названный в 2014 году Риосуке Мотани и его коллегами из единственного почти полного скелета, найденного недалеко от Чаоху, Провинция Аньхой, Китай. Вместе со своим близким родственником Склерокормус, Cartorhynchus была частью диверсификации морских рептилий, которая произошла внезапно (примерно за миллион лет) во время Спатианский подэтап, вскоре после разрушительного Пермско-триасовое вымирание, но впоследствии они были истреблены из-за вулканизма и изменения уровня моря из-за Средний триас.
Длиной около 40 сантиметров (16 дюймов), Cartorhynchus было маленьким животным с телом, похожим на ящерицу, и коротким туловищем; вероятно, он плавал, как угорь, на медленных скоростях. Его конечности имели обширный хрящ и могли сгибаться, как ласты, что, возможно, позволяло ему ходить по суше. Наиболее отличительные особенности Cartorhynchus были его короткая суженная морда и несколько рядов коренной зуб -подобные зубы, выросшие на внутренней поверхности его челюстных костей. Эти зубы не были обнаружены до тех пор, пока образец не был подвергнут КТ сканирование. Cartorhynchus вероятно, охотятся на беспозвоночных с твердым панцирем, используя всасывающее питание, хотя как именно он использовал обращенные внутрь зубы, пока не известно. Это было одно из пяти независимых приобретений моляровидных зубов среди ихтиозавров.
Открытие и наименование

В 2011 г. единственный известный экземпляр Cartorhynchus был обнаружен в пласте 633 на втором уровне карьера Мацзяшан недалеко от центра города. Чаоху, Провинция Аньхой, Китай; пласты горных пород в этом карьере относятся к верхней пачке Формация Нанлингху.[1] Экземпляр состоит из почти полного скелета, в котором отсутствует только часть хвоста.[2] и некоторые кости левой части задней части черепа. Сохранность образца, вероятно, была результатом того, что он был отложен в осадке правой стороной вниз, таким образом оставив левую сторону открытой для элементов. Он получил полевой номер МТ-II, а позже номер экземпляра AGB 6257 на Геологический музей Аньхой.[3]
В 2014 году экземпляр был описан Риосуке Мотани и его коллегами в Природа как представитель нового рода и вида, Cartorhynchus lenticarpus. Они получили родовое имя Cartorhynchus от греческих слов Картос (καρτοσ, «укороченный») и ринчиос (ρηψνχηοσ, «морда»), а конкретное имя чечевица от латинских слов лентус («гибкий») и запястье ("запястье"). Оба названия относятся к анатомическим характеристикам, которые он имел бы при жизни.[1] Образец считался беззубым, пока не был обнаружен изолированный зуб во время дальнейших попыток удалить камень между закрытыми челюстями. Поскольку образец был слишком хрупким, чтобы обнажить внутреннюю часть челюстей, Цзянь-Донг Хуанг, Мотани и другие коллеги впоследствии отсканировали и визуализировали образец в 3D, используя микрокомпьютерная томография (микро-КТ), проведенная в Yinghua Testing Company в г. Шанхай, Китай. В 2020 году результаты их дальнейшей работы были опубликованы в Научные отчеты.[3]
Описание

На момент открытия Cartorhynchus был самым маленьким известным членом Ихтиозаврообразные. Сохранившийся образец имел длину 21,4 см (8 дюймов); Предполагая, что у него были пропорции хвоста, сопоставимые с пропорциями хвоста близких родственников, Мотани и его коллеги оценили полную длину тела в 40 сантиметров (1 фут 4 дюйма) и вес в 2 килограмма (4,4 фунта).[1][2]
Череп
Cartorhynchus имел необычно короткую и стянутую морду, занимавшую лишь половину длины черепа, и глубокую челюсть. Кончик морды был всего 6 миллиметров (0,2 дюйма) в ширину.[3] В отличие от большинства рептилий, его носовая кость достиг передней части морды. Благодаря так же удлиненной предчелюстная кость, его костные ноздри были расположены относительно далеко позади черепа, а его лобная кость также не хватало расширения в заднем внешнем углу. Все эти характеристики были общими с его близким родственником. Склерокормус. Однако, в отличие от последнего, лобная кость не влияла на глазницу в Cartorhynchus; то префронтальный и постфронтальные кости не встречались над глазницей;[4] и расположение большого отверстия для шишковидная железа на крыше черепа отличался: он находился на стыке лобной и теменные кости в Склерокормус, но исключительно на теменных костях в Cartorhynchus. Cartorhynchus также имел характерно большой подъязычная кость.[1][5]

Первоначально Мотани и его коллеги пришли к выводу, что Cartorhynchus был беззубым; тем не менее, сканирование с помощью микро-КТ впоследствии показало наличие округлых, коренной зуб -подобные зубы, которые выступают внутрь почти перпендикулярно длинной оси челюсти, что делает их невидимыми снаружи. Все зубы были либо полностью сплюснутыми, либо слабо заостренными, и многие зубы имели сужение между корнем и коронкой. В отличие от других ихтиозавров с коренными зубами, все коронки зубов были «раздутыми» в одинаковой степени. На верхняя челюсть (верхняя челюсть) и зубной (нижняя челюсть) зубы располагались в три ряда, причем крайний ряд имел самые большие и большие зубы; у верхнечелюстных костей по семь, пять и, вероятно, по одному зубу, а у зубных костей по десять, семь и четыре зуба. Среди ихтиозавриформ только Cartorhynchus и Ксинминозавр иметь несколько рядов зубов.[6] Расположение зубов означало, что самые передние нижние зубы не имели соответствующего верхнего зуба, а также что два зубных протеза, образующие нижнюю челюсть, не могли быть плотно слиты. Эту характеристику можно было бы разделить со своими близкими родственниками, беззубыми hupehsuchians.[3][7]
Посткраниальный скелет

Cartorhynchus кажется, было 5 шейные позвонки и 26 задние позвонки, всего 31 прекрестцовый позвонок (позвонки перед своим крестец, или бедро). Вместе с Склерокормус (с 34 прекакральными) и Chaohusaurus (с 36 пре-крестцовыми), Cartorhynchus попадает в типичный диапазон для наземных животных, в отличие от 40-80 пре-крестцовых, распространенных среди более полученный (специализированный) ихтиоптериги.[1] В отличие от Склерокормус, то нервные отростки выступающий из верхушки позвонков в Cartorhynchus были относительно узкими и наклонными вместо широких и фланцевых. Cartorhynchus также можно отличить по парапофизам, позвоночным отросткам, которые соединяются с ребрами; их передние края сливались с краями позвонков.[5]
Обе Cartorhynchus и Склерокормус имел плотно сложенные грудные клетки, которые были наиболее глубокими около плеча, с широкими, уплощенными и толстостенный ребра, что обычно наблюдается у ранних представителей вторично-водных рептилий.[1][8] При ихтиоптеригии эти уплощенные ребра исчезли, за исключением Моллезавр.[9] На нижней стороне груди гастралия из Cartorhynchus были тонкими и стержневидными, в отличие от приплюснутой «корзины» Склерокормус, но у обоих отсутствовала еще одна пара симметричных элементов по средней линии тела.[5]
Конечности Cartorhynchus были плохо окостеневший (окостенели только три пальца руки) с широко расставленными костями, особенно между кости запястья (запястья) и пальцев, что указывает на наличие обширного хряща в конечностях. Это сделало бы конечности подобными ластам. Ласты передних конечностей Cartorhynchus были изогнуты назад, при этом пальцы были наклонены на 50 ° относительно оси длинных костей (zeugopodium), в то время как задние конечности были изогнуты вперед. В бедренная кость из Cartorhynchus был прямым и не расширялся на нижнем конце. Склерокормус у них были похожие конечности, за исключением того, что они были лучше окостеневшими и их сохранившаяся кривизна могла быть не естественной.[1][5]
Классификация

Отсутствие полных ископаемых останков привело к отсутствию ясности в отношении происхождения ихтиоптеригии, включая ихтиозавры. Многие годы считалось, что их окаменелости внезапно появились в Средний триас с сильными водными приспособлениями. Открытие Cartorhynchus и Склерокормус частично восполнили этот пробел. Филогенетические анализы Мотани и его коллеги обнаружили, что эти двое были тесно связаны друг с другом, образуя клады (группа) назвала Насоростра - и к ихтиоптеригии, к которой назоространс сформировал сестринская группа. Cartorhynchus и Склерокормус были объединены их короткой мордой, удлиненными носами, глубокими челюстями, недоразвитыми лобными частями, наиболее глубокими грудными клетками около плеча и лопатки (лопатки) на нижнем конце шире, чем на верхнем.[1][5]
Включение nasorostrans в филогенетический анализ также предоставило доказательства в поддержку hupehsuchians как близких родственников ихтиоптеригов. В 2014 году Мотани и его коллеги назвали кладу, образованную Nasorostra и Ichthyopterygia, Ichthyosauriformes, а кладу, образованную Ichthyosauriformes и Hupehsuchia, - Ихтиозавроморфа. Примечательно, что тесная связь между этими различными группами была восстановлена их анализами независимо от того, были ли из анализа удалены характеристики, связанные с водной адаптацией.[1][5] Такие характеристики могли развиться гомоплазно (из конвергентная эволюция ) среди нескольких линий из-за схожего образа жизни, что может смещать филогенетический анализ, чтобы реконструировать их как гомологии (происходит от общего происхождения). Устойчивость этих реконструированных отношений даже после удаления водных характеристик указывает на их надежность.[10][11]
Ниже филогенетическое дерево из филогенетического анализа, опубликованного Хуангом, Мотани и его коллегами в 2019 г., в описании Chaohusaurus brevifemoralis, воспроизводится частично.
| Ихтиозавроморфа |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Эволюционная история
Появление ихтиозавроморфов было частью восстановления морских экосистем после разрушительного Пермско-триасовое вымирание. Обычно считалось, что морские экосистемы не восстановили свое полное разнообразие до 5-10 миллионов лет после массового вымирания, и что морские рептилии восстанавливались после исчезновения медленнее, чем другие линии.[12][13] Однако открытие множества разнообразных фаун морских рептилий, обитающих в раннем триасе, включая Cartorhynchus - впоследствии показал, что это не так.[7][14] В частности, похоже, что ихтиозавриформы впервые появились во время спатийского подэтапа Оленекян сцена и быстро достигла высоких функциональное разнообразие в первый миллион лет их эволюции.[5] Они занимали множество ниши несмотря на относительно низкое видовое разнообразие,[15] включая оба демерсальный (обитающие на дне) виды, такие как hupehsuchians и nasorostrans, и пелагический (открытая вода) виды, такие как ихтиоптериги.[5]

Многие ихтиозавриды раннего и Средний триас имели молярные зубы (показаны на филогенетическом дереве выше), в том числе Cartorhynchus; такие зубы указывают на диету, хотя бы частично основанную на животных с твердым панцирем.[16] В 2020 году Хуанг и его коллеги выполнили реконструкция родового государства зубов среди ихтиозавроморфов. Вероятностные методы предполагают, что округлые или плоские зубы, скорее всего, развивались независимо пять раз, тогда как методы, основанные на скупость предположил, что они развивались независимо от трех до пяти раз. Хуанг и его коллеги заметили, что развитие моляризованных зубов происходило независимо много раз у водных животных (включая несколько клонов варан, мурена, и спарид и цихлида рыба), и, следовательно, частота встречаемости среди ихтиозавриформ не является необычно высокой. Они также заметили, что у ихтиозавриформ раннего триаса обычно были маленькие округлые зубы; зубы ихтиозавров среднего триаса были более разнообразными по размеру и форме, что коррелирует с увеличением разнообразия беспозвоночных.[17] Таким образом, они предположили, что диверсификация ихтиозавриформ частично была вызвана эволюцией добычи с твердым панцирем.[3]
Однако hupehsuchians и nasorostrans в конечном итоге вымерли на границе между ранним и средним триасом, создав «таксономическое узкое место». На границе меняется уровень моря[18] и вулканизм[19] привели к плохо насыщенным кислородом океанам, создавая характерные сигнатура изотопа углерода от разлагающегося органического материала в пластах горных пород на границе.[20] Ихтиозавриформы не восстановили свое разнообразие после этого оборота,[21] с Зауроптеригия и Saurosphargidae запуск второй волны диверсификации, продолжающейся от трех до пяти миллионов лет.[5][15]
Палеобиология
Рацион питания
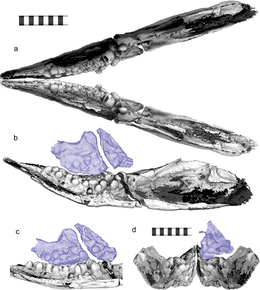
Мотани и его коллеги предположили в 2014 году, что Cartorhynchus был всасывающий питатель который питался, концентрируя давление в своей узкой морде. Это также было подтверждено прочностью его подъязычной и поджаберной части (которые должны были закрепить язык), а также неправильным наблюдением беззубости.[1] Впоследствии аналогичные выводы были сделаны для Склерокормус.[5] Шастазавр и Шонизавр ранее интерпретировались как ихтиозавры, питающиеся всасыванием,[22] но количественный анализ ихтиозавров триасового и юрского периодов, проведенный Мотани и его коллегами в 2013 году, показал, что ни у одного из них не было достаточно крепких поджаберных костей и достаточно узких морд, чтобы можно было кормить их всасыванием.[23]
Открытие коренных зубов в Cartorhynchus привели Хуанга и его коллег к выводу в 2020 году, что Cartorhynchus был дурофаг, питаясь добычей с твердым панцирем. Они отметили, что это не противоречит образу жизни с отсасыванием; некоторые спаридные рыбы являются одновременно дурофагами и питаются всасыванием.[24] Однако они предположили, что Cartorhynchus были ограничены питанием мелкой добычей. Что касается горизонтальной ориентации зубьев, они наблюдали поверхности износа, которые указывали на то, что стороны зубьев окклюзия друг с другом, а не коронами. Тем не менее, они отметили, что зубы структурно сильнее всего на концах, а не на боковых сторонах, под высоким подчеркивает раздавливающих укусов.[25] Они предположили, что челюсть могла быть скручена во время консервации, или мягкие ткани, такие как коллаген которые удерживали зубы при жизни, возможно, были утеряны, хотя они признали, что ни одна из гипотез не может объяснить характер износа. Наконец, они отметили, что нижние зубы без соответствующих верхних зубов были необычными; на них нет следов износа, и нет никаких доказательств наличия мышечных механизмов, которые позволили бы использовать две челюсти друг против друга. Поэтому они сделали вывод, что эти зубы, вероятно, не использовались против других зубов.[3]
Конечности и движения

Cartorhynchus у него были плохо окостеневшие конечности, несмотря на хорошо окостеневшие череп и позвонки. Однако в 2014 году Мотани и его коллеги предположили, что это все еще взрослый человек, потому что многие ранне расходящиеся представители морских рептилий имеют плохо окостеневшие конечности. педоморфоз (сохранение незрелых черт во взрослом возрасте), хотя они не полностью отвергли возможность того, что это был молодой особь из-за существования только одного экземпляра.[1]
В случае Cartorhynchus, Мотани и его коллеги предположили, что большие, похожие на ласты передние конечности позволили ему передвигаться по суше, что сделало его амфибией. Обширный хрящ в лучезапястном суставе позволил бы плавнику сгибаться без локтя; У молодых морских черепах есть такие же хрящевые ласты, которые они используют для передвижения по суше.[26] Хотя его плавник не был бы особенно сильным, Cartorhynchus был относительно легким, с соотношением массы тела к площади поверхности ласт меньше, чем у Chaohusaurus. Изогнутые ласты позволили бы держать их близко к телу, тем самым увеличивая их механическое преимущество. Мотани и его коллеги предположили, что другие черты Cartorhynchus также способствовали бы образу жизни амфибий, включая короткое туловище и морду, а также утолщенные ребра (которые служили бы балласт, стабилизирующий животное в прибрежных водах).[1]

В исследовании 2019 года, проведенном Сусаной Гутарра и его коллегами, использовалось компьютерное моделирование для оценки затрат энергии на плавание у ихтиозавриформ. Раннерасходящиеся ихтиозавры с телами в форме ящериц и удлиненными, без лавинообразными хвостами, как Cartorhynchus, нанял бы угловатый (похожее на угря) плавание, в то время как более поздние ихтиозавры с более глубокими, похожими на рыбу телами и хорошо выраженными хвостовыми сосальщиками использовали панцирная форма (похожее на макарель) плавание. Принято считать, что угловатое плавание менее эффективно, чем тигровое плавание.[27][28][29] Действительно, Гутарра и его коллеги обнаружили, что затраты энергии на плавание со скоростью 1 метр в секунду (3,3 фута / с) были в 24-42 раза выше в Cartorhynchus чем ихтиозавр Офтальмозавр (в зависимости от того, учитывается ли режим плавания), и его коэффициент трения был на 15% выше, чем у бутылконосый Дельфин. Тем не мение, Cartorhynchus вероятно, плавали бы на более медленных скоростях, требующих меньшей эффективности, и преимущества плавания в форме панциря у более поздних, более крупных ихтиозавриформ также были компенсированы увеличением размера тела.[30]
Палеоэкология

Кровать 633 карьера Мацзяшан, местность, где Cartorhynchus был найден, это слой серого глинистый (глинистый) известняк расположен на высоте 13 метров (43 фута) над основанием верхней части формации Нанлингху. Он определяется сверху и снизу слоями желтоватого цвета. мергель. С точки зрения аммонит биостратиграфия, эта кровать принадлежит Субколумбиты зона.[1][31] Оценки дат с высоким разрешением были произведены для отложений оленекского возраста, обнаженных в карьере Маджашан, на основе изотопных записей углерод-13 кататься на велосипеде и спектральные гамма-луч бревна (которые измеряют количество радиации в горных породах астрономического происхождения); Слой 633, в частности, был оценен в 248,41 Ма в возрасте.[32][33][34] В среднем триасе пласты Чаоху откладывались в океанический бассейн относительно далеко от побережья, которое с юга ограничивалось более мелководьем и карбонатные платформы, а на севере по континентальный склон и более глубокие бассейны.[35][36]
Ихтиозавриформы Склерокормус и Chaohusaurus оба найдены в карьере Мацзяшан вместе с Cartorhynchus; Склерокормус известен из более молодого слоя 719 (248,16 млн лет), а Chaohusaurus находится в обеих кроватях.[32] Зауроптеригий Majiashanosaurus Известен из слоя 643.[37] Разнообразие рыб в карьере Мацзяшан ниже, чем в других местах; самая распространенная рыба - это Chaohuperleidus, старейший известный член Perleidiformes, но вид самых разнообразных Saurichthys и несколько неописанных рыб также неизвестны.[38][39] Потенциальная жертва беспозвоночных Cartorhynchus включают мелкие аммониты и двустворчатые моллюски, а тилакоцефалан членистоногие Анкитоказокарис.[3][31]
Смотрите также
Рекомендации
- ^ а б c d е ж грамм час я j k л м Motani, R .; Jiang, D.-Y .; Chen, G.-B .; Тинтори, А .; Rieppel, O .; Ji, C .; Хуанг, Ж.-Д. (2014). «Базальная ихтиозаврическая форма с короткой мордой из нижнего триаса Китая». Природа. 517 (7535): 485–488. Bibcode:2015Натура.517..485M. Дои:10.1038 / природа13866. PMID 25383536. S2CID 4392798.
- ^ а б Перкинс, Сид (2014). «Как у ихтиозавра появились плавники». Наука. Получено 8 мая, 2020.
- ^ а б c d е ж грамм Huang, J.-D .; Motani, R .; Jiang, D.-Y .; Ren, X.-X .; Тинтори, А .; Rieppel, O .; Чжоу, М .; Hu, Y.-C .; Чжан, Р. (2020). «Повторное развитие дурофагии во время облучения ихтиозавра после массового вымирания, на которое указывает скрытый прикус». Научные отчеты. 10 (1): 7798. Bibcode:2020НатСР..10.7798Н. Дои:10.1038 / s41598-020-64854-z. ЧВК 7210957. PMID 32385319.
- ^ Чжоу, М .; Jiang, D.-Y .; Motani, R .; Тинтори, А .; Ji, C .; Вс, З.-Я .; Ni, P.-G .; Лу, Х. (2017). "Черепная остеология, обнаруженная с помощью трехмерно сохранившихся черепов ихтиозавриформ раннего триаса. Chaohusaurus chaoxianensis (Reptilia: Ichthyosauromorpha) из Аньхоя, Китай ". Журнал палеонтологии позвоночных. 37 (4): e1343831. Дои:10.1080/02724634.2017.1343831. S2CID 90332288.
- ^ а б c d е ж грамм час я j Jiang, D.-Y .; Motani, R .; Huang, J.-D .; Тинтори, А .; Hu, Y.-C .; Rieppel, O .; Fraser, N.C .; Ji, C .; Kelley, N.P .; Fu, W.-L .; Чжан, Р. (2016). «Большой аберрантный ихтиозавровидный стебель, указывающий на раннее возникновение и гибель ихтиозавроморфов после вымирания в конце пермского периода». Научные отчеты. 6: 26232. Bibcode:2016НатСР ... 626232J. Дои:10.1038 / srep26232. ЧВК 4876504. PMID 27211319.
- ^ Jiang, D .; Motani, R .; Hao, W .; Schmitz, L .; Rieppel, O .; Sun, Y .; Солнце, З. (2008). «Новый примитивный ихтиозавр (Reptilia, Diapsida) из среднего триаса Панксиана, Гуйчжоу, юго-запад Китая и его положение в восстановлении биотики триаса». Прогресс естествознания. 18 (10): 1315–1319. Дои:10.1016 / j.pnsc.2008.01.039.
- ^ а б Motani, R .; Chen, X.H .; Jiang, D.-Y .; Cheng, L .; Тинтори, А .; Риппель, О. (2015). «Выпад кормления ранних морских рептилий и быстрая эволюция гильдий морских четвероногих». Научные отчеты. 5: 8900. Bibcode:2015НатСР ... 5Э8900М. Дои:10.1038 / srep08900. ЧВК 4354009. PMID 25754468.
- ^ Хусай, А. (2009). ""Пахиостоз «у водных амниот: обзор». Интегративная зоология. 4 (4): 325–340. Дои:10.1111 / j.1749-4877.2009.00146.x. PMID 21392306.
- ^ Талеви, М .; Фернандес, М. (2012). «Неожиданная гистология скелета ихтиозавра из средней юры Патагонии: последствия для эволюции микроструктуры костей среди вторичных водных четвероногих». Naturwissenschaften. 99 (3): 241–244. Bibcode:2012NW ..... 99..241T. Дои:10.1007 / s00114-012-0886-4. PMID 22290413. S2CID 2223683.
- ^ Chen, X.-H .; Motani, R .; Cheng, L .; Jiang, D.-Y .; Риппель, О. (2014). "Загадочная морская рептилия Наньчангозавр из нижнего триаса провинции Хубэй, Китай и филогенетическое родство Хупехсучии ». PLOS ONE. 9 (7): e102361. Bibcode:2014PLoSO ... 9j2361C. Дои:10.1371 / journal.pone.0102361. ЧВК 4094528. PMID 25014493.
- ^ Motani, R .; Jiang, D.-Y .; Тинтори, А .; Rieppel, O .; Chen, G.-B .; Вы, Х. (2015). «Первое свидетельство централии в ихтиоптеригии, повторяющих предвзятость педоморфных признаков при филогенетической реконструкции морских рептилий». Журнал палеонтологии позвоночных. 35 (4): e948547. Дои:10.1080/02724634.2014.948547. S2CID 83576278.
- ^ Chen, Z.Q .; Бентон, М.Дж. (2012). «Время и образец восстановления биоты после массового вымирания в конце пермского периода». Природа Геонауки. 5 (6): 375–383. Bibcode:2012Натуральная ... 5..375C. Дои:10.1038 / ngeo1475.
- ^ Liu, J .; Hu, S.X .; Rieppel, O .; Jiang, D.-Y .; Benton, M.J .; Kelley, N.P .; Aitchison, J.C .; Чжоу, C .; Wen, W .; Huang, J .; Xie, T .; Львов, Т. (2014). «Гигантский нотозавр (Reptilia: Sauropterygia) из среднего триаса на юго-западе Китая и его значение для восстановления биотики триаса». Научные отчеты. 4: 7142. Bibcode:2014НатСР ... 4Э7142Л. Дои:10.1038 / srep07142. ЧВК 4245812. PMID 25429609.
- ^ Benton, M.J .; Zhang, Q .; Hu, S .; Chen, Z.-Q .; Wen, W .; Liu, J .; Huang, J .; Чжоу, C .; Xie, T .; Tong, J .; Чу, Б. (2013). «Исключительные биоты позвоночных из триаса Китая и расширение морских экосистем после массового вымирания в пермо-триасовом периоде». Обзоры наук о Земле. 125: 199–243. Bibcode:2013ESRv..125..199B. Дои:10.1016 / j.earscirev.2013.05.014.
- ^ а б Stubbs, T.L .; Бентон, М.Дж. (2016). «Экоморфологическая диверсификация мезозойских морских рептилий: роль экологических возможностей и исчезновения». Палеобиология. 42 (4): 547–573. Дои:10.1017 / pab.2016.15. S2CID 3722056.
- ^ Kelley, N.P .; Мотани, Р. (2015). «Трофическая конвергенция приводит к морфологической конвергенции у морских четвероногих животных». Письма о биологии. 11 (1): 20140709. Дои:10.1098 / rsbl.2014.0709. ЧВК 4321144. PMID 25631228.
- ^ Fan, J.-X .; Shen, S.-Z .; Erwin, D.H .; Sadler, P.M .; MacLeod, N .; Cheng, Q.-M .; Hou, X.-D .; Yang, J .; Ван, X.-D .; Wang, Y .; Zhang, H .; Чен, X .; Li, G.-X .; Zhang, Y.-C .; Shi, Y.-K .; Юань, Д.-Х .; Chen, Q .; Zhang, L.-N .; Li, C .; Чжао, Ю.-Й. (2020). «Краткое изложение биоразнообразия морских беспозвоночных от кембрия до раннего триаса в высоком разрешении». Наука. 367 (6475): 272–277. Bibcode:2020Sci ... 367..272F. Дои:10.1126 / science.aax4953. PMID 31949075. S2CID 210698603.
- ^ Эмбри, А.Ф. (1997). «Границы глобальных последовательностей триаса и их идентификация в осадочном бассейне Западной Канады». Бюллетень канадской нефтяной геологии. 45 (4): 415–433.
- ^ Paton, M.T .; Иванов, А.В .; Fiorentini, M.L .; McNaughton, N.J .; Мудровская, И .; Резницкий, Л.З .; Демонтерова, Э. (2010). «Позднепермские и раннетриасовые магматические импульсы в Ангаро-Тасеевской синклинали, Южно-Сибирские ловушки и их возможное влияние на окружающую среду». Российская геология и геофизика. 51 (9): 1012–1020. Bibcode:2010RuGG ... 51.1012P. Дои:10.1016 / j.rgg.2010.08.009.
- ^ Takahashi, S .; Yamasaki, S.I .; Ogawa, K .; Kaiho, K .; Цучия, Н. (2015). «Редокс-условия в конце раннего триаса Панталасса». Палеогеография, палеоклиматология, палеоэкология. 432: 15–28. Bibcode:2015ГПП ... 432 ... 15Т. Дои:10.1016 / j.palaeo.2015.04.018.
- ^ Dick, D.G .; Максвелл, E.E. (2015). «Эволюция и исчезновение ихтиозавров с точки зрения количественного экокосмического моделирования». Письма о биологии. 11 (7): 20150339. Дои:10.1098 / rsbl.2015.0339. ЧВК 4528449. PMID 26156130.
- ^ Сандер, П.М.; Чен, X .; Cheng, L .; Ван, X. (2011). «Беззубый ихтиозавр с коротким носом из Китая предлагает позднетриасовую диверсификацию ихтиозавров, питающихся всасыванием». PLOS ONE. 6 (5): e19480. Bibcode:2011PLoSO ... 619480S. Дои:10.1371 / journal.pone.0019480. ЧВК 3100301. PMID 21625429.
- ^ Motani, R .; Ji, C .; Томита, Т .; Kelley, N .; Максвелл, Е .; Jiang, D.-Y .; Сандер, П. (2013). «Отсутствие ихтиозавров с присасывающим питанием и его значение для триасовой мезопелагической палеоэкологии». PLOS ONE. 8 (12): e66075. Bibcode:2013PLoSO ... 866075M. Дои:10.1371 / journal.pone.0066075. ЧВК 3859474. PMID 24348983.
- ^ Vandewalle, P .; Saintin, P .; Шардон, М. (1995). "Структура и движения щечной и глоточной челюстей в связи с кормлением Diplodus sargus". Журнал биологии рыб. 46 (4): 623–656. Дои:10.1111 / j.1095-8649.1995.tb01101.x.
- ^ Crofts, S.B .; Саммерс, А.П. (2014). «Как лучше всего разбить улитку: влияние формы зуба на раздавливающую нагрузку». Журнал интерфейса Королевского общества. 11 (92): 20131053. Дои:10.1098 / rsif.2013.1053. ЧВК 3899874. PMID 24430124.
- ^ Мазухова, Н .; Umbanhowar, P.B .; Гольдман, Д. (2013). «Управляемое ластами наземное движение робота, вдохновленного морской черепахой». Биоинспирация и биомиметика. 8 (2): 026007. Bibcode:2013БиБи .... 8b6007M. Дои:10.1088/1748-3182/8/2/026007. PMID 23612858.
- ^ Рыба, F.E. (2000). «Биомеханика и энергетика водных и полуводных млекопитающих: от утконоса до кита». Физиологическая и биохимическая зоология. 73 (6): 683–698. Дои:10.1086/318108. PMID 11121343.
- ^ Tytell, E.D .; Borazjani, I .; Sotiropoulos, F .; Baker, T.V .; Андерсон, E.J .; Лаудер, Г. (2010). «Разделение функциональных ролей морфологии и движения в плавании рыб». Интегративная и сравнительная биология. 50 (6): 1140–1154. Дои:10.1093 / icb / icq057. ЧВК 2981590. PMID 21082068.
- ^ Borazjani, I .; Сотиропулос, Ф. (2010). «О роли формы и кинематики в гидродинамике плавания самоходного корпуса и хвостового плавника». Журнал экспериментальной биологии. 213 (1): 89–107. Дои:10.1242 / jeb.030932. PMID 20008366. S2CID 27365252.
- ^ Gutarra, S .; Moon, B.C .; Rahman, I.A .; Palmer, C .; Lautenschlager, S .; Brimacombe, A.J .; Бентон, М.Дж. (2019). «Влияние эволюции строения тела на гидродинамическое сопротивление и энергетические потребности плавания у ихтиозавров». Труды Королевского общества B. 286 (1898): 20182786. Дои:10.1098 / rspb.2018.2786. ЧВК 6458325. PMID 30836867.
- ^ а б Ji, C .; Тинтори, А .; Jiang, D .; Мотани, Р. (2017). «Новые виды Thylacocephala (Arthropoda) из спата (нижний триас) Чаоху, провинция Аньхой, Китай». PalZ. 91 (2): 171–184. Дои:10.1007 / s12542-017-0347-7. S2CID 132045789.
- ^ а б Motani, R .; Jiang, D.-Y .; Тинтори, А .; Ji, C .; Хуанг, Ж.-Д. (2017). «Дивергенция мезозойских морских рептилий до и после массового вымирания, продиктованная зависимостью темпов эволюции от времени». Труды Королевского общества B. 284 (1854): 20170241. Дои:10.1098 / rspb.2017.0241. ЧВК 5443947. PMID 28515201.
- ^ Fu, W .; Jiang, D .; Montañez, I .; Meylers, S .; Motani, R .; Тинтори, А. (2016). «Эксцентриситет и наклонность определяли круговорот углерода в раннем триасе и их последствия для восстановления экосистемы после исчезновения». Научные отчеты. 6: 27793. Bibcode:2016НатСР ... 627793F. Дои:10.1038 / srep27793. ЧВК 4904238. PMID 27292969.
- ^ Li, M .; Ogg, J .; Zhang, Y .; Хуанг, С .; Хиннов, Л .; Chen, Z.Q .; Цзоу, З. (2016). «Астрономическая настройка конца пермского вымирания и раннетриасовой эпохи Южного Китая и Германии». Письма по науке о Земле и планетах. 441: 10–25. Bibcode:2016E и PSL.441 ... 10 л. Дои:10.1016 / j.epsl.2016.02.017.
- ^ Ji, C .; Zhang, C .; Jiang, D.Y .; Bucher, H .; Motani, R .; Тинтори, А. (2015). «Контроль возраста аммоноидов морских рептилий раннего триаса из Чаоху (Южный Китай)». Палеомир. 24 (3): 277–282. Дои:10.1016 / j.palwor.2014.11.009.
- ^ Li, S.Y .; Тонг, J.N .; Liu, K.Y .; Wang, F.J .; Хо, Ю.Ю. (2007). «Циклические отложения нижнего триаса в Чаоху, провинция Аньхой, Китай». Палеогеография, палеоклиматология, палеоэкология. 252 (1–2): 188–199. Bibcode:2007ППП ... 252..188С. Дои:10.1016 / j.palaeo.2006.11.043.
- ^ Jiang, D.-Y .; Motani, R .; Тинтори, А .; Rieppel, O .; Chen, G.-B .; Huang, J.-D .; Zhang, R .; Вс, З.-Я .; Цзи, К. (2014). "Раннетриасовый эозауроптеригий Majiashanosaurus discocoracoidis, ген. et sp. ноя (Reptilia, Sauropterygia) из Чаоху, провинция Аньхой, Китайская Народная Республика ". Журнал палеонтологии позвоночных. 34 (5): 1044–1052. Дои:10.1080/02724634.2014.846264. S2CID 129828376.
- ^ Солнце, З .; Тинтори, А .; Jiang, D .; Мотани, Р. (2013). «Новый перлейдид из спата (оленек, ранний триас) Чаоху, провинция Аньхой, Китай». Итальянская Ривиста ди Палеонтология и Стратиграфия. 119 (3): 275–285. Дои:10.13130/2039-4942/6040.
- ^ Тинтори, А .; Huang, J.-D .; Jiang, D.-Y .; Вс, З.-Я .; Motani, R .; Чен, Г. (2014). "Новый Saurichthys (Actinopterygii) из спата (ранний триас) Чаоху (провинция Аньхой, Китай) ». Итальянская Ривиста ди Палеонтология и Стратиграфия. 120 (2): 157–164. Дои:10.13130/2039-4942/6057.
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||




